mitochondria. W cytoplazmie komórek zwierzęcych i roślinnych znajdują się tak zwane organelle energetyczne - mitochondria (z greckiego „mitos” - nić, „chondrion” - ziarno). Kształt mitochondriów jest różny; mogą być owalne, w kształcie pręta, nitkowate o średniej średnicy 1 µm i długości 7 µm. Liczba mitochondriów zależy od czynności funkcjonalnej komórki i może sięgać dziesiątek tysięcy w latających mięśniach owadów.
Struktura wewnętrzna mitochondria (ryc. 11, 12; 15) badano za pomocą mikroskopu elektronowego. Mikrografie elektronowe pokazują, że mitochondria są zewnętrznie ograniczone przez zewnętrzną błonę, która zasadniczo ma taką samą strukturę jak błona plazmatyczna. Pod błoną zewnętrzną znajduje się błona wewnętrzna, która tworzy liczne fałdy - cristae. Wewnątrz mitochondriów znajdują się RNA, DNA i rybosomy, które różnią się od tych cytoplazmatycznych. W jego błony wbudowane są określone enzymy, za pomocą których energia jest przekształcana w mitochondria. składniki odżywcze w energię ATP, niezbędną do życia komórki i organizmu jako całości.
Ryż. 15. Schemat budowy mitochondriów
Plastydy. Są to organelle, które są unikalne dla komórek roślinnych. Istnieją trzy rodzaje plastydów: zielone chloroplasty, kolorowe (ale nie zielone) chromoplasty i bezbarwne leukoplasty.
Chloroplast(ryc. 16) kształtem przypomina dysk lub kulkę o średnicy 4-6 mikronów z podwójną membraną - zewnętrzną i wewnętrzną. Wewnątrz chloroplastu znajduje się DNA, rybosomy i specjalne struktury membranowe - ziarna połączone ze sobą oraz z wewnętrznym memorano i chloroplastem, każdy chloroplast ma około 50 ziaren, ułożonych w szachownicę dla lepszego wychwytywania światła. Membrany Gran zawierają zielony barwnik chlorofil. Dzięki chlorofilowi w chloroplastach energia jest przekształcana światło słoneczne w energię chemiczną ATP. Energia ATP jest wykorzystywana w chloroplastach do syntezy związki organiczne przede wszystkim węglowodany.
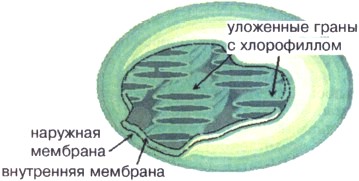
Ryż. 16. Schemat budowy chloroplastu
chromoplasty. Pigmenty czerwieni i żółty kolor, znajdujące się w chromoplastach, nadają różnym częściom roślin czerwony i żółty kolor. Korzeń marchwi, owoce pomidora są zabarwione dzięki pigmentom zawartym w chromoplastach. Połączenie chromoplastów zawierających różne pigmenty tworzy szeroką gamę kolorów kwiatów i owoców roślin.
Leukoplasty są miejscem gromadzenia rezerwowego składnika odżywczego - skrobi. Szczególnie dużo leukoplasty występuje w komórkach bulw ziemniaka. W świetle leukoplasty mogą przekształcić się w chloroplasty (w wyniku czego bulwy ziemniaka stają się zielone). Jesienią chloroplasty zamieniają się w chromoplasty, a zielone liście i owoce żółkną i czerwienieją.
Stały ruch komórki. Organelle ruchu. Wiele komórek jednokomórkowych i Organizmy wielokomórkowe mieć zdolność poruszania się. Odnosi się to zarówno do ruchu komórki w przestrzeni, jak i wewnątrzkomórkowego ruchu jej organelli. W środowisku płynnym ruch komórek odbywa się poprzez ruch wici i rzęsek (ryc. 10, 2, 8, 11). W ten sposób porusza się wiele organizmów jednokomórkowych, na przykład zielona euglena, wiciowiec, infusoria itp. Niektóre rodzaje bakterii poruszają się również za pomocą wici, długich i elastycznych, które szybko się obracają, zapewniając komórkom awans. Ameba i niektóre inne pierwotniaki, a także wyspecjalizowane komórki organizmów wielokomórkowych (na przykład limfocyty) poruszają się za pomocą wyrostków utworzonych na powierzchni komórek.
Komórka jest w środku w ciągłym ruchu. Podczas fagocytozy i pinocytozy błona plazmatyczna wystaje do komórki, lizosomy, pęcherzyki kompleksu Golgiego, poruszają się mitochondria, a na końcu porusza się sama cytoplazma.
Ruch komórkowy zapewnia cytoszkielet składający się z mikrotubul, mikrofilamentów i ośrodek komórkowy. Mikrotubule to długie wydrążone cylindry o średnicy 25 nm, których ściany zbudowane są z białek. Wici i rzęski komórek zwierzęcych i roślinnych składają się z równoległych mikrotubul. Wici różnią się od rzęsek tylko długością. Tak więc plemniki ssaków mają jedną wici o długości do 100 mikronów. Rzęski są krótsze od wici ponad 10 razy, w jednej komórce jest kilka tysięcy rzęsek. Mikrofilamenty to bardzo cienkie struktury złożone z tysięcy połączonych ze sobą cząsteczek białka aktyny. W komórkach mięśniowych wchodzą wraz z innymi filamentami białkowymi - miozyną, w kompleksy, które zapewniają funkcję skurczową tych komórek.
W cytoplazmie komórek wszystkich organizmów, w pobliżu jądra, znajduje się ośrodek komórkowy biorący udział w podziale komórkowym. Skład centrum komórkowego komórek zwierzęcych i niektórych roślin obejmuje centriolę (ryc. 11, 3). Centriole - tworzenie par. Zawiera dwa cylindry wykonane z mikrotubul i ułożone prostopadle do siebie.
Inkluzje komórkowe. Na koniec należy powiedzieć o licznych inkluzjach w cytoplazmie. Inkluzje nazywane są nietrwałymi strukturami cytoplazmy, które w przeciwieństwie do organelli pojawiają się lub znikają podczas życia komórki. Gęste inkluzje w postaci granulek zawierają rezerwowe składniki odżywcze (skrobię, białka, cukry, tłuszcze) lub produkty przemiany materii, których z różnych powodów nie można natychmiast usunąć. Wszystkie plastydy komórek roślinnych mają zdolność syntezy i gromadzenia zapasowych składników odżywczych.
W komórki roślinne gromadzenie rezerwowych składników odżywczych występuje również w wakuolach - workach błonowych z roztwór wodny sole i związki organiczne, które często zajmują prawie całą objętość komórki, wypychając jądro i cytoplazmę do błony plazmatycznej.
- Dlaczego mitochondria nazywane są „elektrowniami” komórki?
- Jakie struktury komórki biorą udział w jej ruchu?
- Co to są inkluzje komórkowe? Jaka jest ich rola w komórce?
- Opisz cechy strukturalne mitochondriów i chloroplastów w powiązaniu z ich funkcjami w komórce.
Centrum komórkowe- organelle widoczne pod mikroskopem optycznym w komórkach zwierzęcych i niższe rośliny. Zwykle znajduje się w pobliżu jądra lub w geometrycznym środku komórki i składa się z dwóch korpusów centrioli w kształcie pręcików, o wielkości około 0,3-1 mikrona. Pod mikroskopem elektronowym stwierdzono, że centriola jest cylindrem, którego ściany zbudowane są z dziewięciu par bardzo cienkich rurek. W środku cylindra znajduje się wnęka wypełniona jednorodną masą. Para centrioli jest otoczona jaśniejszą strefą - centrosferą.
Centrum komórkowe gra ważna rola podczas ruchu chromosomów podczas mitozy. Wiąże się z tym zdolność niektórych komórek do aktywnego ruchu. Świadczy o tym fakt, że u podstawy wici lub rzęsek ruchliwych komórek (pierwotniaków, plemników) znajdują się formacje o tej samej budowie co centrum komórki.
Organelle ruchu. Komórki mogą poruszać się za pomocą wyspecjalizowanych organelli, do których należą rzęski i wici. Rzęski komórkowe są zawsze liczne (u pierwotniaków ich liczba sięga setek i tysięcy), a długość wynosi 10-15 mikronów. Wici są najczęściej 1-8, ich długość wynosi 20-50 mikronów. Struktura rzęsek i wici zarówno w komórkach roślinnych, jak i zwierzęcych jest podobna. Pod mikroskopem elektronowym stwierdzono, że mikrotubule biegną na całej swojej długości. Dwie z nich znajdują się w centrum, a wokół nich na obwodzie leży kolejnych 9 par mikrotubul. Cała ta struktura jest pokryta błona cytoplazmatyczna który jest przedłużeniem błony komórkowej. Ruch wici i rzęsek zapewnia nie tylko ruch komórek w przestrzeni, ale także ruch różne substancje na powierzchni komórek, jak również przedostawanie się cząstek pokarmu do wnętrza komórki. U podstawy rzęsek i wici znajdują się ciała podstawowe, które również składają się z mikrotubul. Uważa się, że ciała podstawne są ośrodkiem powstawania mikrotubul wici i rzęsek. Z kolei ciała podstawowe często wywodzą się z centrum komórki.
Duża liczba organizmów jednokomórkowych i niektóre komórki wielokomórkowe nie mają specjalnych organelli ruchu i poruszają się za pomocą pseudopodiów (pseudopodiów). Ruch za pomocą pseudopodiów nazywa się ruchem ameboidalnym. Opiera się na ruchu cząsteczek specjalnych białek, zwanych białkami kurczliwymi.
Inkluzje najczęściej spotykane w komórkach roślinnych i różnią się od organelli tym, że są formacjami tymczasowymi, nietrwałymi. Ich liczba zależy od intensywności metabolizmu i stanu organizmu. Zazwyczaj są one widoczne w mikroskopie optycznym w postaci ziaren lub kropli o różnych rozmiarach i kształtach. Przez skład chemiczny rozróżnić: wtrącenia węglowodanowe, tłuszczowe i białkowe. Wtrącenia węglowodanowe i białkowe wyglądają jak ziarna, a wtrącenia tłuszczowe wyglądają jak kropelki. W komórkach roślinnych węglowodany odkładane są najczęściej w postaci ziaren skrobi, a u zwierząt – glikogenu. Ziarna białka znajdują się w dużych ilościach w cytoplazmie jaj zwierzęcych w postaci żółtka. Wiele z nich znajduje się w nasionach roślin. Kropelki tłuszczu są obecne w komórkach tkanka łączna zwierzęta i nasiona roślin. Komórki roślinne zawierają inkluzje krystaliczne (sole kwasy organiczne). W pewnych warunkach wszystkie rodzaje inkluzji mogą być wykorzystywane przez komórkę w toku jej aktywności życiowej, a następnie ponownie się kumulować.
W cytoplazmie wielu komórek roślinnych występują specjalne formacje - wakuole. Są to zbiorniki wypełnione mieszaniną roztworów różnych chemikaliów. Szczególnie duże wakuole powstają w starych komórkach.
|
Kryteria Dla porownania |
Typy rybosomów |
|
|
typ prokariotyczny |
typ eukariotyczny |
|
|
Odkrycie w komórkach |
Rybosomy eubakterii i sinic plastydowe rybosomy mitochondrialne Rybosomy Rybosomy Archaebacteria (podobne do eukariotycznych) |
Cytoplazmatyczny rybosomy eukariotyczne: Zwierząt, I rośliny |
|
Stosunek RNA: białko | ||
|
Ilość rRNA podjednostka |
1 cząsteczka o długości 1500-1600 nukleotydów (minimum - 950 w rybosomach mitochondrialnych) |
1 cząsteczka o długości około 1800 nukleotydów |
|
Ilość rRNA w dużym podjednostka |
2 cząsteczki: 1 cząsteczka o długości około 3000 nukleotydów, 1 cząsteczka o długości około 120 nukleotydów |
3 cząsteczki: 1 cząsteczka o długości około 4-5 tysięcy nukleotydów, 1 cząsteczka o długości około 120 nukleotydów, 1 cząsteczka o długości około 160 nukleotydów (cząsteczka ta jest podobna do jednego z regionów prokariotycznego rRNA) |
|
białko (prawie wszystkie cząsteczki są unikalne) |
Około 20 cząsteczek w małej podjednostce i 30 cząsteczek w dużej |
Około 30 cząsteczek na mała podjednostka i 40 cząsteczek w dużej |
Centrum komórkowe
Centrum komórkowe to organelle, które kontrolują tworzenie i wzrost mikrotubul w cytoszkielecie komórki eukariotycznej.(Rys. 38)
Ryż. 38. Centrum komórkowe 1 - trojaczki mikrotubul; 2 - promieniowe szprychy; 3 - centralna konstrukcja „koła wozu”; 4 - satelita; 5 - lizosom; 6 - dictyosomy kompleksu Golgiego; 7 - pęcherzyk graniczący; 8 - cysterna ziarnistej retikulum endoplazmatycznego; 10 - mitochondrium; 11 - szczątkowe ciało; 12 - mikrotubule; 13 - caroteca (wg R. Krstica, ze zmianami).
Główne funkcje centrum komórkowego to tworzenie układów mikrotubul cytoszkieletu, organelli ruchu i wrzeciona podziałowego.
Podstawą centrum komórkowego jest para centrioli (czasami ich liczba sięga 4-6-8 w komórkach poliploidalnych), znajdujących się w centrum komórki.
Centriole zostały po raz pierwszy odkryte i opisane przez V. Flemminga (1875), ale szczegółowo zbadano je dopiero za pomocą mikroskopu elektronowego. Centriole są blisko spokrewnione otoczka jądrowa, aw wielu niższych eukariontach (pierwotniaki, glony, grzyby, mezokarionty) centriole są wbudowane w otoczkę jądra.
Centrum komórkowe zawsze znajduje się w komórkach zwierząt wielokomórkowych. Prokarionty nie mają centrum komórkowego i tubuliny. W komórkach grzybów, glonów i zwierząt jednokomórkowych centrum komórkowe nie zawsze znajduje się, ale w komórkach Wyższe rośliny nie znaleziono (z rzadkimi wyjątkami). W przypadku braku centrum komórkowego jego funkcje u eukariontów pełni centrum tworzenia mikrotubul - CMTC.
Centriola to wydrążony cylinder o średnicy około 0,15 mikrona i długości 0,3-0,5 mikrona (rzadziej - kilka mikronów). Ściany centrioli składają się z 9 trojaczki mikrotubule tubuliny połączone mostkami dyneinowymi. Zazwyczaj centriole są ułożone parami: jedna centriola to centriola macierzysta, a druga to centriola potomna; centriola potomna jest prostopadła do centrioli macierzystej. Ta para centrioli dyplomowany- ma kształt litery T lub L. Diplosom otoczony jest gęstą warstwą cytoplazmy censtosfera, Lub centrosom. Podczas podziału komórki gęste pasma mikrotubul opuszczają centroferę, tworząc się astrosfera.
Podwojenie centrioli następuje mniej więcej w połowie interfazy: centriole matki i córki rozchodzą się, a z każdej centrioli wyrasta nowa centriola. Na różne grupy organizmów, proces ten ma swoje własne cechy.
Oprócz organelli komórki zawierają inkluzje komórkowe. Mogą być zawarte nie tylko w cytoplazmie, ale także w niektórych organellach, takich jak mitochondria i plastydy.
Co to są inkluzje komórkowe?
Są to formacje, które nie są trwałe. W przeciwieństwie do organoidów nie są tak stabilne. Ponadto mają znacznie prostszą budowę i pełnią funkcje pasywne, takie jak np. kopia zapasowa.
Jak są zbudowane?
Większość z nich ma kształt kropli, ale niektóre mogą być inne, na przykład podobne do plamy. Jeśli chodzi o rozmiar, może się różnić. Inkluzje komórkowe mogą być albo mniejsze niż organelle, albo tej samej wielkości lub nawet większe.
Składają się głównie z jednej konkretnej substancji, w większości przypadków organicznej. Może to być tłuszcz, węglowodany lub białko. 
Klasyfikacja
W zależności od tego, skąd pochodzi substancja, z której się składają, wyróżnia się następujące rodzaje inkluzji komórkowych:
- egzogenny;
- endogenny;
- wirusowy.
Egzogenne inkluzje komórkowe są zbudowane z związki chemiczne który wszedł do celi z zewnątrz. Te, które powstają z substancji wytwarzanych przez samą komórkę, nazywane są endogennymi. Wtręty wirusowe, chociaż są syntetyzowane przez samą komórkę, to jednak dzieje się to w wyniku wniknięcia do niej DNA wirusa. Komórka po prostu bierze to za swoje DNA i syntetyzuje z niego białko wirusa.
W zależności od funkcji, jakie pełnią inkluzje komórkowe, dzieli się je na pigmentowe, wydzielnicze i troficzne.
Ponadto inkluzje dzielą się na typy w zależności od konkretnych związków chemicznych, z których się składają. 
Inkluzje komórkowe: funkcje
Mogą pełnić trzy funkcje. Rozważ je w tabeli
To wszystko są funkcje formacji nietrwałych w komórce.
Inkluzje komórek zwierzęcych
Cytoplazma zwierzęcia zawiera zarówno inkluzje troficzne, jak i pigmentowe. Niektóre komórki mają również komórki wydzielnicze.
Troficzne w komórkach zwierzęcych są włączenie glikogenu. Mają kształt granulki o wielkości około 70 nm. 
Glikogen jest główną substancją rezerwową zwierzęcia. Jak dana substancja organizm magazynuje glukozę. Istnieją dwa hormony regulujące metabolizm glukozy i glukogenu: insulina i glukagon. Oba są produkowane przez trzustkę. Insulina jest odpowiedzialna za tworzenie glikogenu z glukozy, a glukagon, przeciwnie, bierze udział w syntezie glukozy.
Większość inkluzji glikogenu znajduje się w komórkach wątroby. Występują również w dużych ilościach w składzie mięśni, w tym serca. Inkluzje glikogenowe komórek wątroby mają postać granulek o wielkości około 70 nm. Zbierają się w małe skupiska. Inkluzje glikogenowe miocytów ( Komórki mięśniowe) mają okrągły kształt. Są pojedyncze, nieco większe niż rybosomy.
Scharakteryzowano również komórki zwierzęce inkluzje lipidowe. Są to również inkluzje troficzne, dzięki którym organizm może czerpać energię nagły wypadek. Składają się z tłuszczów i mają kształt łzy. Zasadniczo takie inkluzje są zawarte w komórkach tkanki łącznej tłuszczowej - lipocytach. Istnieją dwa rodzaje tkanki tłuszczowej: biała i brunatna. Białe lipocyty zawierają jedną dużą kroplę tłuszczu, brązowe komórki zawierają wiele małych.
Jeśli chodzi o inkluzje pigmentowe, komórki zwierzęce charakteryzują się tymi, które składają się z melaniny. Dzięki tej substancji tęczówka oka, skóra i inne części ciała mają określony kolor. Im więcej inkluzji melaniny w komórkach, tym ciemniejszy jest skład tych komórek.
Innym pigmentem, który można znaleźć w komórkach zwierzęcych jest lipofuscyna. Ta substancja ma żółto-brązowy kolor. Gromadzi się w mięśniu sercowym i wątrobie w miarę starzenia się narządów.
Inkluzje komórek roślinnych
Zawarte są również inkluzje komórkowe, których strukturę i funkcje rozważamy
Główne inkluzje troficzne w tych organizmach to ziarna skrobi. W swojej formie rośliny magazynują glukozę. Zazwyczaj inkluzje skrobiowe mają kształt soczewkowaty, kulisty lub jajowaty. Ich wielkość może się różnić w zależności od rodzaju rośliny i narządu, w którego komórkach się znajdują. Może wynosić od 2 do 100 mikronów.
Inkluzje lipidowe charakterystyczne również dla komórek roślinnych. Są to drugie co do częstości występowania inkluzje troficzne. Mają kulisty kształt i cienką membranę. Czasami nazywane są sferosomami.
Inkluzje białkowe występują tylko w komórkach roślinnych, nie są typowe dla zwierząt. Zbudowane są z prostych białek zwanych białkami. Inkluzje białkowe są dwojakiego rodzaju: ziarna aleuronowe i ciałka białkowe. Ziarna aleuronowe mogą zawierać albo kryształy, albo po prostu amorficzne białko. Tak więc pierwsze nazywane są złożonymi, a drugie - prostymi. Rzadziej występują proste ziarna aleuronowe, które składają się z amorficznego białka.
Jeśli chodzi o inkluzje pigmentowe, rośliny charakteryzują się plastoglobule. Zawierają karotenoidy. Takie inkluzje są charakterystyczne dla plastydów.
Wtrącenia komórkowe, których strukturę i funkcje rozważamy, składają się w większości z organicznych związków chemicznych, jednak w komórkach roślinnych są też takie, które powstają z Nie materia organiczna. Ten kryształy szczawianu wapnia. 
Występują tylko w wakuolach komórki. Kryształy te mogą mieć najróżniejsze formy i często są dla nich indywidualne pewne rodzaje rośliny.
Żywe organizmy składające się z jednej komórki często mają zdolność aktywnego poruszania się. Mechanizmy ruchu, które powstały w procesie ewolucji, są bardzo różnorodne.
Organellami ruchu są pseudopodia (u sarkodów i niektórych wiciowców), wici (u wiciowców) lub rzęski (u orzęsków).
Ponadto ruch może być realizowany w wyniku skurczu specjalnych mikrowłókien - myonemów, znajdujących się pod osłonami w cytoplazmie (w apikompleksach)
Ruch ameby ma swoją nazwę od najprostszego organizmu - ameby. (patrz Załącznik nr 6). Narządami ruchu w amebie są fałszywe nogi - pseudopodia, które są wypukłościami cytoplazmy. Tworzą się w różne miejsca powierzchnia cytoplazmy. Mogą zniknąć i pojawić się ponownie w innym miejscu.
pseudopodia(od pseudo... i greckiej ropy, rodzaj p. podos - stopa) (pseudopody), przejściowe wyrostki cytoplazmatyczne w organizmach jednokomórkowych (kłącza, sporozoany itp.) oraz w niektórych komórkach (leukocyty, makrofagi itp.) organizmów wielokomórkowych . Służą do przenoszenia i wychwytywania żywności i innych cząstek.
Ruch za pomocą wici jest charakterystyczny dla wielu jednokomórkowych alg (na przykład chlamydomonas), pierwotniaków (na przykład zielonej eugleny) i bakterii. Narządami ruchu w tych organizmach są wici - cytoplazmatyczne wyrostki na powierzchni cytoplazmy.
wici (Patrz Załącznik nr 7). Rozmiary od 2-5 mikronów do 1 mm. Poruszają się za pomocą jednej lub wielu wici, których długość u jednego osobnika może być różna. Tylko nieliczne gatunki mogą tworzyć pseudopodia. Zwykle rozmnażają się bezpłciowo w wyniku podłużnego podziału komórek; niektórzy widzieli rozmnażanie płciowe. Zdecydowana większość wiciowców to gatunki wolno żyjące w wodach morskich i słodkich, a także w glebie. Formy planktonowe odgrywają ważną rolę w obiegu materii; jest wiele typów wskaźniki biologiczne zanieczyszczenie wody.
Struktura komórkowa.
Komórka, elementarny system życia, podstawa struktury i życia wszystkich zwierząt i roślin. Komórki istnieją jako niezależne organizmy (na przykład pierwotniaki, bakterie) oraz jako część organizmów wielokomórkowych, w których znajdują się komórki płciowe służące do rozmnażania oraz komórki ciała (somatyczne), różniące się budową i funkcjami (na przykład nerwy, kości mięśniowa, wydzielnicza).
Rozmiary komórek wahają się od 0,1-0,25 mikrona (niektóre bakterie) do 155 mm (jajo strusia w skorupce).
Komórka każdego organizmu jest kompletna żyjący system. Składa się z trzech nierozerwalnie połączonych części: błony, cytoplazmy i jądra. (Patrz Załącznik nr 9).
1. Błona komórkowa.
Błona komórkowa bezpośrednio oddziałuje ze środowiskiem zewnętrznym i oddziałuje z sąsiednimi komórkami (w organizmach wielokomórkowych).
Ściana komórkowa ma złożona struktura. Składa się z warstwy zewnętrznej i błony plazmatycznej znajdującej się pod spodem.
Komórki zwierzęce i roślinne różnią się budową warstwy zewnętrznej (patrz Załącznik nr 10). U roślin, podobnie jak u bakterii, niebiesko-zielonych alg i grzybów, na powierzchni komórek znajduje się gęsta skorupa lub ściana komórkowa. W większości roślin składa się z błonnika. Ściana komórkowa pełni niezwykle ważną rolę: jest zewnętrzną ramką, ochronną powłoką, zapewnia turgor komórek roślinnych: woda, sole, cząsteczki wielu substancji organicznych przechodzą przez ścianę komórkową.
Zewnętrzna warstwa powierzchni komórek zwierzęcych, w przeciwieństwie do ścian komórkowych roślin, jest bardzo cienka i elastyczna. Nie jest widoczny pod mikroskopem świetlnym i składa się z różnych polisacharydów i białek. Nazywa się warstwę powierzchniową komórek zwierzęcych glikokaliks.
Glycocalyx pełni przede wszystkim funkcję bezpośredniego łączenia komórek zwierzęcych ze środowiskiem zewnętrznym, ze wszystkimi otaczającymi je substancjami. Mając znikomą grubość (mniej niż 1 mikron) / zewnętrzna warstwa komórki zwierzęcej nie pełni roli podporowej, co jest charakterystyczne dla ścian komórkowych roślin. Tworzenie glikokaliksu, a także ścian komórkowych roślin, następuje z powodu żywotnej aktywności samych komórek.
2. Cytoplazma.
Cytoplazma (z cyto… i osocza), termin „cytoplazma” zaproponował E. Strasburger (1882).
Ograniczony od otoczenie zewnętrzne błona plazmatyczna, cytoplazma jest wewnętrznym półpłynnym środowiskiem komórek (patrz Załącznik nr 8). w cytoplazmie komórki eukariotyczne znajduje się jądro i różne organelle.
Jądro znajduje się w centralnej części cytoplazmy. Zawiera również różnorodne inkluzje - produkty aktywności komórkowej, wakuole, a także najmniejsze rurki i włókna, które tworzą szkielet komórki.
wakuole (Patrz Załącznik nr 8)(francuska wakuola, z łac. vacuus - pusta), ubytki w komórkach zwierzęcych i roślinnych lub Jednokomórkowe organizmy. Istnieją wakuole trawienne i kurczliwe (pulsujące), które regulują ciśnienie osmotyczne i służą do usuwania produktów rozpadu z organizmu. (patrz Załącznik nr 11).
Białka dominują w składzie głównej substancji cytoplazmy.
Główne procesy metaboliczne zachodzą w cytoplazmie, która łączy jądro i wszystkie organelle w jedną całość, zapewnia ich interakcję, aktywność komórki jako jednego integralnego żywego systemu.
Eksperymentalnie możliwe jest uzyskanie żywych niejądrowych komórek cytoplastów, które mogą syntetyzować białka, lipidy, ATP w ciągu 1-3 dni. Potem oczywiście umierają z powodu niemożności syntezy nowych RNA przy braku jądra.
Objętość cytoplazmy różne komórki zmienia się: w limfocytach jest w przybliżeniu równa objętości jądra, aw komórkach wątroby cytoplazma stanowi 94% całkowitej objętości komórek.
Formalnie w cytoplazmie wyróżnia się trzy części: organelle, inkluzje i hialoplazmę.
Organelle- składniki obowiązkowe dla każdej komórki, bez których komórka nie może utrzymać swojego istnienia.
Jednobłonowe organelle układu wakuolarnego obejmują retikulum endoplazmatyczne, lizosomy, aparat Golgiego, peroksysomy i błonę plazmatyczną.
Organelle dwubłonowe to mitochondria i plastydy. Ta grupa obejmuje również Jądro komórkowe.
wspólna własność organelli błonowych polega na tym, że są one zbudowane z błon lipoproteinowych, które zamykają się na sobie, tak że tworzą zamknięte wnęki i tym samym dzielą cytoplazmę na grupy funkcjonalnie różnych przedziałów.
Rybosomy, centrosomy komórek zwierzęcych należą do organelli niebłonowych.
Inkluzje_ _ nie zawsze występują i stanowią złogi substancji zapasowych (glikogen, żółtko) lub nagromadzenia produktów przemiany materii (pigmenty, kryształki soli w roślinach).
Hialoplazma(od „hualine” - przezroczysty) - to jest główna plazma, cytosol, prawda środowisko wewnętrzne komórki. Skład hialoplazmy jest bardzo złożony, a konsystencja zbliża się do żelu.
Żele- ustrukturyzowane układy koloidalne z ciekłym ośrodkiem dyspersyjnym, które pod wpływem czynników zewnętrznych lub czynniki wewnętrzne mogą zmienić swój stan skupienia i przejść do bardziej płynnej fazy - zolu. Podobne przejścia żel-zol mogą zachodzić w cytoplazmie pod wpływem białka aktyny, a jej stan zmienia się w różnych częściach komórki, co zapewnia ruch całej komórki. Gdy aktyna fibrylarna oddziałuje z białkami typu fibryny, żel stabilizuje się, a po związaniu z białkami, których aktywność zależy od stężenia jonów Ca (np. gelsolina), cały układ przechodzi w stan ciekły.
Nasilenie elementów szkieletu komórkowego, w tym jego składnika aktyny, może się znacznie zmieniać podczas cyklu komórkowego.
Funkcjonalne znaczenie cytoplazmy jest bardzo duże.
Hialoplazma, oprócz różnych jonów związków nieorganicznych, zawiera enzymy biorące udział w syntezie aminokwasów, nukleotydów, Kwasy tłuszczowe, cukry. Na rybosomach i polirybosomach osadzonych na błonach syntetyzowane są różne białka, które zapewniają metabolizm komórkowy.
3. Retikulum endoplazmatyczne.
Cała wewnętrzna strefa cytoplazmy jest wypełniona licznymi małymi kanałami i wnękami, których ściany są błonami o strukturze podobnej do błony plazmatycznej. Kanały te rozgałęziają się, łączą ze sobą i tworzą sieć zwaną retikulum endoplazmatycznym lub retikulum endoplazmatycznym.
Retikulum endoplazmatyczne (patrz Załącznik nr 12) niejednorodny w strukturze. Znane są dwa jego rodzaje - ziarnisty (szorstki) i gładki.
Na membranach kanałów i wnęk sieci ziarnistej znajduje się wiele małych okrągłych ciał - rybosomów, które nadają membranom szorstki wygląd.
Błony retikulum endoplazmatycznego gładkiego nie mają na swojej powierzchni rybosomów.
Retikulum endoplazmatyczne spełnia wiele różnych funkcji. Główną funkcją ziarnistej retikulum endoplazmatycznego jest udział w syntezie białek, która odbywa się w rybosomach.
Na błonach gładkiej retikulum endoplazmatycznego syntetyzowane są lipidy i węglowodany. Wszystkie te produkty syntezy gromadzą się w kanałach i jamach, a następnie są transportowane do różnych organelli komórkowych, gdzie są konsumowane lub gromadzone w cytoplazmie jako wtręty komórkowe.
Retikulum endoplazmatyczne łączy główne organelle komórki (patrz Załącznik nr 8).
4. Błony biologiczne.
Błony biologiczne (łac. „membrana” - skóra, błona), cienkie (o grubości nie większej niż 10 nm) błony lipoproteinowe, składające się z podwójnej warstwy cząsteczek lipidów, w skład której wchodzą cząsteczki różnych białek. Znajdują się one na powierzchni komórek (błona plazmatyczna) i cząstek wewnątrzkomórkowych (jądra, mitochondria itp.). Masowo, w zależności od rodzaju błony, lipidy stanowią 25-60%. i udział białek - 40-75%. Wiele błon zawiera węglowodany, których ilość może osiągnąć 2-10%.
Zdrowie komórki, czas jej życia w dużej mierze zależy od stanu błony.
a) Właściwości membran.
Całkowita przepuszczalność dla wody. Membrana zawsze przepuszcza wodę do komórki lub na zewnątrz, w zależności od tego, gdzie stężenie wody jest większe. Ten ruch substancji z obszaru o wysokim stężeniu do obszaru o niższym stężeniu nazywa się dyfuzją. Dyfuzja substancji nie wymaga energii.
Selektywne przewodnictwo substancji rozpuszczonych:
Ujemnie naładowane cząsteczki szybciej i łatwiej przenikają przez membranę. Substancje rozpuszczalne w tłuszczach łatwiej przenikają przez błonę niż substancje rozpuszczalne w wodzie. Małe cząsteczki łatwiej przenikają przez błonę niż duże.
b) Aktywny transport substancji.
Niektóre substancje są w stanie przeniknąć przez membranę w kierunku przeciwnym do ich dyfuzji, czyli z miejsca o niskim poziomie do miejsca o większym wysokie stężenie. Poprzez transport aktywny nadmiar jonów sodu, wodoru i chloru jest stale usuwany z komórki. Przeciwnie, fosforany, glukoza, aminokwasy aktywnie przenikają do cytoplazmy. Transport aktywny zawsze wiąże się z wydatkami energetycznymi.
Błona może zostać częściowo utracona podczas życia komórki, w wyniku tworzenia się wakuoli (pęcherzyków) trawiennych. Błona jest regularnie odnawiana w wyniku pracy specjalnych organelli. syntetyzowanie wakuoli błonowych. Te wakuole przyczepiają się w dowolne miejsce błony komórkowej, przywracając jej dawne wymiary i właściwości.
c) Lipidy błonowe.
Skład lipidów membrany biologiczne bardzo zróżnicowane. charakterystyczni przedstawiciele lipidami błony komórkowej są fosfolipidy, sfingomieliny i cholesterol (lipid steroidowy).
charakterystyczna cecha lipidów błonowych polega na podziale ich cząsteczek na dwie funkcjonalnie różne części: niepolarne, pozbawione ładunku ogony, składające się z kwasów tłuszczowych i naładowane głowy polarne.
Głowy polarne niosą ładunki ujemne lub może być neutralny. Obecność niepolarnych ogonów wyjaśnia dobrą rozpuszczalność lipidów w tłuszczach i rozpuszczalnikach organicznych. W eksperymencie mieszając lipidy wyizolowane z błon z wodą można otrzymać warstwy dwucząsteczkowe lub błony o grubości około 7,5 nm, gdzie strefy obwodowe warstwy to hydrofilowe głowy polarne, a strefa środkowa to nienaładowane ogony cząsteczek lipidów . Wszystkie naturalne błony komórkowe mają taką samą strukturę.
Błony komórkowe różnią się znacznie pod względem składu lipidów. Na przykład, membrany plazmatyczne komórki zwierzęce są bogate w cholesterol (do 30%) i mają mało lecytyny, podczas gdy błony mitochondrialne są bogate w fosfolipidy i ubogie w cholesterol.
Cząsteczki lipidów mogą poruszać się wzdłuż warstwy lipidowej, mogą obracać się wokół własnej osi, a także przemieszczać się z warstwy na warstwę. Białka pływające w „jeziorze lipidowym” mają również pewną ruchliwość boczną. Skład lipidów po obu stronach błony jest różny, co determinuje asymetrię w budowie warstwy bilipidowej.
d) Białka błonowe.
Cząsteczki białka, które są znacznie cięższe i większe niż cząsteczki lipidów, są niejako rozproszone w warstwie bilipidowej błony. Większość cząsteczek białka jest częściowo zanurzona w błonie i wiąże się z lipidami poprzez oddziaływania hydrofobowe. Są to tak zwane białka integralne.
Inne białka są związane tylko z polarnymi „końcami” cząsteczek lipidów i znajdują się na powierzchni warstwy bilipidowej, podczas gdy inne przenikają przez błonę na wylot. Asymetria w budowie błon najwyraźniej przejawia się w odniesieniu do białek integralnych. Są ściśle zorientowane: ich N-końce zaglądają do jamy wakuoli lub do środowiska pozakomórkowego.
Błony różnią się znacznie składem białek, które faktycznie determinują ich właściwości funkcjonalne. Wszystkie białka błonowe można podzielić na trzy grupy: enzymy, białka receptorowe i białka strukturalne.
Różne membrany mają charakterystyczny zestaw enzymów. Na przykład ATPaza zależna od K-Na, która bierze udział w transporcie jonów, jest zlokalizowana w błonie plazmatycznej. Białka receptorowe specyficznie wiążą się z pewnymi substancjami i niejako „rozpoznają” je: są to białka receptorowe dla hormonów, do rozpoznawania powierzchni sąsiednich komórek, wirusów itp.
Białka strukturalne zapewniają wytrzymałość błony i są związane z różnymi strukturami białkowymi w cytoplazmie. Na przykład w komórkach nabłonkowych specjalne białka błony plazmatycznej wiążą się z elementami cytoszkieletu i uczestniczą w tworzeniu szeregu połączeń międzykomórkowych, takich jak desmosomy, kontakty adhezyjne itp.
e) Węglowodany błonowe.
Są one związane głównie z membranami. glikoproteiny - cząsteczki białek połączone kowalencyjnie z łańcuchami węglowodanów. Z reguły cząsteczki węglowodanów znajdują się w zewnętrznych warstwach błon. Najczęściej są to krótkie łańcuchy liniowe lub rozgałęzione, do których należą galaktoza, manioza, fruktoza, sacharoza, N-acetyloglukozamina. arabinoza. ksyloza itp.
f) Synteza błon komórkowych.
Wszystkie błony komórkowe (z wyjątkiem błon mitochondrialnych i plastydowych) są zbudowane w ziarnistej retikulum endoplazmatycznym. Oddzielają się od niego małe błoniaste wakuole, które łączą się z błonami aparatu Golgiego. Pęcherzyki błonowe, wytwarzane z kolei przez aparat Golgiego, służą do budowy błony plazmatycznej, wakuoli wydzielniczych i wszystkich innych składników błony komórkowej.
5. Membrana plazmowa.
błona plazmatyczna (Patrz Załącznik nr 8)(plasmalemma, błona komórkowa), pod glikokaliksem i Ściana komórkowa znajduje się błona plazmatyczna, otaczająca protoplazmę komórek roślinnych i zwierzęcych. Grubość błony plazmatycznej wynosi około 10 nm, badanie jej struktury i funkcji jest możliwe tylko za pomocą mikroskopu elektronowego.
Pełni nie tylko rolę bariery mechanicznej, ale przede wszystkim ogranicza swobodny dwukierunkowy przepływ do i z komórki substancji nisko- i wysokocząsteczkowych. Co więcej, plazmalemia działa jak struktura, która „rozpoznaje” różne substancje chemiczne oraz regulowanie selektywnego transportu tych substancji do komórki. Podobnie jak inne błony komórkowe, powstaje i jest aktualizowana z powodu aktywność syntetyczna retikulum endoplazmatycznego i ma podobną do nich strukturę.
a) Barierowo-transportowa rola plazmalemmy.
Mechaniczna stabilność błony plazmatycznej zależy nie tylko od właściwości samej błony, ale także od właściwości przylegającego do niej glikokaliksu i warstwy korowej cytoplazmy.
Zewnętrzna powierzchnia błony komórkowej (patrz Załącznik nr 13). pokryty luźną włóknistą warstwą substancji o grubości 3-4 nm - glikokaliksu. Składa się z rozgałęzionych łańcuchów polisacharydowych integralnych białek błonowych, pomiędzy którymi mogą znajdować się wydzielane przez komórkę glikolipidy i proteoglikany. Niektóre komórkowe enzymy hydrolityczne zaangażowane w pozakomórkowy rozkład substancji (trawienie pozakomórkowe, na przykład w nabłonku jelitowym) są natychmiast wykrywane.
Warstwa korowa cytoplazmy o grubości 0,1-0,5 µm nie zawiera rybosomów i struktur błonowych, ale jest bogata w mikrofilamenty aktyny.
Błona plazmatyczna, podobnie jak inne błony komórkowe lipoprotein, jest półprzepuszczalna. Woda i rozpuszczone w niej gazy mają największą siłę penetracji. Transport jonów może odbywać się biernie, bez zużycia energii. W tym przypadku niektóre białka transportujące błonę tworzą kompleksy molekularne, kanały, przez które jony przechodzą przez błonę na drodze prostej dyfuzji.
W innych przypadkach specjalne białka nośnikowe błony selektywnie wiążą się z jednym lub drugim jonem i transportują go przez błonę. Ten rodzaj transferu nazywany jest transportem aktywnym i odbywa się za pomocą białkowych pomp jonowych. Na przykład wydając 1 cząsteczkę ATP, system pompy K-Na wypompowuje 3 jony Na z komórki w jednym cyklu i pompuje 2 jony K wbrew gradientowi stężeń.
W połączeniu z aktywnym transportem jonów różne cukry, nukleotydy i aminokwasy przenikają przez plazmalemmę. Makrocząsteczki, takie jak białka, nie przechodzą przez błonę. One, podobnie jak większe cząstki substancji, są transportowane do komórki endocytoza . Podczas endocytozy pewna część plazmalemmy wychwytuje, otacza materiał pozakomórkowy i zamyka go w wakuoli błonowej. Ta wakuola jest endosomem - łączy się w cytoplazmie z pierwotnym lizosomem i następuje trawienie wychwyconego materiału.
Endocytoza jest formalnie podzielona na fagocytozę (wchłanianie dużych cząstek przez komórkę) i pinocytozę (wchłanianie roztworów).
Błona plazmatyczna bierze również udział w usuwaniu substancji z komórki za pomocą egzocytoza - odwrotny proces endocytozy.
b) Receptorowa rola plazmalemmy.
Białka nośnikowe zewnętrznej błony komórkowej są również receptorami, które rozpoznają określone jony i wchodzą z nimi w interakcje. Białka błonowe lub elementy glikokaliksu mogą działać jako receptory na powierzchni komórki. Takie miejsca wrażliwe na poszczególne substancje są rozsiane po powierzchni komórki lub gromadzone w małych strefach.
Rolą wielu receptorów komórkowych jest nie tylko wiązanie określonych substancji, ale także przekazywanie sygnałów z powierzchni do wnętrza komórki. Na przykład, gdy hormon działa na komórkę, łańcuch zdarzeń rozwija się w następujący sposób: cząsteczka hormonu specyficznie oddziałuje z białkiem receptora błony komórkowej i bez wnikania do komórki aktywuje enzym syntetyzujący cykliczny AMP. Ten ostatni aktywuje lub hamuje wewnątrzkomórkowy enzym lub grupę enzymów.
Różnorodność i specyficzność zestawów receptorów na powierzchni komórki prowadzi do powstania bardzo skomplikowany system markery, które pozwalają komórkom odróżnić „swoich” (tego samego osobnika lub tego samego gatunku) od „obcych”.
c) Połączenia międzykomórkowe.
W organizmach wielokomórkowych złożone zespoły komórek powstają w wyniku interakcji międzykomórkowych. W bliskim sąsiedztwie komórek glikokaliks zapewnia adhezję komórek dzięki obecności w nim transbłonowych kadheryn glikoprotein. Jest to prosty kontakt międzykomórkowy, w którym przerwa między komórkami wynosi 10-20 nm.
W nabłonku często występuje gęste lub blokujące połączenie, w którym zewnętrzne warstwy dwóch błon plazmatycznych są jak najbliżej, a globulki integralnych białek błonowych leżą w punktach ich kontaktu. Taki kontakt jest nieprzepuszczalny dla molekuł i jonów, zamyka przestrzenie międzykomórkowe.
Połączenia kotwiące, czyli kontakty, nie tylko łączą błony plazmatyczne sąsiednich komórek, ale także wiążą się z elementami włóknistymi cytoszkieletu. Na przykład w przypadku desmosomów, które wyglądają jak blaszki lub guziki, przestrzeń międzykomórkowa charakteryzuje się obecnością gęstej warstwy glikoprotein desmogleiny.
Po stronie cytoplazmatycznej warstwa białka desmoplakiny, związana z włóknami pośrednimi cytoszkieletu, sąsiaduje z plazmalemmą.
Połączenia szczelinowe są uważane za połączenia komunikacyjne komórek. W strefie kontaktu szczelinowego może ich być od 20-30 do kilku tysięcy koneksony - cylindryczne struktury białkowe z kanał wewnętrzny 2 nm średnicy. Każdy konekson składa się z 6 podjednostek białka łączącego. Koneksony pełnią rolę bezpośrednich kanałów międzykomórkowych, przez które jony i substancje niskocząsteczkowe mogą dyfundować z komórki do komórki.
6. Lizosomy.
Lizosomy (z liz i greckiej somy - ciało). Są to małe okrągłe ciała. Każdy lizosom jest oddzielony od cytoplazmy błoną. Wewnątrz lizosomu znajdują się enzymy, które mogą rozkładać (tj. lizować - stąd nazwa) białka, polisacharydy, peptydy, kwasy nukleinowe (patrz Załącznik nr 8).
Jest to bardzo różnorodna klasa pęcherzyków o wielkości 0,1-0,4 µm, ograniczona pojedynczą błoną (o grubości około 7 nm), z niejednorodną zawartością wewnątrz. Powstają w wyniku działania retikulum endoplazmatycznego i aparatu Golgiego i pod tym względem przypominają wakuole wydzielnicze.
Ich główną rolą jest udział w procesach wewnątrzkomórkowego rozszczepienia zarówno egzogennych, jak i endogennych makrocząsteczek biologicznych. charakterystyczna cecha lizosomów polega na tym, że zawierają one około 40 enzymów hydrolitycznych: proteinazy, nukleazy, fosfatazy, glikozydazy itp., z których optimum przeprowadza się przy pH 5.
W lizosomach kwaśna wartość środowiska powstaje dzięki obecności w ich błonach „pompy” protonowej, która zużywa energię ATP.
Ponadto białka nośnikowe są wbudowane w błony lizosomów w celu transportu z lizosomu do cytoplazmy produktów hydrolizy: monomerów rozszczepionych cząsteczek - aminokwasów, cukrów, nukleotydów, lipidów. Aby nie trawić samych siebie, elementy błon lizosomów są chronione przez oligosacharydy, które zapobiegają interakcji hydrolaz z nimi.
Wśród cząstek lizosomalnych o różnej morfologii wyróżnia się cztery typy: lizosomy pierwotne i wtórne, autofagosomy i ciała szczątkowe.
lizosomy pierwotne.
Zwykle są to małe błoniaste pęcherzyki o średnicy około 100 nm z bezstrukturalną zawartością zawierającą aktywną kwaśną fosfatazę, enzym markerowy dla lizosomów. Wykazano, że enzymy lizosomalne są syntetyzowane, jak zwykle, w siateczce ziarnistej i są upakowane w pęcherzykach błonowych w aparacie Golgiego. Cały sposób powstawania pierwotnych lizosomów jest bardzo podobny do powstawania ziarnistości wydzielniczych np. w komórkach trzustki.
lizosomy wtórne.
Fagocytarne lub pinocytowe wakuole łączą się z pierwotnymi lizosomami, tworząc wtórne lizosomy. Rozpoczyna się proces rozszczepiania substratów wchłoniętych przez komórkę pod działaniem hydrolaz zawartych w pierwotnym lizosomie. Substancje biogenne rozkładane są do monomerów, które są transportowane przez błonę lizosomu do cytoplazmy, gdzie są ponownie wykorzystywane, wchodzą w skład różnych syntetycznych i procesy metaboliczne. Zróżnicowanie wielkości i struktury lizosomów komórkowych związane jest przede wszystkim z różnorodnością lizosomów wtórnych – produktów fuzji wakuoli endocytarnych z lizosomami pierwotnymi.
Pozostałości ciał.
Rozszczepianie makrocząsteczek biogennych wewnątrz lizosomów może nie dobiec końca. W tym przypadku niestrawione produkty gromadzą się w jamie lizosomu, a lizosom wtórny staje się ciałem resztkowym (telolizosomem). Ich zawartość jest zagęszczana i przestawiana. Często w ciałach resztkowych dochodzi do wtórnej strukturyzacji niestrawionych lipidów, które tworzą złożone struktury warstwowe. Osadzają się tam również pigmenty.
Autolizosomy.
Zgodnie z morfologią autolizosomy (autofagosomy) zaliczane są do lizosomów wtórnych, ale wewnątrz tych cząstek znajdują się fragmenty lub nawet całe struktury cytoplazmatyczne, takie jak mitochondria, plastydy, rybosomy, elementy retikulum itp. Przyjmuje się, że proces autofagocytozy jest związane z selekcją i niszczeniem zmienionych, „zepsutych” składników komórkowych.
7. Aparat Golgiego.
Aparat Golgiego (kompleks Golgiego) nazwany na cześć K. Golgiego, organoid komórkowy.
W wielu komórkach zwierzęcych, takich jak komórki nerwowe, przybiera postać złożonej sieci zlokalizowanej wokół jądra. W komórkach roślin i pierwotniaków aparat Golgiego jest reprezentowany przez pojedyncze ciała w kształcie sierpa lub pręta. Struktura tego organoidu jest podobna w komórkach organizmów roślinnych i zwierzęcych, pomimo różnorodności jego kształtu. (patrz Załącznik nr 8).
Skład aparatu Golgiego obejmuje: wnęki ograniczone membranami i rozmieszczone w grupach (po 5-10); duże i małe pęcherzyki znajdujące się na końcach wnęk. Wszystkie te elementy tworzą jeden kompleks.
Aparat Golgiego pełni wiele ważnych funkcji.
Doprowadza białka do ostatecznej formy roboczej, na przykład „zszywa” niektóre białka w duże kompleksy białkowe, przyłącza niezbędne jony metali do niektórych białek.
Tworzy pęcherzyki błonowe, które opuszczając kompleks Golgiego albo przywracają błonę komórkową, albo zamieniają się w lizosomy. Lizosomy to organelle błonowe komórki, reprezentujące mikroskopijne pęcherzyki wypełnione enzymami trawiennymi.
Pełnią funkcje trawienne i ochronne. Na przykład w komórkach trzustkowych ssaków, enzymy trawienne które gromadzą się we wnękach organoidu.
Mogą sklejać się z wakuolą trawienną, wlewając do niej enzymy trawienne. W kontakcie komórki z obcą substancją lub z obcą komórką lizosomy sklejają się Błona komórkowa poprzez uwalnianie ich enzymów poza komórkę. Enzymy lizosomalne mogą również brać udział w „zaprogramowanej śmierci” własnych komórek.
Dzięki aktywności aparatu Golgiego następuje odnowa i wzrost błony plazmatycznej.
8. Rybosomy.
Rybosomy znajdują się w komórkach wszystkich organizmów. Są to mikroskopijne ciała o zaokrąglonym kształcie i średnicy 15-20 nm. Każdy rybosom składa się z dwóch cząsteczek o różnej wielkości, małej i dużej. (patrz Załącznik nr 14).
Jedna komórka zawiera wiele tysięcy rybosomów, znajdują się one albo na błonach ziarnistej retikulum endoplazmatycznego, albo leżą swobodnie w cytoplazmie.
Rybosomy składają się z białek i RNA. Funkcją rybosomów jest synteza białek. Synteza białek - trudny proces, który jest wykonywany nie przez jeden rybosom, ale przez całą grupę, obejmującą nawet kilkadziesiąt połączonych rybosomów. Ta grupa rybosomów nazywa się polisom .
Zsyntetyzowane białka są najpierw gromadzone w kanałach i jamach retikulum endoplazmatycznego, a następnie transportowane do organelli i miejsc komórkowych, gdzie są konsumowane.
Retikulum endoplazmatyczne i rybosomy (Patrz Załącznik nr 8), znajdujące się na jego membranach, stanowią pojedynczy aparat do biosyntezy i transportu białek.
9. Mitochondria.
Samopodzielające się organelle błonowe. Cytoplazma większości komórek zwierzęcych i roślinnych zawiera małe ciała (0,2-7 mikronów) - mitochondria (greckie „mitos” - nić, „chondrion” - ziarno, granulka).
Mitochondria są wyraźnie widoczne w mikroskopie świetlnym, za pomocą którego można zobaczyć ich kształt, położenie, policzyć liczbę (patrz Załącznik nr 8). Wewnętrzną strukturę mitochondriów badano za pomocą mikroskopu elektronowego.
Powłoka mitochondriów składa się z dwóch błon - zewnętrznej i wewnętrznej. Błona zewnętrzna jest gładka, nie tworzy fałd i wypustek. Przeciwnie, błona wewnętrzna tworzy liczne fałdy, które są skierowane do jamy mitochondriów. Nazywa się fałdy błony wewnętrznej cristae (łac. „crista” - grzebień, odrost) Zachodzi w nich proces utleniania kwasu mlekowego, w wyniku którego uwalniana jest energia zmagazynowana w postaci ATP.
Liczba cristae nie jest taka sama w mitochondriach różnych komórek. Może ich być od kilkudziesięciu do kilkuset, a szczególnie wiele cristae występuje w mitochondriach aktywnie funkcjonujących komórek, na przykład komórek mięśniowych.
Mitochondria nazywane są „elektrowniami” komórek, ponieważ ich główną funkcją jest synteza trójfosforanu adenozyny (ATP). Kwas ten syntetyzowany jest w mitochondriach komórek wszystkich organizmów i jest uniwersalnym źródłem energii niezbędnej do realizacji procesów życiowych komórki i całego organizmu.
Mitochondria, podobnie jak bakterie, rozmnażają się przez bezpośredni podział.
10. Centrum komórkowe.
W komórkach w pobliżu jądra znajduje się organelle, które nazywane są centrum komórkowym. Główna część centrum komórkowego składa się z dwóch małych ciał (czasami więcej) - centriole znajduje się na niewielkim obszarze zagęszczonej cytoplazmy. Każda centriola ma kształt walca o długości do 1 µm, którego ścianki tworzą mikrotubule rozmieszczone wzdłuż osi walca. Centriole zawierają białka i duża liczba RNA. Komórka ma dwie pary centrioli. W każdej parze centriole znajdują się prostopadle do siebie.
Centriole odgrywają ważną rolę w podziale komórkowym. Tworzą długie włókna białkowe, tzw. wrzeciono rozszczepienia. Podczas podziału komórki centriole rozchodzą się w kierunku jej biegunów, określając orientację wrzeciona podziałowego.
11. Plastydy.
Plastydy to błoniaste, samodzielące się organelle komórkowe. W przeciwieństwie do wszystkich omówionych powyżej organelli, plastydy występują tylko w komórkach roślinnych.
W budowie przypominają mitochondria: składają się z dwóch błon, zewnętrznej gładkiej i wewnętrznej, tworzących liczne płaskie wyrostki - tylakoidy . Wszystkie tylakoidy są ułożone jak stosy monet. Każdy stos jest wywoływany fasetowany . Pomiędzy granami znajduje się wewnętrzny płyn plastydowy tzw zrąb . Zawiera własne DNA, którego struktura przypomina bakterie.
Plastydy rozmnażają się jak bakterie przez bezpośredni podział.
Zgodnie z cechami strukturalnymi wyróżnia się trzy główne typy plastydów (Patrz Załącznik nr 15): zielony - chloroplasty; czerwony, pomarańczowy i żółty - chromoplasty; bezbarwny - leukoplasty.
a) chloroplast.
Organelle te znajdują się w komórkach liści i innych organach roślin zielonych (pędy, niedojrzałe owoce), a także w różnych algach.
Rozmiar chloroplastów wynosi 4-6 mikronów, najczęściej mają owalny kształt. U roślin wyższych jedna komórka ma zwykle kilkadziesiąt chloroplastów.
Zielony kolor chloroplasty zależy od zawartości w nich barwnika chlorofilowego.
Chloroplast to główna organella komórek roślinnych, w której zachodzi fotosynteza, czyli tworzenie substancji organicznych (węglowodanów) z substancji nieorganicznych (CO2 i H2O) przy wykorzystaniu energii światła słonecznego.
Chloroplasty są strukturalnie podobne do mitochondriów. (patrz Załącznik nr 16). Chloroplast jest oddzielony od cytoplazmy dwiema błonami - zewnętrzną i wewnętrzną. Błona zewnętrzna jest gładka, bez fałd i wypustek, a wewnętrzna tworzy wiele pofałdowanych narośli skierowanych do wnętrza chloroplastu. Dlatego duża liczba membran koncentruje się wewnątrz chloroplastu, tworząc specjalne struktury - grana. Cząsteczki chlorofilu znajdują się w błonach gran, ponieważ to tutaj zachodzi fotosynteza.
ATP jest również syntetyzowany w chloroplastach. Pomiędzy wewnętrznymi błonami chloroplastu znajduje się DNA, RNA i rybosomy. W związku z tym w chloroplastach, a także w mitochondriach, dochodzi do syntezy białka niezbędnego do działania tych organelli. Chloroplasty rozmnażają się przez podział (patrz Załącznik nr 17).
b) Chromoplasty.
Chromoplasty to plastydy zawierające tłuszczopodobne pigmenty, które barwią komórkę na żółto, pomarańczowo i czerwono. Występują w dojrzałych owocach roślin, nadając im odpowiednią barwę, a także jesienią w liściach drzew liściastych.
c) Leukoplasty.
Leukoplasty to bezbarwne plastydy. Na wewnętrznej membranie nie zawierają żadnych pigmentów. W komórce odpowiadają za syntezę i akumulację polisacharydów (skrobi). Występują w dużych ilościach w komórkach podziemnych pędów (bulwy ziemniaka, topinambur), a także w owocach i nasionach.
d) Transformacje plastydowe.
Chloroplasty mogą łatwo przerodzić się w inne rodzaje plastydów. Obserwujemy to przy żółknięciu i zaczerwienieniu dojrzewających owoców lub liści jesienią. W ciemności chloroplasty są w stanie odbarwić się, zamieniając się w leukoplasty. Jednak procesy te są nieodwracalne: leukoplasty i chromoplasty nigdy nie zamieniają się z powrotem w chloroplasty.
Jądro (jądro komórkowe) część obowiązkowa komórek wielu organizmów jednokomórkowych i wszystkich organizmów wielokomórkowych. Rozmiary od 1 mikrona (u niektórych pierwotniaków) do 1 mm (w jajach niektórych ryb i płazów). Terminu „jądro” (łac. Nucleus) użył po raz pierwszy R. Brown w 1833 r., kiedy opisał sferyczne struktury, które zaobserwował w komórkach roślinnych (patrz Załącznik nr 8).
a) otoczka jądrowa.
Wewnętrzna przestrzeń jądra komórkowego jest oddzielona od cytoplazmy błoną jądrową składającą się z dwóch błon. Błony błony jądrowej mają podobną budowę do innych składników błony komórkowej i są zbudowane według tej samej zasady: są to cienkie błony lipoproteinowe składające się z podwójnej warstwy cząsteczek lipidów, w których osadzone są cząsteczki białek.
Przestrzeń między wewnętrzną i zewnętrzną błoną jądrową nazywa się okołojądrowy.
Duża liczba rybosomów jest zwykle zlokalizowana na powierzchni zewnętrznej błony jądrowej, a czasami można zaobserwować bezpośrednie przejście tej błony do systemu kanałów ziarnistej retikulum endoplazmatycznego komórki.
Wewnętrzna błona jądrowa jest otoczona cienką włóknistą warstwą białka - blaszka jądrowa składa się z białek laminowanych. Gęsta sieć włókienek blaszki jądrowej jest w stanie zapewnić integralność jądra, nawet po rozpuszczeniu błon lipidowych otoczki jądra w eksperymencie. Z wewnątrz pętle chromatyny wypełniające jądro są przyczepione do blaszki.
Błona jądrowa ma otwory o średnicy około 90 nm, utworzone przez połączenie zewnętrznej i wewnętrznej błony jądrowej. Takie dziury w otoczce jądrowej otoczone są złożonymi strukturami białkowymi zwanymi kompleksem porów jądrowych.
Osiem podjednostek białkowych tworzących pory jądrowe znajduje się wokół perforacji otoczki jądrowej w postaci pierścieni o średnicy około 120 nm, obserwowanych pod mikroskopem elektronowym po obu stronach otoczki jądrowej. Białkowe podjednostki kompleksu porów mają wyrostki skierowane w kierunku środka porów, gdzie czasami widoczna jest „centralna granulka” o średnicy 10–40 nm.
Wielkość porów jądrowych i ich struktura są standardowe dla wszystkich komórek eukariotycznych. Liczba porów jądrowych zależy od aktywności metabolicznej komórek: im wyższy poziom procesów syntezy w komórce, tym więcej porów na jednostkę powierzchni jądra komórkowego.
W procesie transportu jądrowo-cytoplazmatycznego pory jądrowe pełnią rolę swoistego sita molekularnego, przepuszczając jony i małe cząsteczki (cukry, nukleotydy, ATP itp.) biernie, wzdłuż gradientu stężeń, i realizując aktywny selektywny transport dużych cząsteczek białek i rybonukleoprotein, czyli kwasu rybonukleinowego kompleksującego kwasy (RNA) z białkami. I tak np. białka transportowane do jądra z cytoplazmy, gdzie są syntetyzowane, muszą mieć określone sekwencje około 50 aminokwasów (tzw. sekwencje NLS), „rozpoznawane” przez kompleks porów jądrowych. W tym przypadku kompleks porów jądrowych, wykorzystując energię w postaci ATP, aktywnie przemieszcza białko z cytoplazmy do jądra.
b) Chromatyna.
Jądro komórkowe jest pojemnikiem na prawie całą informację genetyczną komórki, więc główną zawartością jądra komórkowego jest chromatyna: kompleks kwasu dezoksyrybonukleinowego (DNA) i różnych białek.
W jądrze, a zwłaszcza w chromosomach mitotycznych, DNA chromatyny jest wielokrotnie fałdowane, pakowane w specjalny sposób, aby osiągnąć wysoki stopień zagęszczanie. Przecież wszystkie długie nici DNA, których całkowita długość np. u ludzi wynosi około 164 cm, muszą być umieszczone w jądrze komórkowym, którego średnica wynosi zaledwie kilka mikrometrów. Zadanie to rozwiązuje się poprzez sekwencyjne pakowanie DNA w chromatynie za pomocą specjalnych białek.
Większość białek chromatyny to białka histonowe, które są częścią globularnych podjednostek chromatyny zwanych nukleosomy . W sumie istnieje 5 rodzajów białek histonowych.
Nukleosom to cylindryczna cząsteczka, składająca się z 8 cząsteczek histonów o średnicy około 10 nm, na których „nawinięte” są nieco mniej niż dwa zwoje cząsteczki DNA. W mikroskop elektronowy taka sztucznie zdekondensowana chromatyna wygląda jak „koraliki na sznurku”.
W żywym jądrze komórki nukleosomu są ściśle połączone ze sobą za pomocą innego białka histonowego łącznika, tworząc tzw. elementarne fibryle chromatyny o średnicy 30 nm. Inne białka o charakterze niehistonowym, które tworzą chromatynę, zapewniają dalsze zagęszczanie. tj. układanie w stosy chromatyny fibrylowej, która dociera do niej maksymalne wartości podczas podziału komórki w chromosomach mitotycznych lub mejotycznych.
W jądrze komórkowym chromatyna występuje zarówno w postaci gęstej chromatyny skondensowanej, w której gęsto upakowane są elementarne fibryle o długości 30 nm, jak iw postaci jednorodnej chromatyny rozproszonej. Stosunek ilościowy tych dwóch rodzajów chromatyny zależy od charakteru aktywności metabolicznej komórki, stopnia jej zróżnicowania. Na przykład jądra ptasich erytrocytów, w których nie zachodzą aktywne procesy replikacji i transkrypcji, zawierają praktycznie tylko gęstą, skondensowaną chromatynę.
Część chromatyny zachowuje swój zwarty, skondensowany stan przez cały cykl komórkowy – ta chromatyna to tzw heterochromatyna i różni się od euchromatyny szeregiem właściwości.
c) Replikacja i transkrypcja.
Komórki eukariotyczne zawierają zwykle kilka chromosomów (od dwóch do kilkuset), które w jądrze tracą swój zwarty kształt (w interfazie, czyli między podziałami mitotycznymi), rozluźniają się i wypełniają objętość jądra w postaci chromatyny. Pomimo stanu zdekondensowanego każdy chromosom zajmuje ściśle określoną pozycję w jądrze i jest połączony z otoczką jądrową za pomocą blaszki. ściśle ustalone wewnętrzna powierzchnia otoczki jądra takie struktury chromosomów jak centromery i telomery.
Na pewnym etapie cyklu życiowego komórki, w okresie syntezy, następuje replikacja, czyli podwojenie całego DNA jądra, a chromatyna staje się dwukrotnie większa. Białka niezbędne do tego procesu pochodzą oczywiście z cytoplazmy przez pory jądrowe. W ten sposób komórka przygotowuje się do nadchodzącego podziału komórki - mitozy, kiedy całkowity DNA w jądrze powróci do swojego pierwotnego poziomu.
Implementacja informacji genetycznej zawartej w DNA w postaci genów rozpoczyna się od transkrypcji, czyli od syntezy informacyjnego RNA (i-RNA) – dokładnych kopii genów, dzięki którym białka będą następnie budowane w cytoplazmie na rybosomach.
Proces ten zachodzi w różnych punktach objętości jądra, morfologicznie nie do odróżnienia od otaczającej chromatyny. Najczęściej można zaobserwować transkrypcję rozproszoną, tj. zdekondensowana chromatyna.
Oprócz chromatyny, która tworzy chromosomy, jądra eukariotyczne zwykle zawierają jedno lub więcej jąderek.
d) jądrowa macierz białkowa.
Aby przeprowadzić procesy replikacji, transkrypcji, a także utrzymać określoną pozycję chromosomów w objętości jądra, istnieją szkieletowe struktury białkowe zwane jądrową macierzą białkową. Taka macierz składa się z co najmniej trzech elementów morfologicznych:
obwodowa warstwa włóknista - blaszki;
wewnętrzna lub międzychromatynowa macierz jądra
i macierz jąderka.
Obserwacje pokazują, że składniki macierzy jądrowej nie są sztywnymi zamrożonymi strukturami, są dynamiczne i mogą się znacznie zmieniać w zależności od cechy funkcjonalne jądra. Wykazano, że macierz białkowa ma wiele punktów silnego wiązania z rdzeniem DNA, który z kolei ma specjalne, niezbędne do tego sekwencje nukleotydowe.
13. Jądra.
a) Liczba jąderek w komórce.
Począwszy od zielonych alg, grzybów i pierwotniaków niższych, a skończywszy na wyższe organizmy, wszystkie komórki mają obowiązkowe struktury wewnątrzjądrowe - jąderka (patrz Załącznik nr 18). Reguła ta ma dużą liczbę wyjątków, które tylko podkreślają wagę i konieczność obecności jąderka w organizmie koło życia komórki. Do takich wyjątków należą komórki rozszczepionych jaj, w których jąderka są nieobecne we wczesnych stadiach embriogenezy lub komórki, które zakończyły rozwój i mają nieodwracalną specjalizację, na przykład niektóre krwinki.
Liczba jąderek w komórce może być różna, ale ich liczba w jądrze zależy od równowagi genów w komórce. Stwierdzono, że bierze udział w tworzeniu jąderek pewne miejsca niektóre chromosomy, których połączenie z jąderkiem można dobrze prześledzić w telofazie i profazie. Takie chromosomy z reguły mają wtórne przewężenia, których strefy są miejscami rozwoju jąderek. McClintock (1934) nazwał te sekcje chromosomów „organizatory jąderkowe”.
Miejsca przewężeń wtórnych są szczególnie charakterystyczne dla lokalizacji organizatorów jąderkowych, ale te ostatnie mogą czasami znajdować się na końcach chromosomów lub w kilku miejscach wzdłuż chromosomu.
Łączna jąderka przypadające na jądro jest określane przez liczbę organizatorów jąderek i wzrasta zgodnie z ploidią jądra. Jednak często liczba jąderek na jądro jest mniej niż liczba organizatorzy jąder. Wykazano, że jąderka mogą się łączyć; ponadto czasami kilku organizatorów uczestniczy w tworzeniu jednego jąderka.
Nawet w pracach M. S. Navashin (1934) wykazano, że locus chromosomalny, który w normalne warunki tworzy duże jąderko, staje się nieaktywne, gdy po hybrydyzacji w jądrze na innym chromosomie pojawia się „silniejszy” locus. Fakt, że w pewnych warunkach aktywność niektórych organizatorów jąderek może zostać stłumiona lub aktywność innych, które wcześniej znajdowały się w stanie utajonym, ukrytym, może wzrosnąć, wskazuje, że w komórkach zachowana jest pewna równowaga ilości materiału jąderkowego , czyli innymi słowy produkty „brutto” wytwarzane przez jąderka.
Na podstawie powyższych faktów można wyciągnąć następujące wnioski:
Þ powstawanie jąderek i ich liczba są związane z aktywnością pewnych odcinków chromosomów - organizatorów jąderkowych, które są zlokalizowane przez większą część w strefach ograniczeń wtórnych;
Þ zmiany liczby jąderek w komórkach tego typu może wystąpić z powodu fuzji jąderek lub z powodu przesunięć w równowadze chromosomalnej komórki.
b) Fizjologia i chemia jąderka.
Jąderko, w porównaniu z innymi składnikami komórki, charakteryzuje się najgęstszą strukturą z największą koncentracją RNA, z niezwykle wysoka aktywność do syntezy RNA.
Stężenie RNA w jąderkach jest zawsze wyższe niż stężenie RNA w innych składnikach komórki, więc stężenie RNA w jąderku może być 2-8 razy większe niż w jądrze i 1-3 razy większe niż w jądrze cytoplazma. Stosunek stężeń RNA w jądrze, jąderku i cytoplazmie komórek wątroby myszy wynosi 1:7, 3:4, 1, w komórkach trzustki 1:9, 6:6, 6.
W jąderku nie znaleziono DNA, ale mimo to podczas badania utrwalonych komórek wokół jąderka zawsze wyróżnia się strefa chromatyny. Ta okołojądrowa chromatyna, według mikroskopii elektronowej, wydaje się być integralną częścią złożona struktura jąderko.
Jąderko jest jednym z najbardziej aktywnych miejsc w komórce do włączania prekursorów do RNA. Jąderkowy RNA jest prekursorem cytoplazmatycznego RNA.
Cytoplazmatyczny RNA jest syntetyzowany w jąderku.
c) jąderkowy RNA.
Ocenianie treść ogólna we frakcjach jąderkowych białek, RNA i DNA można zauważyć, że RNA stanowi około 10% całkowitej masy jąderka.
Ponieważ większość cytoplazmatycznego RNA to rybosomalny RNA, możemy powiedzieć, że jądrowy RNA należy do tej klasy.
Przekonanie, że jąderko jest miejscem syntezy rRNA i powstawania rybosomów, zostało potwierdzone przez fakt, że cząstki RNP zostały wyizolowane z preparatów jąderkowych, które zarówno pod względem składu RNA (zgodnie z właściwościami sedymentacyjnymi), jak i wielkości można scharakteryzować jako rybosomy lub ich prekursory o różnych współczynnikach sedymentacji.
d) jądrowy DNA.
Badania biochemiczne wykazały pewną ilość DNA w wyizolowanych jąderkach, które można zidentyfikować za pomocą chromatyny okołojądrowej lub organizatorów chromosomów jąderkowych. Zawartość DNA w wyizolowanych jąderkach wynosi 5-12% suchej masy i 6-17% całkowitego DNA jądra.
DNA organizatora jąderka jest tym samym DNA, na którym zachodzi synteza jąderka, tj. rybosomalny, RNA.
Tak więc z badań biochemicznych zrodziły się pomysły, że wiele identycznych genów do syntezy rRNA zostało zlokalizowanych na DNA w jąderku. Synteza rRNA przebiega poprzez utworzenie ogromnego prekursora i jego dalszą transformację (dojrzewanie) do krótszych cząsteczek RNA dla dużych i małych podjednostek rybosomów.
Badając jąderka oocytów traszek, badacze napotkali ciekawe zjawisko - nadmiar jąderek. Podczas wzrostu oocytu pojawia się do 1000 małych jąderek, które nie są związane z chromosomami. To właśnie te jąderka zidentyfikował O. Miller, wraz z tym wzrasta ilość rDNA na jądro oocytu. Zjawisko to zostało nazwane wzmocnienie . Polega ona na tym, że dochodzi do nadreplikacji strefy organizatora jąderka, liczne kopie odchodzą od chromosomów i stają się dodatkowo działającymi jąderkami. Taki proces jest niezbędny do zgromadzenia ogromnej (10 12) liczby rybosomów w komórce jajowej, co zapewni dalszy rozwój zarodka na wczesne stadia nawet przy braku nowej syntezy rybosomów. Nadliczbowe jąderka zanikają po dojrzewaniu komórki jajowej.
e) Ultrastruktura jąder.
podczas nauki duża liczba różne komórki U zwierząt i roślin obserwuje się włóknistą lub siatkową strukturę jąderek, zamkniętą w mniej lub bardziej gęstej rozproszonej masie. Zaproponowano nazwy dla tych części: część włóknista - jąderko i rozproszona, jednorodna część - substancja amorficzna, lub część amorficzna . Badania mikroskopowe elektronowe wykonane niemal równocześnie z tym ujawniły również włóknisto-włóknistą strukturę jąderek.
Jednak taka nitkowata struktura jąderka nie zawsze jest wyraźnie wyrażona. W niektórych komórkach pojedyncze nici nukleonemów łączą się, a jąderka mogą być całkowicie jednorodne.
Po bliższym zbadaniu jąderka można zauważyć, że główny Elementy konstrukcyjne jąderko - gęste granulki o średnicy około 15 nm i cienkie fibryle o grubości 4-8 nm. W wielu przypadkach (oocyty ryb i płazów, merystematyczne komórki roślinne) składnik włóknisty składa się w gęstą strefę środkową (rdzeń) pozbawioną ziarnistości, a ziarnistości zajmują obwodową strefę jąderka. W wielu przypadkach (na przykład komórki korzeni roślin) w tej strefie ziarnistej nie obserwuje się dodatkowej struktury.
Stwierdzono, że amorficzne regiony jąderka są heterogeniczne. W ich strukturze ujawniają się strefy o niskim zabarwieniu - centra włókniste - i otaczające je ciemniejsze obszary, które również mają strukturę włóknistą.
Oprócz tych dwóch składników jąderka w Ostatnio wiele uwagi poświęcono budowie chromatyny okołojądrowej. Ta chromatyna i wewnątrzjądrowa sieć DNA są ujednolicony system i są integralną częścią jąderka.
Granulki i część włóknista składają się z rybonukleoprotein.
Wykazano, że rDNA zawierają lekkie centra fibrylarne.