प्लांट सेल की संरचना
पादप कोशिका कमोबेश कठोर होती है कोशिका भित्तिऔर प्रोटोप्लास्ट। कोशिका भित्ति कोशिका भित्ति है और कोशिकाद्रव्य की झिल्ली. प्रोटोप्लास्ट शब्द प्रोटोप्लाज्म शब्द से आया है, जो कब कासभी जीवित चीजों को संदर्भित करता था। प्रोटोप्लास्ट एक व्यक्तिगत कोशिका का प्रोटोप्लाज्म है।
प्रोटोप्लास्ट में साइटोप्लाज्म और न्यूक्लियस होते हैं। साइटोप्लाज्म में ऑर्गेनेल (राइबोसोम, माइक्रोट्यूबुल्स, प्लास्टिड्स, माइटोकॉन्ड्रिया) और मेम्ब्रेन सिस्टम (एंडोप्लाज्मिक रेटिकुलम, डिक्टीयोसोम) होते हैं। साइटोप्लाज्म में साइटोप्लाज्मिक मैट्रिक्स (मूल पदार्थ) भी शामिल होता है जिसमें ऑर्गेनेल और मेम्ब्रेन सिस्टम डूबे रहते हैं। साइटोप्लाज्म को कोशिका भित्ति से अलग किया जाता है प्लाज्मा झिल्ली, जो एक प्राथमिक झिल्ली है। अधिकांश पशु कोशिकाओं के विपरीत, पादप कोशिकाओं में एक या एक से अधिक रिक्तिकाएँ होती हैं। ये पुटिकाएं तरल से भरी होती हैं और एक प्रारंभिक झिल्ली (टोनोप्लास्ट) से घिरी होती हैं।
एक जीवित पादप कोशिका में, मुख्य पदार्थ स्थित होता है निरंतर गति में. ऑर्गेनेल आंदोलन में शामिल होते हैं, जिसे साइटोप्लाज्म या साइक्लोसिस का प्रवाह कहा जाता है। साइक्लोसिस कोशिका में पदार्थों की आवाजाही और कोशिका और पर्यावरण के बीच उनके आदान-प्रदान की सुविधा प्रदान करता है।
प्लाज्मा झिल्ली। यह एक बाइलेयर फॉस्फोलिपिड संरचना है। पादप कोशिकाओं की विशेषता प्लाज्मा झिल्ली के अंतर्वलन से होती है।
प्लाज्मा झिल्ली निम्नलिखित कार्य करती है:
कोशिका और पर्यावरण के बीच पदार्थों के आदान-प्रदान में भाग लेता है;
कोशिका भित्ति के सेल्युलोज सूक्ष्मतंतुओं के संश्लेषण और संयोजन का समन्वय करता है;
हार्मोनल और बाहरी संकेतों को प्रसारित करता है जो सेल के विकास और भेदभाव को नियंत्रित करते हैं।
मुख्य। यह यूकेरियोटिक कोशिका के साइटोप्लाज्म में सबसे प्रमुख संरचना है। कर्नेल दो महत्वपूर्ण कार्य करता है:
कोशिका की महत्वपूर्ण गतिविधि को नियंत्रित करता है, यह निर्धारित करता है कि कौन से प्रोटीन और किस समय संश्लेषित किए जाने चाहिए;
आनुवंशिक जानकारी को संग्रहीत करता है और इसे कोशिका विभाजन के दौरान संतति कोशिकाओं तक पहुंचाता है।
एक यूकेरियोटिक कोशिका का केंद्रक दो प्राथमिक झिल्लियों से घिरा होता है जो परमाणु आवरण बनाती हैं। यह 30 से 100 एनएम के व्यास के साथ कई छिद्रों से व्याप्त है, जो केवल एक इलेक्ट्रॉन माइक्रोस्कोप में दिखाई देता है। छिद्रों की एक जटिल संरचना होती है। बाहरी झिल्लीकुछ स्थानों पर परमाणु झिल्ली को एंडोप्लाज्मिक रेटिकुलम के साथ जोड़ा जाता है। परमाणु लिफाफे को एंडोप्लाज्मिक रेटिकुलम (ईआर) के एक विशेष, स्थानीय रूप से विभेदित भाग के रूप में देखा जा सकता है।
विशेष रंगों से सना हुआ नाभिक में, कोई भी भेद कर सकता है पतले धागेऔर क्रोमैटिन और न्यूक्लियोप्लाज्म (नाभिक का मुख्य पदार्थ) की गांठ। क्रोमेटिन हिस्टोन नामक विशेष प्रोटीन से बंधे डीएनए से बना होता है। कोशिका विभाजन की प्रक्रिया में, क्रोमैटिन अधिक से अधिक संकुचित हो जाता है और गुणसूत्रों में इकट्ठा हो जाता है। डीएनए आनुवंशिक जानकारी को एनकोड करता है।
दैहिक कोशिकाओं में जीव गुणसूत्रों की संख्या में भिन्न होते हैं। उदाहरण के लिए, गोभी में - 20 गुणसूत्र होते हैं; सूरजमुखी - 34; गेहूँ - 42; मनुष्य - 46, और फ़र्न ओफ़ियोग्लोसम की प्रजातियों में से एक - 1250। सेक्स कोशिकाओं (युग्मकों) में शरीर की दैहिक कोशिकाओं के गुणसूत्रों की संख्या का केवल आधा हिस्सा होता है। युग्मकों में गुणसूत्रों की संख्या को अगुणित (एकल) कहा जाता है, दैहिक कोशिकाओं में - द्विगुणित (डबल)। जिन कोशिकाओं में गुणसूत्रों के दो से अधिक सेट होते हैं उन्हें पॉलीप्लोइड कहा जाता है।
एक प्रकाश सूक्ष्मदर्शी के तहत, गोलाकार संरचनाएं - न्यूक्लियोली - देखी जा सकती हैं। प्रत्येक नाभिक में एक या एक से अधिक नाभिक होते हैं, जो अविभाजित नाभिक में दिखाई देते हैं। राइबोसोमल आरएनए को न्यूक्लियोलस में संश्लेषित किया जाता है। आमतौर पर द्विगुणित जीवों के नाभिक में दो नाभिक होते हैं, प्रत्येक के लिए एक अगुणित सेटगुणसूत्र। न्यूक्लियोली की अपनी झिल्ली नहीं होती है। जैव रासायनिक रूप से, नाभिक की विशेषता होती है बहुत ज़्यादा गाड़ापनआरएनए, जो यहां फॉस्फोप्रोटीन से जुड़ा हुआ है। नाभिक का आकार निर्भर करता है कार्यात्मक अवस्थाकोशिकाओं। यह देखा गया है कि एक तेजी से बढ़ने वाली कोशिका में, जिसमें प्रोटीन संश्लेषण की गहन प्रक्रियाएँ हो रही होती हैं, नाभिक का आकार बढ़ जाता है। न्यूक्लियोली में, एमआरएनए और राइबोसोम उत्पन्न होते हैं, जो केवल न्यूक्लियस में सिंथेटिक कार्य करते हैं।
न्यूक्लियोप्लाज्म (कार्योप्लाज्म) एक सजातीय तरल द्वारा दर्शाया जाता है जिसमें एंजाइम सहित विभिन्न प्रोटीन घुल जाते हैं।
प्लास्टिड्स। रिक्तिकाएं, सेल्युलोज कोशिका भित्ति और प्लास्टिड पादप कोशिकाओं के विशिष्ट घटक हैं। प्रत्येक प्लास्टिड का अपना खोल होता है, जिसमें दो प्राथमिक झिल्लियाँ होती हैं। प्लास्टिड के अंदर, एक झिल्ली प्रणाली प्रतिष्ठित है और बदलती डिग्रीसजातीय पदार्थ - स्ट्रोमा। परिपक्व प्लास्टिड्स को उनमें मौजूद पिगमेंट के आधार पर वर्गीकृत किया जाता है।
क्लोरोप्लास्ट, जिसमें प्रकाश संश्लेषण होता है, में क्लोरोफिल और कैरोटीनॉयड होते हैं। आमतौर पर 4 - 5 माइक्रोन के व्यास वाली डिस्क का रूप होता है। मेसोफिल (पत्ती के मध्य) की एक कोशिका में 40-50 क्लोरोप्लास्ट हो सकते हैं; मिमी 2 शीट्स में - लगभग 500,000। साइटोप्लाज्म में, क्लोरोप्लास्ट आमतौर पर कोशिका झिल्ली के समानांतर स्थित होते हैं।
क्लोरोप्लास्ट की आंतरिक संरचना जटिल है। स्ट्रोमा को पुटिकाओं - थायलाकोइड्स के रूप में झिल्लियों की एक विकसित प्रणाली द्वारा अनुमति दी जाती है। प्रत्येक थायलाकोइड में दो झिल्ली होते हैं। थायलाकोइड्स रूप एकल प्रणाली. एक नियम के रूप में, वे ढेर में एकत्र किए जाते हैं - अनाज, सिक्कों के स्तंभों जैसा। अलग-अलग ग्रैने के थायलाकोइड्स स्ट्रोमा थायलाकोइड्स, या इंटरग्रेनल थायलाकोइड्स द्वारा परस्पर जुड़े होते हैं। क्लोरोफिल और कैरोटीनॉयड थायलाकोइड झिल्ली में एम्बेडेड होते हैं। हरे पौधों और शैवाल में क्लोरोप्लास्ट में अक्सर स्टार्च के दाने और छोटे लिपिड (वसा) की बूंदें होती हैं। स्टार्च के दाने प्रकाश संश्लेषण उत्पादों के अस्थायी भंडार होते हैं। वे केवल 24 घंटों के लिए अंधेरे में रहने वाले पौधे के क्लोरोप्लास्ट से गायब हो सकते हैं, और पौधे को प्रकाश में स्थानांतरित करने के 3-4 घंटे बाद फिर से प्रकट हो सकते हैं।
क्लोरोप्लास्ट अर्ध-स्वायत्त अंगक होते हैं और बैक्टीरिया के समान होते हैं। उदाहरण के लिए, बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम में काफी उच्च समानता होती है। वे यूकेरियोटिक राइबोसोम से छोटे होते हैं। बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम पर प्रोटीन संश्लेषण क्लोरैम्फेनिकॉल द्वारा बाधित होता है, जिसका यूकेरियोटिक कोशिकाओं पर कोई प्रभाव नहीं पड़ता है। इसके अलावा, बैक्टीरिया और क्लोरोप्लास्ट दोनों में एक समान प्रकार का न्यूक्लियॉइड होता है, जो समान तरीके से व्यवस्थित होता है। इस तथ्य के बावजूद कि क्लोरोप्लास्ट का निर्माण और उनमें निहित पिगमेंट का संश्लेषण काफी हद तक कोशिका के क्रोमोसोमल डीएनए द्वारा नियंत्रित होता है, फिर भी, अपने स्वयं के डीएनए की अनुपस्थिति में, क्लोरोप्लास्ट नहीं बनते हैं।
क्लोरोप्लास्ट को मुख्य सेलुलर अंग माना जा सकता है, क्योंकि वे सौर ऊर्जा के परिवर्तन की श्रृंखला में पहले हैं, जिसके परिणामस्वरूप मानवता को भोजन और ईंधन दोनों प्राप्त होते हैं। क्लोरोप्लास्ट में न केवल प्रकाश संश्लेषण होता है। वे अमीनो एसिड के संश्लेषण में भी शामिल हैं और वसायुक्त अम्ल, स्टार्च के अस्थायी भंडार के भंडारण के रूप में काम करते हैं।
क्रोमोप्लास्ट रंजित प्लास्टिड हैं। विविध रूप में, उनके पास क्लोरोफिल नहीं है, लेकिन कैरोटीनॉयड को संश्लेषित और संचित करते हैं, जो फूलों, पुरानी पत्तियों, फलों और जड़ों को पीला, नारंगी, लाल रंग देते हैं। क्रोमोप्लास्ट क्लोरोप्लास्ट से विकसित हो सकते हैं, जो इस मामले में क्लोरोफिल और आंतरिक झिल्ली संरचनाओं को खो देते हैं, कैरोटीनॉयड जमा करते हैं। यह कई फलों के पकने के दौरान होता है। क्रोमोप्लास्ट कीड़ों और अन्य जानवरों को आकर्षित करते हैं जिनके साथ वे विकसित हुए हैं।
ल्यूकोप्लास्ट गैर-रंजित प्लास्टिड हैं। उनमें से कुछ स्टार्च (अमाइलोप्लास्ट) को संश्लेषित करते हैं, अन्य बनाने में सक्षम हैं विभिन्न पदार्थ, लिपिड और प्रोटीन सहित। प्रकाश में, ल्यूकोप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं।
प्रोप्लास्टिड्स छोटे, रंगहीन या हल्के हरे रंग के, अविभाजित प्लास्टिड्स होते हैं जो जड़ों और अंकुरों की मेरिस्टेमेटिक (विभाजित) कोशिकाओं में पाए जाते हैं। वे अन्य, अधिक विभेदित प्लास्टिड्स - क्लोरोप्लास्ट्स, क्रोमोप्लास्ट्स और एमिनोप्लास्ट्स के पूर्ववर्ती हैं। यदि प्रकाश की कमी के कारण प्रोटोप्लास्टिड्स के विकास में देरी हो रही है, तो उनमें एक या एक से अधिक प्रोलामेलर निकाय दिखाई दे सकते हैं, जो ट्यूबलर झिल्ली के अर्ध-क्रिस्टलीय संचय हैं। प्रोलामेलर बॉडी वाले प्लास्टिड्स को इटियोप्लास्ट्स कहा जाता है। प्रकाश में, एटिओप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं, जबकि प्रोलामेलर निकायों की झिल्लियां थायलाकोइड्स बनाती हैं। इटियोप्लास्ट पौधों की पत्तियों में बनते हैं जो अंधेरे में होते हैं। बीज भ्रूण के प्रोटोप्लास्ट पहले इटियोप्लास्ट में बदलते हैं, जिससे क्लोरोप्लास्ट प्रकाश में विकसित होते हैं। प्लास्टिड्स को एक प्रकार से दूसरे प्रकार में अपेक्षाकृत आसान संक्रमणों की विशेषता है। प्लास्टिड्स, बैक्टीरिया की तरह, दो में विभाजित करके प्रजनन करते हैं। मेरिस्टेमेटिक कोशिकाओं में, प्रोटोप्लास्टिड्स के विभाजन का समय लगभग कोशिका विभाजन के समय के साथ मेल खाता है। हालांकि, परिपक्व कोशिकाओं में के सबसेपरिपक्व प्लास्टिड्स के विभाजन के परिणामस्वरूप प्लास्टिड बनता है।
माइटोकॉन्ड्रिया। क्लोरोप्लास्ट की तरह, माइटोकॉन्ड्रिया दो प्राथमिक झिल्लियों से घिरा होता है। आंतरिक झिल्ली कई सिलवटों और प्रोट्रूशियंस - cristae का निर्माण करती है, जो माइटोकॉन्ड्रिया की आंतरिक सतह को काफी बढ़ा देती है। वे प्लास्टिड्स से काफी छोटे हैं, व्यास में लगभग 0.5 माइक्रोन और लंबाई और आकार में परिवर्तनशील हैं।
माइटोकॉन्ड्रिया में, श्वसन की प्रक्रिया होती है, जिसके परिणामस्वरूप कार्बनिक अणु ऊर्जा की रिहाई के साथ टूट जाते हैं और एटीपी अणुओं में स्थानांतरित हो जाते हैं, जो सभी यूकेरियोटिक कोशिकाओं का मुख्य ऊर्जा भंडार है। अधिकांश पादप कोशिकाओं में सैकड़ों या हजारों माइटोकॉन्ड्रिया होते हैं। एक सेल में उनकी संख्या सेल की एटीपी की आवश्यकता से निर्धारित होती है। माइटोकॉन्ड्रिया निरंतर गति में हैं, कोशिका के एक भाग से दूसरे भाग में जाते हैं, एक दूसरे के साथ विलय और विभाजित होते हैं। माइटोकॉन्ड्रिया आमतौर पर वहां इकट्ठा होते हैं जहां ऊर्जा की जरूरत होती है। यदि प्लाज़्मा झिल्ली सक्रिय रूप से कोशिका से कोशिका में पदार्थों को स्थानांतरित करती है, तो माइटोकॉन्ड्रिया झिल्ली की सतह के साथ स्थित होते हैं। मोटिव एककोशिकीय शैवाल में, माइटोकॉन्ड्रिया फ्लैगेल्ला के आधार पर जमा होता है, जो उनके आंदोलन के लिए आवश्यक ऊर्जा की आपूर्ति करता है।
माइटोकॉन्ड्रिया, प्लास्टिड्स की तरह, अर्ध-स्वायत्त अंग होते हैं जिनमें अपने स्वयं के प्रोटीन के संश्लेषण के लिए आवश्यक घटक होते हैं। आंतरिक झिल्ली एक द्रव मैट्रिक्स को घेरती है जिसमें प्रोटीन, आरएनए, डीएनए, बैक्टीरिया जैसे राइबोसोम और विभिन्न विलेय होते हैं। डीएनए एक या एक से अधिक न्यूक्लियॉइड्स में स्थित गोलाकार अणुओं के रूप में मौजूद होता है।
माइटोकॉन्ड्रिया और यूकेरियोटिक कोशिकाओं के क्लोरोप्लास्ट के साथ बैक्टीरिया की समानता के आधार पर, यह माना जा सकता है कि माइटोकॉन्ड्रिया और क्लोरोप्लास्ट बैक्टीरिया से उत्पन्न हुए हैं जो बड़े हेटरोट्रॉफ़िक कोशिकाओं में "शरण" पाए गए - यूकेरियोट्स के अग्रदूत।
माइक्रोबॉडीज। प्लास्टिड्स और माइटोकॉन्ड्रिया के विपरीत, जो दो झिल्लियों द्वारा सीमांकित होते हैं, माइक्रोबॉडी एक झिल्ली से घिरे गोलाकार अंग होते हैं। माइक्रोबॉडी में एक दानेदार (दानेदार) सामग्री होती है, कभी-कभी उनमें क्रिस्टलीय प्रोटीन का समावेश भी होता है। माइक्रोबॉडी एंडोप्लाज्मिक रेटिकुलम के एक या दो क्षेत्रों से जुड़े होते हैं।
कुछ माइक्रोबॉडीज, जिन्हें प्रॉक्सिसोम कहा जाता है, खेलते हैं महत्वपूर्ण भूमिकाग्लाइकोलिक एसिड के चयापचय में, जो सीधे प्रकाश श्वसन से संबंधित है। हरी पत्तियों में, वे माइटोकॉन्ड्रिया और क्लोरोप्लास्ट से जुड़े होते हैं। अन्य माइक्रोबॉडीज, जिन्हें ग्लायॉक्सिसोम कहा जाता है, में वसा को कार्बोहाइड्रेट में बदलने के लिए आवश्यक एंजाइम होते हैं। यह कई बीजों में अंकुरण के दौरान होता है।
रिक्तिकाएं एक झिल्ली द्वारा परिसीमित कोशिका के क्षेत्र हैं, जो तरल - कोशिका रस से भरी होती हैं। वे एक टोनोप्लास्ट (वैक्यूलर मेम्ब्रेन) से घिरे होते हैं।
एक युवा पादप कोशिका में कई छोटे रिक्तिकाएँ होती हैं, जो कोशिका आयु के रूप में एक बड़ी रसधानी में विलीन हो जाती हैं। एक परिपक्व कोशिका में, इसकी मात्रा का 90% तक रिक्तिका द्वारा कब्जा किया जा सकता है। इस मामले में, साइटोप्लाज्म कोशिका झिल्ली को एक पतली परिधीय परत के रूप में दबाया जाता है। कोशिका आकार में वृद्धि मुख्य रूप से रसधानी की वृद्धि के कारण होती है। नतीजतन, टर्गर दबाव उत्पन्न होता है और ऊतक लोच बनाए रखा जाता है। यह रिक्तिका और टोनोप्लास्ट के मुख्य कार्यों में से एक है।
रस का मुख्य घटक पानी है, बाकी पौधे के प्रकार और उसकी शारीरिक अवस्था के आधार पर भिन्न होता है। रिक्तिका में लवण, शर्करा, कम अक्सर प्रोटीन होते हैं। टोनोप्लास्ट रिक्तिका में कुछ आयनों के परिवहन और संचय में सक्रिय भूमिका निभाता है। सेल सैप में आयनों की सांद्रता पर्यावरण में इसकी सांद्रता से काफी अधिक हो सकती है। पर उच्च सामग्रीकुछ पदार्थ रिक्तिका में क्रिस्टल बनाते हैं। सबसे आम कैल्शियम ऑक्सालेट क्रिस्टल हैं अलग आकार.
रिक्तिकाएं चयापचय उत्पादों (चयापचय) के संचय के स्थान हैं। ये प्रोटीन, अम्ल और यहां तक कि मनुष्य के लिए विषाक्त पदार्थ (एल्कलॉइड) भी हो सकते हैं। पिगमेंट अक्सर जमा होते हैं। नीला, बैंगनी, बैंगनी, गहरा लाल, किरमिजी पौधे की कोशिकाओं को एंथोसायनिन समूह से वर्णक देते हैं। अन्य पिगमेंट के विपरीत, वे पानी में अच्छी तरह से घुल जाते हैं और सेल सैप में पाए जाते हैं। वे कई सब्जियों (मूली, शलजम, गोभी), फल (अंगूर, आलूबुखारा, चेरी), फूल (कॉर्नफ्लॉवर, जेरेनियम, डेल्फीनियम, गुलाब, peonies) के लाल और नीले रंग का निर्धारण करते हैं। कभी-कभी ये वर्णक पत्तियों में क्लोरोफिल को ढक लेते हैं, उदाहरण के लिए, सजावटी लाल मेपल में। एंथोसायनिन रंग शरद ऋतु के पत्ते चमकदार लाल रंग. वे ठंडी धूप के मौसम में बनते हैं, जब पत्तियों में क्लोरोफिल का संश्लेषण बंद हो जाता है। पत्तियों में, जब एंथोसायनिन नहीं बनते हैं, क्लोरोफिल के विनाश के बाद, क्लोरोप्लास्ट के पीले-नारंगी कैरोटीनॉयड ध्यान देने योग्य हो जाते हैं। ठंडे, स्पष्ट शरद ऋतु में पत्तियाँ सबसे चमकीले रंग की होती हैं।
कोशिका में उनके घटकों के संचलन में, रिक्तिकाएं मैक्रोमोलेक्युलस के विनाश में शामिल होती हैं। राइबोसोम, माइटोकॉन्ड्रिया, प्लास्टिड, रिक्तिका में गिरकर नष्ट हो जाते हैं। इस पाचन गतिविधि के अनुसार, उनकी तुलना लाइसोसोम - पशु कोशिकाओं के अंग से की जा सकती है।
एंडोप्लाज्मिक रेटिकुलम (रेटिकुलम) से रिक्तिकाएं बनती हैं
राइबोसोम। छोटे कण (17 - 23nm), जिसमें प्रोटीन और RNA की लगभग समान मात्रा होती है। राइबोसोम में, अमीनो एसिड मिलकर प्रोटीन बनाते हैं। सक्रिय चयापचय वाले कोशिकाओं में उनमें से अधिक हैं। राइबोसोम कोशिका के साइटोप्लाज्म में स्वतंत्र रूप से स्थित होते हैं या एंडोप्लाज्मिक रेटिकुलम (80S) से जुड़े होते हैं। वे नाभिक (80S), माइटोकॉन्ड्रिया (70S), प्लास्टिड्स (70S) में भी पाए जाते हैं।
राइबोसोम एक जटिल बना सकते हैं जिस पर समान पॉलीपेप्टाइड्स का एक साथ संश्लेषण होता है, जिसके बारे में जानकारी एक अणु और आरएनए से ली जाती है। इस तरह के एक जटिल को पॉलीराइबोसोम (पॉलीसोम) कहा जाता है। बड़ी मात्रा में प्रोटीन को संश्लेषित करने वाली कोशिकाओं में पॉलीसोम्स की एक व्यापक प्रणाली होती है जो अक्सर जुड़ी होती है बाहरी सतहनाभिक के गोले।
अन्तः प्रदव्ययी जलिका। यह अनिश्चितकालीन सीमा की एक जटिल त्रि-आयामी झिल्ली प्रणाली है। ईआर के संदर्भ में उनके बीच एक संकीर्ण पारदर्शी स्थान के साथ दो प्राथमिक झिल्लियों की तरह दिखता है। ईआर का आकार और लंबाई सेल प्रकार, इसकी चयापचय गतिविधि और भेदभाव के चरण पर निर्भर करती है। कोशिकाओं में जो प्रोटीन को स्रावित या संग्रहीत करते हैं, ईआर को सपाट थैली या सिस्टर्न के आकार का होता है, जिसकी बाहरी सतह पर कई राइबोसोम जुड़े होते हैं। इस रेटिकुलम को रफ एंडोप्लाज्मिक रेटिकुलम कहा जाता है। एक चिकनी ईआर आमतौर पर आकार में ट्यूबलर होती है। खुरदरी और चिकनी अंतर्द्रव्यी जालिका एक ही कोशिका में मौजूद हो सकती है। एक नियम के रूप में, उनके बीच कई संबंध हैं।
एंडोप्लाज्मिक रेटिकुलम सेल की संचार प्रणाली के रूप में कार्य करता है। वह से जुड़ा हुआ है बाहरी आवरणगुठली। वास्तव में, ये दो संरचनाएं एकल झिल्ली प्रणाली बनाती हैं। जब कोशिका विभाजन के दौरान परमाणु लिफाफा फट जाता है, तो इसके टुकड़े ER अंशों के समान हो जाते हैं। एंडोप्लाज्मिक रेटिकुलम पदार्थों के परिवहन के लिए एक प्रणाली है: प्रोटीन, लिपिड, कार्बोहाइड्रेट, कोशिका के विभिन्न भागों में। पड़ोसी कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम साइटोप्लाज्मिक स्ट्रैंड्स - प्लास्मोडेस्मेटा - से जुड़े होते हैं जो कोशिका झिल्ली से गुजरते हैं।
एंडोप्लाज्मिक रेटिकुलम संश्लेषण का मुख्य स्थल है कोशिका की झिल्लियाँ. कुछ पादप कोशिकाओं में, रसधानियों और सूक्ष्म निकायों की झिल्लियों, तानाशाहों के गढ्ढों का निर्माण होता है।
गॉल्जीकाय। इस शब्द का प्रयोग एक सेल में सभी तानाशाही, या गोल्गी निकायों को संदर्भित करने के लिए किया जाता है। डिक्टियोसोम फ्लैट, डिस्क के आकार के वेसिकल्स या सिस्टर्न के समूह होते हैं जो शाखा में होते हैं जटिल सिस्टमनलिकाएं। उच्च पौधों में डिक्टायोसोम में एक साथ एकत्रित 4 - 8 कुंड होते हैं।
आमतौर पर, टैंकों के एक पैकेट में, एक विकासशील और परिपक्व पक्ष प्रतिष्ठित होता है। बनने वाले गढ्ढों की झिल्लियां संरचना में ईआर की झिल्लियों से मिलती-जुलती हैं, और परिपक्व हौदों की झिल्लियां प्लाज्मा झिल्ली से मिलती-जुलती हैं।
डिक्टायोसोम स्राव में शामिल होते हैं और अधिकांश उच्च पौधों में कोशिका झिल्लियों के निर्माण में शामिल होते हैं। डिक्टायोसोम द्वारा संश्लेषित कोशिका भित्ति पॉलीसेकेराइड पुटिकाओं में जमा हो जाते हैं, जो तब परिपक्व होने वाले सिस्टर्न से अलग हो जाते हैं। ये स्रावी पुटिकाएं सीएमपी के साथ माइग्रेट और फ्यूज हो जाती हैं; जबकि उनमें निहित पॉलीसेकेराइड कोशिका झिल्ली में निर्मित होते हैं। कुछ पदार्थ जो तानाशाहों में जमा होते हैं, अन्य संरचनाओं में बनते हैं, उदाहरण के लिए, ईआर में, और फिर तानाशाहों में ले जाया जाता है, जहां स्राव से पहले उन्हें संशोधित (संशोधित) किया जाता है। उदाहरण के लिए, ग्लाइकोप्रोटीन एक महत्वपूर्ण हैं निर्माण सामग्रीकोशिका झिल्ली। प्रोटीन भाग को किसी न किसी ईआर के पॉलीसोम द्वारा संश्लेषित किया जाता है, कार्बोहाइड्रेट भाग को डिक्टायोसोम में संश्लेषित किया जाता है, जहां दोनों भाग ग्लाइकोप्रोटीन बनाने के लिए संयोजित होते हैं।
झिल्लियां गतिशील, मोबाइल संरचनाएं हैं जो लगातार अपना आकार और क्षेत्र बदलती रहती हैं। एंडोप्लाज्मिक सिस्टम की अवधारणा झिल्ली की गतिशीलता पर आधारित है। इस अवधारणा के अनुसार, माइटोकॉन्ड्रिया और प्लास्टिड्स की झिल्लियों को छोड़कर, साइटोप्लाज्म की आंतरिक झिल्लियां एक ही पूरी होती हैं और एंडोप्लाज्मिक रेटिकुलम से उत्पन्न होती हैं। मध्यवर्ती पुटिका चरण के माध्यम से एंडोप्लाज्मिक रेटिकुलम से डिक्टायोसोम के नए कुंड बनते हैं, और स्रावी पुटिकाएं जो डिक्टीयोसोम से अलग होती हैं, अंततः प्लाज्मा झिल्ली के निर्माण में योगदान करती हैं। इस प्रकार, एंडोप्लाज्मिक रेटिकुलम और डिक्टियोसोम एक कार्यात्मक संपूर्ण बनाते हैं जिसमें डिक्टियोसोम एक भूमिका निभाते हैं मध्यवर्ती संरचनाएंप्लाज्मा के समान झिल्ली में एंडोप्लाज्मिक रेटिकुलम के समान झिल्ली को परिवर्तित करने की प्रक्रिया में। उन ऊतकों में जिनकी कोशिकाएं बढ़ती हैं और कमजोर रूप से विभाजित होती हैं, झिल्ली के घटकों को लगातार अद्यतन किया जाता है।
माइक्रोट्यूबुल्स लगभग सभी यूकेरियोटिक कोशिकाओं में पाए जाते हैं। वे लगभग 24 एनएम के व्यास के साथ बेलनाकार संरचनाएं हैं। इनकी लंबाई अलग-अलग होती है। प्रत्येक नलिका ट्यूबुलिन नामक प्रोटीन की उपइकाइयों से बनी होती है। उपइकाइयां केंद्रीय गुहा के चारों ओर 13 अनुदैर्ध्य तंतु बनाती हैं। सूक्ष्मनलिकाएं गतिशील संरचनाएं हैं, वे नियमित रूप से नष्ट हो जाती हैं और कोशिका चक्र के कुछ चरणों में बनती हैं। में इकट्ठे होते हैं विशेष स्थानसूक्ष्मनलिका आयोजन केंद्र कहलाते हैं। पादप कोशिकाओं में, उनके पास कमजोर व्यक्त अनाकार संरचना होती है।
सूक्ष्मनलिकाएं के कार्य: कोशिका झिल्ली के निर्माण में भाग लेते हैं; बनाने वाली झिल्ली के लिए तानाशाहों के सीधे बुलबुले, जैसे धुरी के धागे जो एक विभाजित कोशिका में बनते हैं; सेल प्लेट (बेटी कोशिकाओं के बीच प्रारंभिक सीमा) के निर्माण में भूमिका निभाते हैं। इसके अलावा, सूक्ष्मनलिकाएं महत्वपूर्ण घटकफ्लैगेल्ला और सिलिया, जिसके आंदोलन में वे महत्वपूर्ण भूमिका निभाते हैं।
माइक्रोफिलामेंट्स, जैसे सूक्ष्मनलिकाएं, लगभग सभी यूकेरियोटिक कोशिकाओं में पाए जाते हैं। वे 5-7 एनएम मोटे लंबे तंतु होते हैं, जिनमें सिकुड़ा हुआ प्रोटीन एक्टिन होता है। उच्च पौधों की कई कोशिकाओं में माइक्रोफ़िल्मेंट्स के बंडल पाए जाते हैं। जाहिर है, वे साइटोप्लाज्म की धाराओं में एक महत्वपूर्ण भूमिका निभाते हैं। माइक्रोफ़िल्मेंट्स, सूक्ष्मनलिकाएं के साथ मिलकर एक लचीला नेटवर्क बनाते हैं जिसे साइटोस्केलेटन कहा जाता है।
ग्राउंड पदार्थ को लंबे समय से एक सजातीय (सजातीय) प्रोटीन युक्त समाधान माना जाता है जिसमें कुछ या कोई संरचना नहीं होती है। हालांकि, वर्तमान में, एक उच्च-वोल्टेज इलेक्ट्रॉन माइक्रोस्कोप का उपयोग करके, यह स्थापित किया गया है कि जमीनी पदार्थ एक त्रि-आयामी जाली है जो पतली (3–6 एनएम व्यास) किस्में से निर्मित होती है जो पूरे सेल को भरती है। सूक्ष्मनलिकाएं और माइक्रोफ़िल्मेंट्स सहित साइटोप्लाज्म के अन्य घटक इस माइक्रोट्रैब्युलर जाली से निलंबित हैं।
माइक्रोट्राबेकुलर संरचना प्रोटीन स्ट्रैंड्स की एक जाली है, जिसके बीच का स्थान पानी से भरा होता है। पानी के साथ, जाली में जेल की स्थिरता होती है; जेल जिलेटिनस निकायों की तरह दिखता है।
ऑर्गेनेल माइक्रोट्राबेकुलर जाली से जुड़े होते हैं। जाली के बीच एक लिंक प्रदान करता है अलग हिस्सेकोशिकाओं और इंट्रासेल्युलर परिवहन को निर्देशित करता है।
लिपिड की बूंदें गोलाकार संरचनाएं होती हैं जो साइटोप्लाज्म को ग्रैन्युलैरिटी देती हैं पौधा कोशाणुएक प्रकाश खुर्दबीन के नीचे। इलेक्ट्रॉन माइक्रोग्राफ पर, वे अनाकार दिखते हैं। प्लास्टिड्स में बहुत समान लेकिन छोटी बूंदें पाई जाती हैं।
लिपिड बूँदें, गलती से उन्हें ऑर्गेनेल समझकर, उन्हें स्फेरोसोम्स कहते हैं और मानते हैं कि वे एक या दो परत वाली झिल्ली से घिरे हुए हैं। हालाँकि, हाल के आंकड़े बताते हैं कि लिपिड बूंदों में झिल्ली नहीं होती है, लेकिन प्रोटीन के साथ लेपित हो सकते हैं।
एर्गैस्टिक पदार्थ प्रोटोप्लास्ट के "निष्क्रिय उत्पाद" हैं: आरक्षित पदार्थ या अपशिष्ट उत्पाद। वे प्रकट हो सकते हैं और अंदर गायब हो सकते हैं विभिन्न अवधिकोशिका चक्र। स्टार्च अनाज, क्रिस्टल, एंथोसायनिन पिगमेंट और लिपिड बूंदों को छोड़कर। इनमें रेजिन, गोंद, टैनिन और प्रोटीन शामिल हैं। एर्गैस्टिक पदार्थ कोशिका झिल्ली का हिस्सा होते हैं, साइटोप्लाज्म और ऑर्गेनेल का मुख्य पदार्थ, जिसमें रिक्तिकाएं भी शामिल हैं।
फ्लैगेल्ला और सिलिया पतली, बालों जैसी संरचनाएं हैं जो कई यूकेरियोटिक कोशिकाओं की सतह से फैली हुई हैं। उनका एक स्थिर व्यास है, लेकिन लंबाई 2 से 150 माइक्रोन तक भिन्न होती है। परंपरागत रूप से, उनमें से लंबे और कुछ को फ्लैगेल्ला कहा जाता है, और छोटे और अधिक कई को सिलिया कहा जाता है। इन दो प्रकार की संरचनाओं के बीच कोई स्पष्ट अंतर नहीं है, इसलिए फ्लैगेलम शब्द का उपयोग दोनों को संदर्भित करने के लिए किया जाता है।
कुछ शैवालों तथा कवकों में कशाभिका (Flagella) चलन अंग होते हैं, जिनकी सहायता से ये जल में गति करते हैं। पौधों में (उदाहरण के लिए, काई, लिवरवॉर्ट्स, फ़र्न, कुछ जिम्नोस्पर्म), केवल जर्म कोशिकाओं (युग्मक) में फ्लैगेल्ला होता है।
प्रत्येक फ्लैगेलम का एक विशिष्ट संगठन होता है। सूक्ष्मनलिकाएं के 9 जोड़े की एक बाहरी रिंग फ्लैगेलम के केंद्र में स्थित दो अतिरिक्त सूक्ष्मनलिकाएं को घेरे हुए है। एंजाइम युक्त "हैंडल" प्रत्येक बाहरी जोड़े के एक सूक्ष्मनलिका से फैलता है। यह यूकेरियोटिक जीवों में सभी कशाभों में पाया जाने वाला बुनियादी 9+2 संगठन पैटर्न है। यह माना जाता है कि फ्लैगेल्ला की गति सूक्ष्मनलिकाएं के फिसलने पर आधारित होती है, जबकि सूक्ष्मनलिकाएं के बाहरी जोड़े बिना संकुचन के एक दूसरे के साथ चलते हैं। एक दूसरे के सापेक्ष सूक्ष्मनलिकाएं के जोड़े के फिसलने से फ्लैगेलम का स्थानीय झुकाव होता है।
फ्लैगेल्ला साइटोप्लाज्मिक बेलनाकार संरचनाओं से "बढ़ता है" जिसे बेसल बॉडी कहा जाता है, जो फ्लैगेलम का बेसल भाग भी बनाता है। बेसल निकायों के पास है आंतरिक संरचना, एक फ्लैगेलम की संरचना जैसा दिखता है, सिवाय इसके कि बाहरी नलिकाएं ट्रिपल में एकत्र की जाती हैं, न कि जोड़े में, और केंद्रीय नलिकाएं अनुपस्थित हैं।
कोशिका भित्ति। कोशिका भित्ति प्रोटोप्लास्ट के आकार को परिसीमित करती है और रिक्तिका द्वारा पानी के अवशोषण के कारण इसके टूटने को रोकती है।
कोशिका भित्ति में विशिष्ट कार्य होते हैं जो न केवल कोशिका और उस ऊतक के लिए महत्वपूर्ण होते हैं जिसमें कोशिका स्थित होती है, बल्कि पूरे पौधे के लिए भी महत्वपूर्ण होती है। कोशिका भित्ति पदार्थों के अवशोषण, परिवहन और उत्सर्जन में एक आवश्यक भूमिका निभाती है, और इसके अलावा, लाइसोसोमल या पाचन गतिविधि उनमें केंद्रित हो सकती है।
सेल की दीवार के घटक। कोशिका भित्ति का सबसे विशिष्ट घटक सेल्युलोज है, जो काफी हद तक इसकी वास्तुकला को निर्धारित करता है। सेल्युलोज अणु अंत से अंत तक जुड़े ग्लूकोज अणुओं को दोहराते हुए बने होते हैं। लंबे पतले सेल्युलोज अणुओं को 10 - 25 एनएम की मोटाई के साथ माइक्रोफाइब्रिल में जोड़ा जाता है। माइक्रोफ़ाइब्रिल्स आपस में जुड़ते हैं और पतले धागे बनाते हैं, जो बदले में एक दूसरे के चारों ओर लपेटे जा सकते हैं, जैसे कि रस्सी में किस्में। ऐसी प्रत्येक "रस्सी", या मैक्रोफिब्रिल की मोटाई लगभग 0.5 माइक्रोन होती है, जो 4 माइक्रोन की लंबाई तक पहुंचती है। मैक्रोफाइब्रिल समान आकार के स्टील के तार जितने मजबूत होते हैं।
सेल की दीवार का सेल्यूलोज फ्रेम इसके साथ जुड़े सेल्यूलोज मैट्रिक्स अणुओं से भरा होता है। इसमें पॉलीसेकेराइड होते हैं जिन्हें हेमिकेलुलोज कहा जाता है, और पेक्टिन, या पेक्टिन, रासायनिक रूप से हेमिकेलुलोज के बहुत करीब होते हैं।
कोशिका भित्ति का एक अन्य घटक, लिग्निन, सेल्युलोज के बाद पादप कोशिकाओं में सबसे प्रचुर मात्रा में पाया जाने वाला बहुलक है। लिग्निन दीवार की कठोरता को बढ़ाता है और आमतौर पर उन कोशिकाओं में पाया जाता है जो एक सहायक या यांत्रिक कार्य करती हैं।
क्यूटिन, सुबेरिन, मोम - आमतौर पर पौधों के सुरक्षात्मक ऊतकों के गोले में जमा होते हैं। क्यूटिन, उदाहरण के लिए, एपिडर्मिस की कोशिका झिल्लियों में पाया जाता है, और सुबेरिन द्वितीयक सुरक्षात्मक ऊतक, कॉर्क में पाया जाता है। दोनों पदार्थ वैक्स के संयोजन में पाए जाते हैं और पौधे से अत्यधिक पानी के नुकसान को रोकते हैं।
कोशिका भित्ति की परतें। पादप कोशिकाओं की दीवार की मोटाई व्यापक रूप से भिन्न होती है, जो पादप संरचना में कोशिकाओं की भूमिका और स्वयं कोशिका की आयु पर निर्भर करती है। अंतर्गत इलेक्ट्रॉन सूक्ष्मदर्शीप्लांट सेल वॉल में दो परतें दिखाई देती हैं: मीडियन लैमिना (जिसे इंटरसेलुलर पदार्थ भी कहा जाता है), और प्राइमरी सेल वॉल। कई कोशिकाएं एक और परत बिछाती हैं - द्वितीयक कोशिका भित्ति. मीडियन प्लेट आसन्न कोशिकाओं की प्राथमिक दीवारों के बीच स्थित होती है। द्वितीयक दीवार, यदि कोई हो, प्राथमिक सेल दीवार की आंतरिक सतह पर सेल प्रोटोप्लास्ट द्वारा जमा की जाती है।
बीच की थाली। मीडियन लैमिना में मुख्य रूप से पेक्टिन होता है। जहां एक कोशिका भित्ति उत्पन्न होनी चाहिए, दो नवगठित कोशिकाओं के बीच, एंडोप्लाज़मिक रेटिकुलम के नलिकाओं का एक घना जाल और गोल्गी तंत्र (डिक्टायोसोम) का कुंड पहले नोट किया जाता है। फिर इस जगह में पेक्टिन (पॉलीसेकेराइड से) से भरे बुलबुले दिखाई देते हैं। इन बुलबुलों को गॉल्जी उपकरण की टंकियों से अलग किया जाता है। प्रारंभिक कोशिका भित्ति में विभिन्न पॉलीसेकेराइड होते हैं, जिनमें से मुख्य हैं पेक्टिन और हेमिकेलुलोज। बाद में, इसमें सघन पदार्थ - सेल्युलोज और लिग्निन होते हैं।
प्राथमिक कोशिका भित्ति। यह सेल्युलोज आवरण की एक परत है जो कोशिका वृद्धि से पहले या उसके दौरान जमा होती है। सेलूलोज़, हेमिसेल्यूलोज़ और पेक्टिन के अलावा, प्राथमिक झिल्ली में ग्लाइकोप्रोटीन होता है। प्राथमिक गोले लिग्निफाई कर सकते हैं। पेक्टिन घटक नमनीयता प्रदान करता है, जो प्राथमिक खोल को जड़, तना या पत्ती के रूप में विस्तारित करने की अनुमति देता है।
सक्रिय रूप से विभाजित होने वाली कोशिकाओं (प्रकाश संश्लेषण, श्वसन और स्राव की प्रक्रियाओं में शामिल अधिकांश परिपक्व कोशिकाओं) में प्राथमिक झिल्ली होती है। एक प्राथमिक झिल्ली और एक जीवित प्रोटोप्लास्ट वाली ऐसी कोशिकाएं खोने में सक्षम होती हैं विशेषता आकार, विभाजित करें और एक नए सेल प्रकार में अंतर करें। वे पौधों में घाव भरने और ऊतक पुनर्जनन में शामिल हैं।
प्राथमिक कोशिका झिल्लियां अपनी पूरी लंबाई में मोटाई में एक समान नहीं होती हैं, लेकिन पतले खंड होते हैं, जिन्हें प्राथमिक छिद्र क्षेत्र कहा जाता है। साइटोप्लाज्म, या प्लास्मोडेस्माटा की किस्में, जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ती हैं, आमतौर पर प्राथमिक छिद्र क्षेत्रों से गुजरती हैं।
द्वितीयक कोशिका भित्ति। इस तथ्य के बावजूद कि कई पादप कोशिकाओं में केवल एक प्राथमिक दीवार होती है, कुछ में, प्रोटोप्लास्ट कोशिका के केंद्र की ओर एक द्वितीयक दीवार जमा करता है। यह आमतौर पर कोशिका वृद्धि की समाप्ति के बाद होता है और प्राथमिक झिल्ली का क्षेत्र अब नहीं बढ़ता है। इस कारण से, द्वितीयक खोल प्राथमिक से भिन्न होता है। माध्यमिक झिल्लियों की विशेष रूप से विशेष कोशिकाओं द्वारा आवश्यकता होती है जो पौधे को मजबूत करती हैं और पानी का संचालन करती हैं। द्वितीयक झिल्ली के निक्षेपण के बाद, इन कोशिकाओं के प्रोटोप्लास्ट, एक नियम के रूप में, मर जाते हैं। माध्यमिक गोले में प्राथमिक की तुलना में अधिक सेल्यूलोज होता है, और उनमें पेक्टिन पदार्थ और ग्लाइकोप्रोटीन अनुपस्थित होते हैं। द्वितीयक झिल्ली को फैलाना मुश्किल होता है, इसके मैट्रिक्स में हेमिकेलुलोज होता है।
द्वितीयक खोल में, तीन परतों को प्रतिष्ठित किया जा सकता है - बाहरी, मध्य और आंतरिक (S 1, S 2, S 3)। द्वितीयक गोले की स्तरित संरचना उनकी ताकत में काफी वृद्धि करती है। द्वितीयक झिल्ली में सेल्युलोज माइक्रोफाइब्रिल्स प्राथमिक की तुलना में सघन रूप से जमा होते हैं। लिग्निन - साधारण घटकलकड़ी के द्वितीयक गोले।
संपर्क कोशिकाओं की झिल्लियों में छिद्र एक दूसरे के विपरीत स्थित होते हैं। दो विरोधी छिद्र और एक छिद्र झिल्ली छिद्रों की एक जोड़ी बनाते हैं। द्वितीयक झिल्लियों वाली कोशिकाओं में, दो मुख्य प्रकार के छिद्र होते हैं: सरल और सीमाबद्ध। बॉर्डर वाले छिद्रों में, द्वितीयक दीवार छिद्र गुहा के ऊपर लटकी होती है। सिंपल पोर्स में ऐसा नहीं होता है।
कोशिका भित्ति का विकास। जैसे-जैसे कोशिका बढ़ती है, कोशिका झिल्ली की मोटाई और क्षेत्रफल बढ़ता जाता है। शेल स्ट्रेचिंग एक जटिल प्रक्रिया है। यह प्रोटोप्लास्ट द्वारा नियंत्रित होता है और हार्मोन ऑक्सिन द्वारा नियंत्रित होता है।
सभी दिशाओं में समान रूप से बढ़ने वाली कोशिकाओं में, मायोफिब्रिल्स का जमाव यादृच्छिक होता है। ये मायोफिब्रिल एक अनियमित नेटवर्क बनाते हैं। ऐसी कोशिकाएं तने के मूल भाग, भंडारण के ऊतकों और इन विट्रो में कोशिका संवर्धन के दौरान पाई गईं। लम्बी कोशिकाओं में, पार्श्व आवरण के पेशीतंतुओं को बढ़ाव के अक्ष पर समकोण पर जमा किया जाता है।
मैट्रिक्स पदार्थ - पेक्टिन, हेमिकेलुलोज और ग्लाइकोप्रोटीन को डिक्टीयोसोम के पुटिकाओं में झिल्ली में स्थानांतरित किया जाता है। इसी समय, पेक्टिन बढ़ती कोशिकाओं की अधिक विशेषता है, जबकि हेमिकेलुलोज गैर-बढ़ती कोशिकाओं में प्रबल होते हैं।
सेल्युलोज माइक्रोफिब्रिल को कोशिका की सतह पर प्लाज्मा झिल्ली से जुड़े एक एंजाइम कॉम्प्लेक्स की मदद से संश्लेषित किया जाता है। सूक्ष्मतंतुओं का उन्मुखीकरण स्थित सूक्ष्मनलिकाओं द्वारा नियंत्रित होता है भीतरी सतहप्लाज्मा झिल्ली।
प्लाज्मोडेस्मा। ये साइटोप्लाज्म के पतले धागे हैं जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ते हैं। प्लास्मोडेस्माटा या तो कहीं भी कोशिका भित्ति से होकर गुजरता है, या प्राथमिक छिद्र क्षेत्रों में या छिद्रों के जोड़े के बीच झिल्लियों में केंद्रित होता है। एक इलेक्ट्रॉन माइक्रोस्कोप के तहत, प्लास्मोडेमाटा प्लाज्मा झिल्ली के साथ पंक्तिबद्ध संकीर्ण चैनल के रूप में दिखाई देते हैं। नहर की धुरी के साथ, एक छोटी बेलनाकार ट्यूब, डेस्मोटुबुल, एक कोशिका से दूसरी कोशिका तक फैली हुई है, जो दोनों आसन्न कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम के साथ संचार करती है। कोशिका विभाजन के दौरान कई प्लास्मोडेस्माटा बनते हैं, जब ट्यूबलर एंडोप्लाज्मिक रेटिकुलम विकासशील सेल प्लेट द्वारा लिया जाता है। प्लास्मोडेस्माटा अविभाजित कोशिकाओं की झिल्लियों में भी बन सकता है। ये संरचनाएं कोशिका से कोशिका में कुछ पदार्थों का कुशल स्थानांतरण प्रदान करती हैं।
कोशिका विभाजन। पर बहुकोशिकीय जीवकोशिका विभाजन उनके आकार में वृद्धि के साथ पूरे जीव के विकास का एक तरीका है। विभाजन के दौरान बनने वाली नई कोशिकाएं मूल कोशिका और एक दूसरे दोनों की संरचना और कार्य में समान होती हैं। यूकेरियोट्स में विभाजन प्रक्रिया को दो आंशिक रूप से अतिव्यापी चरणों में विभाजित किया जा सकता है: माइटोसिस और साइटोकाइनेसिस।
माइटोसिस एक नाभिक से दो बेटी नाभिक का निर्माण होता है, एक दूसरे के रूपात्मक और आनुवंशिक रूप से समतुल्य। साइटोकिनेसिस बेटी कोशिकाओं के गठन के साथ कोशिका के साइटोप्लाज्मिक भाग का विभाजन है।
कोशिका चक्र। एक जीवित कोशिका क्रमिक घटनाओं की एक श्रृंखला से गुजरती है जो कोशिका चक्र बनाती है। चक्र की अवधि स्वयं कोशिका के प्रकार और के आधार पर भिन्न होती है बाह्य कारक, उदाहरण के लिए, तापमान या सुरक्षा पर पोषक तत्त्व. आमतौर पर चक्र को इंटरपेज़ और माइटोसिस के चार चरणों में विभाजित किया जाता है।
अंतरावस्था। क्रमिक माइटोटिक विभाजनों के बीच की अवधि।
इंटरपेज़ को तीन अवधियों में विभाजित किया गया है, जिसे G 1, S, G 2 के रूप में दर्शाया गया है।
जी 1 अवधि के दौरान, जो माइटोसिस के बाद शुरू होती है। इस अवधि के दौरान, विभिन्न जीवों सहित साइटोप्लाज्म की मात्रा बढ़ जाती है। इसके अलावा, आधुनिक परिकल्पना के अनुसार, जी 1 अवधि के दौरान, पदार्थ संश्लेषित होते हैं जो एस अवधि और शेष चक्र को उत्तेजित या बाधित करते हैं, इस प्रकार विभाजन प्रक्रिया का निर्धारण करते हैं।
एस अवधि जी 1 अवधि का अनुसरण करती है, जिस समय आनुवंशिक सामग्री (डीएनए) को दोहराया जाता है।
G2 अवधि के दौरान, जो S का अनुसरण करता है, माइटोसिस में सीधे शामिल संरचनाएं, जैसे कि स्पिंडल घटक, बनते हैं।
कुछ कोशिकाएं असीमित संख्या में सेल चक्रों से गुजरती हैं। यह एककोशिकीय जीवऔर सक्रिय विकास क्षेत्रों (मेरिस्टेम) की कुछ कोशिकाएँ। परिपक्वता के बाद कुछ विशिष्ट कोशिकाएं प्रजनन करने की अपनी क्षमता खो देती हैं। कोशिकाओं का तीसरा समूह, उदाहरण के लिए, घाव के ऊतक (कैलस) बनाने वाले, केवल विशेष परिस्थितियों में विभाजित करने की क्षमता को बरकरार रखते हैं।
मिटोसिस, या परमाणु विखंडन। यह एक सतत प्रक्रिया है जिसे चार चरणों में बांटा गया है: प्रोफ़ेज़, मेटाफ़ेज़, एनाफ़ेज़, टेलोफ़ेज़। माइटोसिस के परिणामस्वरूप, आनुवंशिक सामग्री जो इंटरपेज़ में दोगुनी हो गई है, दो बेटी नाभिकों के बीच समान रूप से विभाजित हो जाती है।
सबसे ज्यादा शुरुआती संकेतविभाजन के लिए एक कोशिका का संक्रमण सीधे प्लाज्मा झिल्ली के नीचे सूक्ष्मनलिकाएं की एक संकीर्ण, अंगूठी के आकार की बेल्ट की उपस्थिति है। यह अपेक्षाकृत सघन बेल्ट भविष्य के माइटोटिक स्पिंडल के विषुवतीय तल में नाभिक को घेरता है। चूंकि यह प्रोफ़ेज़ से पहले प्रकट होता है, इसलिए इसे प्रीप्रोफ़ेज़ गर्डल कहा जाता है। यह माइटोटिक स्पिंडल के बाद गायब हो जाता है, सेल प्लेट के लेट टेलोफ़ेज़ में प्रकट होने से बहुत पहले, जो केंद्र से परिधि तक बढ़ता है और प्रीप्रोफ़ेज़ बेल्ट द्वारा कब्जा किए गए क्षेत्र में मातृ कोशिका झिल्ली के साथ विलीन हो जाता है।
प्रोफ़ेज़। प्रोफ़ेज़ की शुरुआत में, गुणसूत्र नाभिक के अंदर बिखरे हुए लंबे धागों के समान होते हैं। फिर, जैसे-जैसे तंतु छोटे और मोटे होते जाते हैं, कोई यह देख सकता है कि प्रत्येक गुणसूत्र एक नहीं, बल्कि दो गुंथे हुए तंतुओं से बना होता है जिन्हें क्रोमैटिड कहा जाता है। देर से प्रोफ़ेज़ में, प्रत्येक गुणसूत्र के दो छोटे युग्मित क्रोमैटिड समानांतर में एक साथ होते हैं, जो सेंट्रोमियर नामक एक संकीर्ण खंड से जुड़े होते हैं। यह प्रत्येक गुणसूत्र पर एक विशिष्ट स्थिति रखता है और गुणसूत्र को अलग-अलग लंबाई की दो भुजाओं में विभाजित करता है।
सूक्ष्मनलिकाएं धुरी की धुरी के साथ नाभिक की सतह के समानांतर स्थित होती हैं। यह माइटोटिक स्पिंडल असेंबली की सबसे पहली अभिव्यक्ति है।
प्रोफ़ेज़ के अंत तक, नाभिक धीरे-धीरे अपनी स्पष्ट रूपरेखा खो देता है और अंत में गायब हो जाता है। इसके तुरंत बाद, परमाणु लिफाफा भी बिखर जाता है।
रूपक। मेटाफ़ेज़ की शुरुआत में, स्पिंडल, जो एक त्रि-आयामी संरचना है, मध्य में सबसे चौड़ा और ध्रुवों की ओर पतला होता है, पहले नाभिक द्वारा कब्जा कर लिया गया स्थान लेता है। स्पिंडल फाइबर सूक्ष्मनलिकाएं के बंडल हैं। मेटाफ़ेज़ के दौरान, क्रोमोसोम, जिनमें से प्रत्येक में दो क्रोमैटिड होते हैं, को व्यवस्थित किया जाता है ताकि उनके सेंट्रोमर्स स्पिंडल के इक्वेटोरियल प्लेन में स्थित हों। इसके सेंट्रोमियर के साथ, प्रत्येक गुणसूत्र स्पिंडल थ्रेड्स से जुड़ा होता है। हालाँकि, कुछ किस्में गुणसूत्रों से जुड़े बिना एक ध्रुव से दूसरे ध्रुव तक जाती हैं।
जब सभी गुणसूत्र विषुवतीय तल में स्थित होते हैं, तो मेटाफ़ेज़ पूरा हो जाता है। गुणसूत्र विभाजित होने के लिए तैयार होते हैं।
पश्चावस्था। प्रत्येक गुणसूत्र के क्रोमैटिड अलग हो जाते हैं। अब ये पुत्री गुणसूत्र हैं। सबसे पहले, सेंट्रोमियर विभाजित होता है, और दो बेटी गुणसूत्र विपरीत ध्रुवों पर खींचे जाते हैं। इस मामले में, सेंट्रोमर्स आगे बढ़ते हैं, और गुणसूत्रों की भुजाएँ पीछे की ओर खिंचती हैं। क्रोमोसोम से जुड़े स्पिंडल फिलामेंट्स छोटे हो जाते हैं, क्रोमैटिड्स को अलग करने और विपरीत दिशाओं में बेटी क्रोमोसोम के आंदोलन में योगदान करते हैं।
टेलोफ़ेज़। टेलोफ़ेज़ में, गुणसूत्रों के दो समान समूहों का पृथक्करण पूरा हो जाता है, जबकि उनमें से प्रत्येक के चारों ओर एक परमाणु झिल्ली बन जाती है। रफ रेटिकुलम इसमें सक्रिय भाग लेता है। धुरी तंत्र गायब हो जाता है। टेलोफ़ेज़ के दौरान, गुणसूत्र अपनी रूपरेखा की स्पष्टता खो देते हैं, फैल जाते हैं, पतले धागों में बदल जाते हैं। न्यूक्लियोली को बहाल किया जाता है। जब गुणसूत्र अदृश्य हो जाते हैं, तो माइटोसिस पूरा हो जाता है। दो संतति केंद्रक अंतरावस्था में प्रवेश करते हैं। वे आनुवंशिक रूप से एक दूसरे के और मूल नाभिक के बराबर हैं। यह बहुत महत्वपूर्ण है, क्योंकि अनुवांशिक कार्यक्रम, और इसके साथ सभी संकेतों को बेटी जीवों में स्थानांतरित किया जाना चाहिए।
माइटोसिस की अवधि भिन्न होती है विभिन्न जीवऔर यह कपड़े के प्रकार पर निर्भर करता है। हालाँकि, प्रोफ़ेज़ सबसे लंबा है और एनाफ़ेज़ सबसे छोटा है। रूट टिप की कोशिकाओं में, प्रोफ़ेज़ की अवधि 1-2 घंटे होती है; रूपक - 5 - 15 मिनट; पश्चावस्था - 2 - 10 मिनट; टेलोफ़ेज़ - 10 - 30 मिनट। इंटरपेज़ की अवधि 12 से 30 घंटे तक है।
कई यूकेरियोटिक कोशिकाओं में, माइटोटिक स्पिंडल के गठन के लिए जिम्मेदार सूक्ष्मनलिका आयोजन केंद्र सेंट्रीओल्स से जुड़े होते हैं।
साइटोकिनेसिस। यह साइटोप्लाज्म के विभाजन की प्रक्रिया है। अधिकांश जीवों में, कोशिकाएँ कोशिका भित्ति को पीछे खींचकर और एक विखंडन खांचे का निर्माण करके विभाजित होती हैं, जो धीरे-धीरे गहरा होता है, माइटोटिक धुरी के शेष तंतुओं को संकुचित करता है। सभी पौधों (ब्रायोफाइट्स और संवहनी पौधों) में और कुछ शैवाल में, कोशिका प्लेट के निर्माण के कारण कोशिकाएं विभाजित होती हैं।
शुरुआती टेलोफेज में, दो बेटी नाभिकों के बीच एक बैरल के आकार का तंतुओं की प्रणाली जिसे फेटामोप्लास्ट कहा जाता है। फैगमोप्लास्ट फाइबर, माइटोटिक स्पिंडल के तंतुओं की तरह, सूक्ष्मनलिकाएं होते हैं। फ्रैगमोप्लास्ट के विषुवतीय तल में छोटी-छोटी बूंदें दिखाई देती हैं। वे एक सेल प्लेट बनाने के लिए फ्यूज हो जाते हैं जो तब तक बढ़ता है जब तक कि वह विभाजित सेल की दीवार तक नहीं पहुंच जाता। यह दो बेटी कोशिकाओं के अलगाव को पूरा करता है। विलय की बूंदें गोल्गी तंत्र से निकलने वाले बुलबुले हैं। मूल रूप से, उनमें पेक्टिन पदार्थ होते हैं, जिनसे मध्य प्लेट बनती है। प्लेट के दोनों किनारों पर प्लाज्मा झिल्ली के निर्माण में पुटिका झिल्ली शामिल होती है। इसी समय, ट्यूबलर एंडोप्लाज्मिक रेटिकुलम के टुकड़ों से प्लास्मोडेस्मेटा बनता है।
मीडियन प्लेट के बनने के बाद, प्रत्येक प्रोटोप्लास्ट उस पर एक प्राथमिक झिल्ली जमा करता है। इसके अलावा, प्रत्येक बेटी कोशिका पूरे प्रोटोप्लास्ट के चारों ओर झिल्ली की एक नई परत जमा करती है, जो कोशिका प्लेट से उत्पन्न हुई झिल्ली को जारी रखती है। बेटी कोशिकाओं के बढ़ने पर मूल कोशिका का मूल खोल नष्ट हो जाता है।
विभिन्न प्रकार के यूकेरियोटिक माइटोसिस
पौधों और जानवरों में ऊपर वर्णित कोशिका विभाजन अप्रत्यक्ष कोशिका विभाजन का एकमात्र रूप नहीं है। माइटोसिस का सबसे सरल प्रकार प्लुरोमिटोसिस है। यह बाइनरी डिवीजन जैसा दिखता है प्रोकैरियोटिक कोशिकाएं, जिसमें प्रतिकृति के बाद न्यूक्लियॉइड प्लाज्मा झिल्ली से जुड़े रहते हैं। झिल्ली डीएनए बाध्यकारी बिंदुओं के बीच बढ़ने लगती है और इस प्रकार गुणसूत्रों को फैलती है अलग - अलग क्षेत्रकोशिकाओं। उसके बाद, एक कोशिका संकुचन के निर्माण के दौरान, प्रत्येक डीएनए अणु एक नई अलग कोशिका में होगा।
यूकेरियोटिक कोशिकाओं के विभाजन के लिए विशेषता सूक्ष्मनलिकाएं से निर्मित एक धुरी का गठन है। बंद pleuromitosis के साथ (इसे बंद कहा जाता है क्योंकि गुणसूत्रों का विचलन परमाणु लिफाफे को तोड़े बिना होता है), सेंट्रीओल्स नहीं, बल्कि अन्य संरचनाएं स्थित होती हैं अंदरआणविक झिल्ली। ये अनिश्चित आकृति विज्ञान के तथाकथित ध्रुवीय निकाय हैं, जिनसे सूक्ष्मनलिकाएं फैलती हैं। इनमें से दो शरीर हैं। वे बिना संपर्क खोए एक-दूसरे से अलग हो जाते हैं परमाणु लिफाफा. इसके परिणामस्वरूप, गुणसूत्रों से जुड़े दो अर्ध-स्पिंडल बनते हैं। इस मामले में माइटोटिक स्पिंडल गठन और गुणसूत्र अलगाव की पूरी प्रक्रिया परमाणु लिफाफे के अंतर्गत होती है। इस प्रकार का माइटोसिस प्रोटोजोआ के बीच होता है और व्यापक रूप से कवक (काइट्रिडिया, ज़ाइगोमाइसेट्स, यीस्ट, ओमीसाइकेट्स, एस्कोमाइसेट्स, मायक्सोमाइसेट्स, आदि) में वितरित किया जाता है। सेमी-क्लोज्ड प्लूरोमिटोसिस के रूप हैं, जब गठित स्पिंडल के ध्रुवों पर परमाणु लिफाफा नष्ट हो जाता है।
माइटोसिस का अगला रूप ऑर्थोमाइटोसिस है। इस मामले में, COMTs साइटोप्लाज्म में स्थित होते हैं, शुरुआत से ही, अर्ध-स्पिंडल नहीं बनते हैं, लेकिन एक द्विध्रुवीय स्पिंडल होता है। ऑर्थोमाइटोसिस (सामान्य माइटोसिस) के तीन रूप हैं, अर्ध-बंद और बंद। अर्ध-बंद ऑर्थोमाइटोसिस में, साइटोप्लाज्म में स्थित TsOMT की मदद से एक बिसिमेट्रिक स्पिंडल बनता है, ध्रुवीय क्षेत्रों के अपवाद के साथ, परमाणु लिफाफा पूरे माइटोसिस में संरक्षित होता है। दानेदार सामग्री, या सेंट्रीओल्स के द्रव्यमान को COMT के रूप में पहचाना जा सकता है। माइटोसिस का यह रूप कुछ में हरे, भूरे, लाल शैवाल के जूस्पोर्स में पाया जाता है कम मशरूमऔर ग्रेगरीन। बंद ऑर्थोमाइटोसिस के साथ, परमाणु झिल्ली पूरी तरह से संरक्षित होती है, जिसमें एक वास्तविक धुरी बनती है। सूक्ष्मनलिकाएं कार्योप्लाज्म में बनती हैं, कम बार वे आंतरिक TsOMT से बढ़ती हैं, जो परमाणु झिल्ली के साथ संबद्ध (प्लुरोमिटोसिस के विपरीत) नहीं होती है। इस प्रकार का माइटोसिस सिलियेट माइक्रोन्यूक्लि के विभाजन के लिए विशिष्ट है, लेकिन प्रोटोजोआ में भी हो सकता है। खुले ऑर्थोमाइटोसिस में, परमाणु लिफाफा पूरी तरह से विघटित हो जाता है। इस प्रकार का कोशिका विभाजन पशु जीवों, कुछ प्रोटोजोआ और उच्च पौधों की कोशिकाओं की विशेषता है। माइटोसिस का यह रूप, बदले में, सूक्ष्म और अनास्ट्रल प्रकारों द्वारा दर्शाया जाता है।
संक्षिप्त समीक्षा से, यह स्पष्ट है कि मुख्य विशेषतामिटोसिस सामान्य रूप से विखंडन धुरी की संरचनाओं का उद्भव है, जो TsOMT के संबंध में बनता है, जो संरचना में विविध है।
माइटोटिक आकृति की आकृति विज्ञान
माइटोसिस के मेटाफ़ेज़ चरण में माइटोटिक उपकरण विशेष रूप से अच्छी तरह से व्यक्त किया गया है। मेटाफ़ेज़ में, कोशिका के विषुवतीय तल में, गुणसूत्र स्थित होते हैं, जिससे तथाकथित स्पिंडल धागे विपरीत दिशाओं में फैलते हैं, माइटोटिक आकृति के दो अलग-अलग ध्रुवों पर परिवर्तित होते हैं। इस प्रकार, माइटोटिक स्पिंडल गुणसूत्रों, ध्रुवों और तंतुओं का एक संग्रह है। धुरी के तंतु एकल सूक्ष्मनलिकाएं या उनके बंडल होते हैं। सूक्ष्मनलिकाएं धुरी के ध्रुवों से शुरू होती हैं, और उनमें से कुछ सेंट्रोमर्स तक जाती हैं, जहां गुणसूत्र कीनेटोकोर (कीनेटोचोर माइक्रोट्यूबुल्स) स्थित होते हैं, कुछ विपरीत ध्रुव की ओर जाते हैं, लेकिन उस तक नहीं पहुंचते। उन्हें "इंटरपोलर माइक्रोट्यूबुल्स" कहा जाता है। रेडियल सूक्ष्मनलिकाएं का एक समूह ध्रुवों से निकलता है, जिससे उनके चारों ओर एक "उज्ज्वल चमक" जैसी संरचना बनती है - ये सूक्ष्म सूक्ष्मनलिकाएं हैं।
आकारिकी द्वारा, माइटोटिक आकृतियों को सूक्ष्म और अनास्ट्रल प्रकारों में विभाजित किया जाता है।
सूक्ष्म धुरी प्रकार, या अभिसारी, इस तथ्य की विशेषता है कि इसके ध्रुवों को एक छोटे से क्षेत्र द्वारा दर्शाया जाता है जिसमें सूक्ष्मनलिकाएं अभिसरण (अभिसरण) करती हैं। आमतौर पर, सेंट्रीओल्स वाले सेंट्रोसोम सूक्ष्म स्पिंडल के ध्रुवों पर स्थित होते हैं। हालांकि, सेंट्रीओलर एस्ट्रल माइटोस के मामले ज्ञात हैं (कुछ अकशेरूकीय के अर्धसूत्रीविभाजन के दौरान)। इसके अलावा, ध्रुवों से निकलने वाली रेडियल सूक्ष्मनलिकाएं नोट की जाती हैं, जो धुरी का हिस्सा नहीं हैं, लेकिन तारकीय क्षेत्र - सिटास्टर बनाती हैं। इस प्रकार का माइटोटिक डिवीजन एक डंबल जैसा दिखता है। माइटोटिक आकृति के एनास्ट्रियल प्रकार में ध्रुवों पर सिटास्टर नहीं होते हैं। यहां धुरी के ध्रुवीय क्षेत्र चौड़े हैं, उन्हें ध्रुवीय टोपी कहा जाता है, उनमें सेंट्रीओल्स शामिल हैं। इस मामले में, धुरी के तंतु एक बिंदु से नहीं हटते हैं, लेकिन ध्रुवीय टोपी के पूरे क्षेत्र से एक विस्तृत मोर्चे (विचलन) में विचलन करते हैं। इस प्रकार की धुरी उच्च पौधों की कोशिकाओं को विभाजित करने की विशेषता है, लेकिन उच्च जानवरों में भी पाई जा सकती है। स्तनधारियों के प्रारंभिक भ्रूणजनन में, सेंट्रीओलर (अपसारी) माइटोस डिम्बाणुजनकोशिका परिपक्वता के विभाजन के दौरान और जाइगोट के विभाजन I और II में देखे जाते हैं। लेकिन पहले से ही तीसरे कोशिका विभाजन में और बाद की सभी कोशिकाओं में, वे सूक्ष्म स्पिंडल की भागीदारी के साथ विभाजित होते हैं, जिनके ध्रुवों में सेंट्रीओल्स हमेशा पाए जाते हैं। सामान्य तौर पर, माइटोसिस के सभी रूपों के लिए, उनके किनेटोफ़ोर्स, ध्रुवीय निकायों (सेंट्रोसोम्स) और स्पिंडल फाइबर के साथ गुणसूत्र सामान्य संरचनाएं बने रहते हैं।
सेंट्रोमियर और काइनेटोकोर
सेंट्रोमर्स हो सकते हैं अलग स्थानीयकरणगुणसूत्रों की लंबाई के साथ। Holocentric centromeres तब होते हैं जब सूक्ष्मनलिकाएं पूरे गुणसूत्र (कुछ कीड़े, नेमाटोड, कुछ पौधे) की लंबाई के साथ जुड़ी होती हैं। मोनोसेंट्रिक सेंट्रोमियर - जब सूक्ष्मनलिकाएं एक साइट में गुणसूत्रों से जुड़ी होती हैं। मोनोसेन्ट्रिक सेंट्रोमर्स पंचर हो सकते हैं (उदाहरण के लिए, कुछ नवोदित यीस्ट में), जब केवल एक सूक्ष्मनलिका कीनेटोचोर तक पहुंचती है, और ज़ोनल, जहां सूक्ष्मनलिकाएं का एक बंडल जटिल किनेटोकोर तक पहुंचता है। सेंट्रोमियर जोन की विविधता के बावजूद, वे सभी जुड़े हुए हैं जटिल संरचनाकाइनेटोकोर, जिसमें सभी यूकेरियोट्स में संरचना और कार्य में मौलिक समानता है। काइनेटोकोर विशेष प्रोटीन संरचनाएं हैं अधिकाँश समय के लिएगुणसूत्रों के सेंट्रोमीटर में स्थित है। ये कई प्रोटीनों से युक्त जटिल परिसर हैं। रूपात्मक रूप से, वे बहुत समान हैं, समान संरचना है, डायटम से लेकर मनुष्यों तक। वे तीन-परत संरचनाएं हैं: गुणसूत्र के शरीर से सटे आंतरिक घने परत, मध्य ढीली परत और बाहरी घने परत। कई तंतु बाहरी परत से फैलते हैं, जो कीनेटोचोर के तथाकथित रेशेदार मुकुट का निर्माण करते हैं। में सामान्य फ़ॉर्मकिनेटोकोर्स में क्रोमोसोम के प्राथमिक कसना के क्षेत्र में, सेंट्रोमियर में प्लेट या डिस्क का रूप होता है। आमतौर पर प्रति क्रोमोसोम या क्रोमैटिड में एक काइनेटोकोर होता है। एनाफेज से पहले, प्रत्येक बहन क्रोमैटिड पर काइनेटोकोर्स स्थित होते हैं, प्रत्येक सूक्ष्मनलिकाएं के अपने स्वयं के बंडल के साथ संचार करता है। पौधों में, काइनेटोकोर प्लेटों की तरह नहीं, बल्कि गोलार्द्धों की तरह दिखता है। कीनेटोकोर्स की कार्यात्मक भूमिका बहन क्रोमैटिड्स को एक दूसरे से बांधना है, माइटोटिक माइक्रोट्यूबुल्स को ठीक करना, क्रोमोसोम पृथक्करण को विनियमित करना और वास्तव में माइटोसिस के दौरान माइक्रोट्यूबुल्स की भागीदारी के साथ क्रोमोसोम को स्थानांतरित करना है। सामान्य तौर पर, प्रोटीन संरचनाएं, किनेटोकोर्स, एस-अवधि में दोगुनी, गुणसूत्र दोहरीकरण के समानांतर। लेकिन उनके प्रोटीन कोशिका चक्र की सभी अवधियों में गुणसूत्रों पर मौजूद होते हैं।
संयंत्र ऊतक
शैक्षिक कपड़े(मेरिस्टम्स)
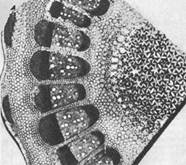
पौधों के शरीर में शैक्षिक ऊतक स्थित होते हैं अलग - अलग जगहें, इसलिए उन्हें निम्नलिखित समूहों में विभाजित किया गया है (चित्र 0; 1)।
1. एपिकल (एपिकल) मेरिस्टेम्स अक्षीय अंगों - तने, जड़ के शीर्ष या शीर्ष पर स्थित होते हैं। इन विभज्योतकों की सहायता से पौधों के कायिक अंग लंबाई में वृद्धि करते हैं।
2. पार्श्व विभज्योतक अक्षीय अंगों की विशेषता है। वहां वे क्लच के रूप में केंद्रित रूप से स्थित हैं।
3. इंटरक्लेरी, या इंटरक्लेरी, मेरिस्टेम एपिकल मेरिस्टेम से उत्पन्न होते हैं। ये कोशिकाओं के समूह हैं जो अभी तक गुणा करने में सक्षम नहीं हैं, लेकिन भेदभाव के मार्ग पर चल पड़े हैं। उनमें कोई प्रारंभिक कोशिकाएँ नहीं हैं, लेकिन कई विशिष्ट कोशिकाएँ हैं।
4. घाव मेरिस्टेम शरीर के क्षतिग्रस्त हिस्से की बहाली प्रदान करते हैं। पुनर्जनन की शुरुआत निर्विभेदीकरण से होती है, यानी विशिष्ट कोशिकाओं से विभज्योतक कोशिकाओं तक का उल्टा विकास। वे फेलोजेन में बदल जाते हैं, जो एक प्लग बनाता है जो घाव की सतह को ढकता है। अलग-अलग विभाजित कोशिकाएं, विभाजित, एक ढीले पैरेन्काइमल ऊतक - कैलस बना सकती हैं। इससे कुछ शर्तों के तहत पौधे के अंगों का निर्माण होता है।
पूर्णांक ऊतक
वे एक सीमा बाधा के रूप में कार्य करते हैं, अंतर्निहित ऊतकों को अलग करते हैं पर्यावरण. पौधे के प्राथमिक अध्यावरण में केवल जीवित कोशिकाएँ होती हैं। द्वितीयक और तृतीयक अध्यावरण - अधिकतर मृत कोशिका भित्तियों से प्राप्त होता है।
पूर्णांक ऊतकों के मुख्य कार्य:
पौधे को सूखने से बचाना;
·प्रवेश संरक्षण हानिकारक सूक्ष्मजीव;
· से रक्षा धूप की कालिमा;
· से रक्षा यांत्रिक क्षति;
संयंत्र और पर्यावरण के बीच चयापचय का विनियमन;
जलन की धारणा।
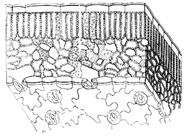
प्राथमिक पूर्णांक ऊतक एपिडर्मिस, एपिडर्मिस है। जीवित कोशिकाओं से मिलकर बनता है। शीर्ष विभज्योतक से निर्मित। युवा बढ़ते तनों और पत्तियों को ढकता है।
एपिडर्मिस का गठन पौधों में जलीय आवास से बाहर निकलने के संबंध में किया गया था ताकि इसे सूखने से रोका जा सके। स्टोमेटा के अलावा, एपिडर्मिस की सभी कोशिकाएं कसकर आपस में जुड़ी होती हैं। मुख्य कोशिकाओं की बाहरी दीवारें बाकी की तुलना में मोटी होती हैं। पूरी सतह क्यूटिन और वेजिटेबल वैक्स की एक परत से ढकी होती है। इस परत को क्यूटिकल (त्वचा) कहते हैं। यह पौधों की बढ़ती जड़ों और पानी के नीचे के हिस्सों पर अनुपस्थित है। सूखने पर छल्ली की पारगम्यता काफी कमजोर हो जाती है।
मुख्य कोशिकाओं के अलावा, एपिडर्मिस में अन्य भी होते हैं, विशेष रूप से बाल या ट्राइकोम्स में। वे एककोशिकीय और बहुकोशिकीय हैं (चित्र 2)। कार्यात्मक रूप से, वे एपिडर्मिस की सतह को बढ़ाते हैं, उदाहरण के लिए, जड़ विकास क्षेत्र में, यांत्रिक सुरक्षा के रूप में कार्य करते हैं, एक समर्थन से चिपकते हैं, और पानी के नुकसान को कम करते हैं। कई पौधों में ग्रंथियों के बाल होते हैं, जैसे बिछुआ।
केवल उच्च पौधों में एपिडर्मिस में रंध्र होते हैं, जो पानी और गैसों के आदान-प्रदान को नियंत्रित करते हैं। यदि छल्ली नहीं है, तो रंध्रों की कोई आवश्यकता नहीं है। स्टोमेटा कोशिकाओं का एक समूह है जो रंध्र तंत्र का निर्माण करता है, जिसमें दो अनुगामी कोशिकाएँ और आस-पास के एपिडर्मल कोशिकाएँ - पार्श्व कोशिकाएँ होती हैं। वे मुख्य एपिडर्मल कोशिकाओं (चित्र 3) से भिन्न होते हैं। गार्ड कोशिकाएं अपने आसपास की कोशिकाओं से आकार और बड़ी संख्या में क्लोरोप्लास्ट और असमान रूप से मोटी दीवारों की उपस्थिति से भिन्न होती हैं। जो एक दूसरे का सामना करते हैं वे बाकी की तुलना में मोटे होते हैं (चित्र 4)। गार्ड कोशिकाओं के बीच, एक रंध्र अंतर बनता है, जो एक सबस्टोमेटल स्पेस की ओर जाता है जिसे सबस्टोमेटल कैविटी कहा जाता है। गार्ड कोशिकाएं अत्यधिक प्रकाश संश्लेषक होती हैं। वे होते हैं एक बड़ी संख्या कीअतिरिक्त स्टार्च और कई माइटोकॉन्ड्रिया।
रंध्रों की संख्या और वितरण, रंध्र तंत्र के प्रकार विभिन्न पौधों में व्यापक रूप से भिन्न होते हैं। आधुनिक ब्रायोफाइट्स में रंध्र अनुपस्थित होते हैं। उनमें प्रकाश संश्लेषण गैमेटोफाइट पीढ़ी द्वारा किया जाता है, और स्पोरोफाइट स्वतंत्र अस्तित्व में सक्षम नहीं होते हैं।
आमतौर पर रंध्र पत्ती के नीचे की तरफ स्थित होते हैं। पानी की सतह पर तैरने वाले पौधों में - ऊपरी सतह पर। अनाज की पत्तियों में, रंध्र अक्सर दोनों तरफ समान रूप से स्थित होते हैं। ऐसी पत्तियाँ अपेक्षाकृत समान रूप से प्रकाशित होती हैं। सतह के 1 मिमी 2 पर, 100 से 700 रंध्र स्थित हो सकते हैं।
द्वितीयक पूर्णांक ऊतक (पेरिडर्म)। यह ऊतक कब एपिडर्मिस को बदल देता है हरा रंगवार्षिक शूटिंग को भूरे रंग से बदल दिया जाता है। यह बहुस्तरीय है और कैम्बियल कोशिकाओं की एक केंद्रीय परत होती है - फेलोजेन। फेलोजेन कोशिकाएं, विभाजन, बाहर की ओर फेलेम की एक परत और अंदर की ओर फेलोडर्म (चित्र 5) बिछाती हैं।
फेलिमा, या कॉर्क। सबसे पहले इसमें जीवित पतली दीवार वाली कोशिकाएँ होती हैं। समय के साथ, उनकी दीवारें साबरिन और वनस्पति मोम से संतृप्त हो जाती हैं और मर जाती हैं। सेल की सामग्री हवा से भरी हुई है।
फेलिमा कार्य:
नमी की कमी को रोकता है
पौधे को यांत्रिक क्षति से बचाता है;
रोगजनकों से बचाता है
थर्मल इन्सुलेशन प्रदान करता है, क्योंकि कोशिकाएं हवा से भरी होती हैं।
एपिडर्मिस में स्थित फेलोजेन की कोशिकाएं, अंतर्निहित सबपीडर्मल परत, प्राथमिक प्रांतस्था की गहरी परतों में अक्सर कम होती हैं, प्राथमिक प्रांतस्था का निर्माण आधार होती हैं।
कॉर्क परत स्थायी नहीं है। इसमें ब्रेक होते हैं, जो पास में स्थित इंटरसेलुलर स्पेस के साथ संचार करते हैं। इसी समय, सतह पर छोटे ट्यूबरकल बनते हैं - लेंटिकल्स, जो वायुमंडलीय वायु (चित्र। 6.7) के साथ अंतरकोशिकीय स्थानों के रिक्त स्थान का संचार करते हैं।
शरद ऋतु में, मसूर के नीचे फेलोजेन कॉर्क कोशिकाओं की एक परत देता है, जो वाष्पोत्सर्जन को बहुत कम करता है, लेकिन इसे पूरी तरह से बाहर नहीं करता है। वसंत ऋतु में यह परत अंदर से ढह जाती है। हल्की बर्च की छाल पर, काले धब्बे के रूप में दाल स्पष्ट रूप से दिखाई देती है।
तृतीयक पूर्णांक ऊतक (क्रस्ट) भी केवल पौधों के लकड़ी के रूपों की विशेषता है।
कॉर्टेक्स की गहरी परतों में फेलोजेन को बार-बार रखा जाता है। इसके बाहर के ऊतक अंततः मर जाते हैं, एक पपड़ी बनाते हैं। इसकी कोशिकाएँ मर चुकी होती हैं और फैलने में असमर्थ होती हैं। हालांकि, जीवित कोशिकाएं गहरे विभाजन में स्थित होती हैं, जिससे तने के अनुप्रस्थ आकार में वृद्धि होती है। समय के साथ, परत की बाहरी परत टूट जाती है। इस तरह के अंतराल की घटना का समय व्यक्तिगत पौधों के लिए काफी स्थिर मूल्य है। सेब के पेड़ में, यह जीवन के सातवें वर्ष में होता है, हॉर्नबीम में - पचासवें में। कुछ प्रजातियों में यह बिल्कुल नहीं होता है। छीलने का मुख्य कार्य यांत्रिक और थर्मल क्षति से सुरक्षा है।
पैरेन्काइमा
यह विशिष्ट ऊतकों का एक समूह है जो प्रवाहकीय और यांत्रिक ऊतकों (चित्र 8) के बीच संयंत्र शरीर के अंदर रिक्त स्थान को भरता है। अधिक बार पैरेन्काइमा कोशिकाओं का एक गोल आकार होता है, कम अक्सर एक लम्बी आकृति। विकसित अंतरकोशिकीय स्थानों की उपस्थिति विशेषता है। कोशिकाओं के बीच के रिक्त स्थान एक साथ एक परिवहन प्रणाली - एपोप्लास्ट बनाते हैं। इसके अलावा, इंटरसेलुलर रिक्त स्थान पौधे की "वेंटिलेशन सिस्टम" बनाते हैं। रंध्रों, या मसूर के माध्यम से, वे वायुमंडलीय हवा से जुड़े होते हैं और इष्टतम प्रदान करते हैं गैस रचनापौधे के अंदर। जलभराव वाली मिट्टी पर उगने वाले पौधों के लिए विकसित अंतरकोशिकीय स्थान विशेष रूप से आवश्यक होते हैं, जहां सामान्य गैस विनिमय मुश्किल होता है। इस पैरेन्काइमा को एरेन्काइमा (चित्र 9) कहा जाता है।
पैरेन्काइमा के तत्व, अन्य ऊतकों के बीच अंतराल को भरते हुए, एक समर्थन का कार्य भी करते हैं। पैरेन्काइमा कोशिकाएं जीवित होती हैं, उनमें स्क्लेरेन्काइमा की तरह मोटी कोशिका भित्ति नहीं होती है। इसलिए, यांत्रिक गुण टर्गर द्वारा प्रदान किए जाते हैं। यदि पानी की मात्रा कम हो जाती है, जिससे पौधे का प्लास्मोलिसिस और विल्टिंग हो जाता है।
एसिमिलेशन पैरेन्काइमा पतली दीवार वाली कोशिकाओं द्वारा कई अंतरकोशिकीय स्थानों के साथ बनता है। इस संरचना की कोशिकाओं में बहुत सारे क्लोरोप्लास्ट होते हैं, इसलिए इसे क्लोरेन्काइमा कहा जाता है। क्लोरोप्लास्ट एक दूसरे को छायांकित किए बिना दीवार के साथ स्थित होते हैं। एसिमिलेशन पैरेन्काइमा में, प्रकाश संश्लेषण प्रतिक्रियाएं होती हैं, जो पौधे को कार्बनिक पदार्थ और ऊर्जा प्रदान करती हैं। प्रकाश संश्लेषक प्रक्रियाओं का परिणाम पृथ्वी पर सभी जीवित जीवों के अस्तित्व की संभावना है।
आत्मसात ऊतक केवल पौधे के प्रबुद्ध भागों में मौजूद होते हैं, वे एक पारदर्शी एपिडर्मिस द्वारा पर्यावरण से अलग होते हैं। यदि एपिडर्मिस को अपारदर्शी द्वितीयक पूर्णांक ऊतकों द्वारा प्रतिस्थापित किया जाता है, तो एसिमिलेशन पैरेन्काइमा गायब हो जाता है।
भंडारण पैरेन्काइमा कार्बनिक पदार्थों के लिए एक पात्र के रूप में कार्य करता है जो अस्थायी रूप से पौधे जीव द्वारा उपयोग नहीं किया जाता है। मूल रूप से स्थगित करना कार्बनिक पदार्थजैसा विभिन्न प्रकारसमावेशन जीवित प्रोटोप्लास्ट के साथ किसी भी कोशिका के लिए सक्षम हैं, लेकिन कुछ कोशिकाएं इसमें विशेषज्ञ हैं (चित्र 10, 11)। ऊर्जा से भरपूर यौगिकों को केवल बढ़ते मौसम के दौरान जमा किया जाता है, सुप्त अवधि के दौरान और अगले बढ़ते मौसम की तैयारी में खपत होती है। इसलिए, आरक्षित पदार्थ केवल बारहमासी पौधों में वनस्पति अंगों में जमा होते हैं।
शेयरों का भंडार सामान्य अंग (पलायन, जड़), साथ ही विशेष वाले (राइजोम, कंद, बल्ब) हो सकते हैं। सभी बीज पौधे बीजों (बीजपत्र, एंडोस्पर्म) में ऊर्जावान रूप से मूल्यवान पदार्थों को संग्रहित करते हैं। एक शुष्क जलवायु के कई पौधे न केवल कार्बनिक पदार्थ, बल्कि पानी भी जमा करते हैं (चित्र 12)। उदाहरण के लिए, मुसब्बर मांसल पत्तियों में पानी जमा करता है, शूटिंग में कैक्टि।
यांत्रिक कपड़े
पादप कोशिकाओं के यांत्रिक गुणों द्वारा प्रदान किया जाता है:
कोशिका का कठोर खोल
स्फीति, अर्थात् कोशिकाओं की स्फीति अवस्था।
इस तथ्य के बावजूद कि लगभग सभी ऊतक कोशिकाओं में यांत्रिक गुण होते हैं, हालांकि, पौधे में ऐसे ऊतक होते हैं जिनके लिए यांत्रिक गुण बुनियादी होते हैं। ये कॉलेन्काइमा और स्क्लेरेन्काइमा हैं। वे आमतौर पर अन्य ऊतकों के साथ बातचीत में कार्य करते हैं। शरीर के अंदर पौधे एक प्रकार का ढाँचा बनाते हैं। इसलिए, उन्हें सुदृढीकरण कहा जाता है।
सभी पौधे समान रूप से अच्छी तरह अभिव्यक्त नहीं होते हैं यांत्रिक कपड़े. बहुत कम हद तक, रहने वाले पौधे जलीय वातावरणस्थलीय की तुलना में। कारण यह है कि जलीय पौधोंकम आंतरिक समर्थन की जरूरत है। उनका शरीर काफी हद तक आसपास के पानी से समर्थित है। जमीन पर हवा इस तरह का समर्थन नहीं बनाती है, क्योंकि इसमें पानी की तुलना में कम घनत्व होता है। यही कारण है कि विशिष्ट यांत्रिक ऊतकों की उपस्थिति प्रासंगिक हो जाती है।
विकास की प्रक्रिया में आंतरिक समर्थन संरचनाओं में सुधार हुआ।
Collenchyma। यह केवल जीवित कोशिकाओं द्वारा बनता है, जो अंग की धुरी के साथ लम्बी होती हैं। प्राथमिक वृद्धि की अवधि के दौरान इस प्रकार के यांत्रिक ऊतक बहुत जल्दी बनते हैं। इसलिए, यह महत्वपूर्ण है कि कोशिकाएं जीवित रहें और पास की खींची हुई कोशिकाओं के साथ-साथ फैलने की क्षमता बनाए रखें।
कोलेन्काइमा कोशिकाओं की विशेषताएं:
खोल का असमान मोटा होना, जिसके परिणामस्वरूप इसके कुछ हिस्से पतले रहते हैं, जबकि अन्य मोटे हो जाते हैं;
गोले लिग्निफाई नहीं करते हैं।
Collenchyma कोशिकाएं एक दूसरे के सापेक्ष अलग-अलग स्थित होती हैं। आसन्न कोशिकाओं में, एक दूसरे के सामने वाले कोनों पर गाढ़ापन बनता है (चित्र 13)। ऐसे कोलेन्काइमा को कॉर्नर कॉलेन्काइमा कहा जाता है। दूसरे मामले में, कोशिकाओं को समानांतर परतों में व्यवस्थित किया जाता है। इन परतों का सामना करने वाली कोशिका झिल्लियाँ अत्यधिक मोटी होती हैं। यह लैमेलर कोलेंकाइमा है। प्रचुर मात्रा में अंतरकोशिकीय रिक्त स्थान के साथ कोशिकाएं शिथिल रूप से स्थित हो सकती हैं - यह एक ढीला कोलेन्काइमा है। इस तरह के कोलेंकाइमा अक्सर जलभराव वाली मिट्टी पर पौधों में पाए जाते हैं।
Collenchyma युवा पौधों, जड़ी-बूटियों के रूपों, और पौधों के उन हिस्सों में भी विशेष महत्व रखता है जहाँ द्वितीयक वृद्धि नहीं होती है, जैसे कि पत्तियाँ। इस मामले में, इसे सतह के बहुत करीब रखा जाता है, कभी-कभी सिर्फ एपिडर्मिस के नीचे। यदि अंग के चेहरे हैं, तो उनके शिखर के साथ कोलेन्काइमा की मोटी परतें पाई जाती हैं।
Collenchyma कोशिकाएं केवल स्फीति की उपस्थिति में क्रियाशील होती हैं। पानी की कमी से कॉलेंकाइमा की प्रभावशीलता कम हो जाती है और पौधा अस्थायी रूप से मुरझा जाता है, उदाहरण के लिए, ककड़ी के पत्ते गर्म दिन में गिर जाते हैं। कोशिकाओं को पानी से भरने के बाद, कोलेंकाइमा के कार्य बहाल हो जाते हैं।
दृढ़ोतक। दूसरे प्रकार के यांत्रिक ऊतक। Collenchyma के विपरीत, जहाँ सभी कोशिकाएँ जीवित हैं, स्क्लेरेन्काइमा कोशिकाएँ मृत हैं। इनकी दीवारें बहुत मोटी होती हैं। वे एक यांत्रिक कार्य करते हैं। झिल्ली का एक मजबूत मोटा होना पदार्थों के परिवहन में व्यवधान पैदा करता है, जिसके परिणामस्वरूप प्रोटोप्लास्ट मर जाता है (चित्र 14)। स्क्लेरेन्काइमा की कोशिका झिल्लियों का लिग्निफिकेशन तब होता है जब पौधे का अंग पहले ही अपनी वृद्धि पूरी कर चुका होता है। इसलिए, वे अब आसपास के ऊतकों के खिंचाव में हस्तक्षेप नहीं करते हैं।
आकार के आधार पर, दो प्रकार की स्क्लेरेन्काइमा कोशिकाएं प्रतिष्ठित होती हैं - तंतु और स्केलेरिड।
तंतुओं में बहुत मोटी दीवारों और एक छोटी गुहा के साथ एक मजबूत लम्बी आकृति होती है। वे लकड़ी के तंतुओं से कुछ छोटे होते हैं। अक्सर एपिडर्मिस के नीचे अनुदैर्ध्य परतें और किस्में बनती हैं। फ्लोएम या जाइलम में ये अकेले या समूहों में पाए जाते हैं। फ्लोएम में उन्हें बास्ट फाइबर (चित्र 15) कहा जाता है, और जाइलम में उन्हें लाइब्रिफॉर्म फाइबर (चित्र 16) कहा जाता है।
स्केलेरिड्स, या स्टोनी कोशिकाएं, शक्तिशाली झिल्लियों के साथ गोल या शाखित कोशिकाओं द्वारा दर्शाई जाती हैं। एक पौधे के शरीर में, वे अकेले (सहायक कोशिकाओं) (चित्र 17) या समूहों (चित्र 18) में पाए जा सकते हैं। यह ध्यान दिया जाना चाहिए कि यांत्रिक गुण दृढ़ता से स्केलेरिड्स के स्थान पर निर्भर करते हैं। स्क्लेराइड्स का हिस्सा निरंतर परतें बनाता है, उदाहरण के लिए, नट के खोल में या फलों के पत्थरों (पत्थर के फल) में।
उत्सर्जक ऊतक
कोई लिविंग सेलप्रारंभ में पदार्थों को हटाने का कार्य होता है, जबकि परिवहन एकाग्रता ढाल के साथ और ढाल के विरुद्ध दोनों जा सकता है। पदार्थ जो कोशिका में संश्लेषित किए गए हैं उन्हें हटाया जा सकता है और बाद में अन्य कोशिकाओं (फाइटोहोर्मोन, एंजाइम) की गतिविधि को प्रभावित करेगा। इस प्रक्रिया को स्राव कहते हैं। मामले में जब पदार्थ जो कोशिका के अपशिष्ट उत्पाद हैं, उत्सर्जित होते हैं, इस प्रक्रिया को उत्सर्जन कहा जाता है। इस तथ्य के बावजूद कि पौधों और जानवरों की कोशिकाओं में पदार्थों के उत्सर्जन में मौलिक समानता है, पौधों में कई विशेषताएं हैं जो महत्वपूर्ण गतिविधि में मौलिक अंतर से उत्पन्न होती हैं। पौधों में चयापचय प्रतिक्रियाओं का स्तर जानवरों की तुलना में बहुत कम है। इसलिए, आनुपातिक रूप से कम मात्रा में कचरा उत्सर्जित होता है। एक और विशेषता यह है कि स्वतंत्र रूप से लगभग सभी आवश्यक संश्लेषण करता है कार्बनिक यौगिक, पौधे कभी भी अत्यधिक भंडार नहीं बनाते हैं। जारी किए गए पदार्थ अन्य प्रतिक्रियाओं के लिए प्रारंभिक सामग्री के रूप में काम कर सकते हैं (उदाहरण के लिए, सीओ 2 और एच 2 ओ)।
यदि जानवरों में विषाक्त पदार्थों के उत्सर्जन की प्रक्रिया शरीर से उनके निष्कासन से जुड़ी होती है, तो पौधे ऐसा नहीं कर सकते हैं, मृत कोशिकाओं (अधिकांश ऊतकों) में जीवित प्रोटोप्लास्ट (वैक्यूल में विभिन्न पदार्थों को हटाने) में अनावश्यक पदार्थों को अलग करते हैं। बारहमासी पौधाऐसी कोशिकाओं से मिलकर बनता है), अंतरकोशिकीय स्थानों में।
मुक्त पदार्थों को दो भागों में विभाजित किया जा सकता है बड़े समूह. पहला समूह कार्बनिक पदार्थ है जो सीधे कोशिका द्वारा संश्लेषित होता है (एंजाइम, पॉलीसेकेराइड, लिग्निन, टेरपेन, बाद वाले घटक तत्व हैं) ईथर के तेलऔर राल)। दूसरा समूह - पदार्थ जो शुरू में जाइलम या फ्लोएम परिवहन (पानी, पानी) की मदद से बाहर से कोशिका में प्रवेश करते हैं। खनिज लवण, अमीनो एसिड, मोनोसुगर, आदि)। उत्सर्जित रहस्य शायद ही कभी सजातीय होता है और आमतौर पर एक मिश्रण होता है जिसमें एक पदार्थ में उच्चतम एकाग्रता होती है।
पौधों, जानवरों के विपरीत, पूर्ण की कमी है निकालनेवाली प्रणाली. पूरे संयंत्र में बिखरी हुई केवल विशेष संरचनाएं हैं - इडियोब्लास्ट्स। उन्हें बनाने वाली कोशिकाएँ पास में पड़ी पैरेन्काइमा कोशिकाओं से छोटी होती हैं। उनके पास एक इलेक्ट्रॉन-सघन साइटोप्लाज्म है, जिसमें एंडोप्लाज्मिक रेटिकुलम और गोल्गी कॉम्प्लेक्स के विकसित तत्व हैं। बहुधा व्यक्त नहीं किया जाता है केंद्रीय रिक्तिका. अपने और अन्य जीवित कोशिकाओं के बीच, ये कोशिकाएँ असंख्य प्लास्मोडेस्माटा द्वारा जुड़ी होती हैं।
स्थान के आधार पर, उत्सर्जक ऊतक बाहरी और आंतरिक हो सकते हैं।
बाहरी उत्सर्जक ऊतक। ग्रंथियों के बाल एपिडर्मिस के डेरिवेटिव हैं। Morphologically, वे परिवर्तनशील हैं - उनके पास एक बहुकोशिकीय सिर हो सकता है, लम्बी हो सकती है, एक पैर पर ढाल के रूप में, आदि। (अंजीर। 19)। इनमें चुभने वाले बिछुआ बाल शामिल हैं।
हाइडेथोड ऐसी संरचनाएं हैं जो कम वाष्पोत्सर्जन और उच्च आर्द्रता - गट्टेशन की स्थितियों में अतिरिक्त पानी छोड़ती हैं। ये संरचनाएं रंगहीन जीवित कोशिकाओं के समूहों द्वारा पतली दीवारों के साथ बनाई जाती हैं - एपिथेम। यह ऊतक प्रवाहकीय बंडल (चित्र 20) के निकट है। पानी को विशेष जल रंध्रों के माध्यम से छोड़ा जाता है, जो सामान्य रंध्रों से गतिहीनता और लगातार खुले अंतराल से भिन्न होते हैं। गुटेशन तरल पदार्थ की संरचना व्यापक रूप से लगभग भिन्न होती है शुद्ध पानीपदार्थों के एक बहुत ही जटिल मिश्रण के लिए।
अमृत। कई पौधों को 7 से 87% मोनो- और ओलिगोसेकेराइड युक्त तरल की रिहाई की विशेषता है। यह प्रक्रिया विशेष संरचनाओं - अमृत द्वारा की जाती है। स्थान के आधार पर, फूल, या पुष्प, अमृत प्रतिष्ठित हैं (चित्र 21), साथ ही तनों और पत्तियों पर स्थित पौधे - अतिरिक्त पुष्प, या अतिरिक्त अमृत। मकरंद अपरिवर्तित फ्लोएम रस हो सकता है, जो अंतरकोशिकीय स्थानों के माध्यम से सतह पर पहुंचाया जाता है और रंध्रों के माध्यम से उत्सर्जित होता है। ग्रंथियों के पैरेन्काइमा द्वारा अधिक जटिल अमृत बनते हैं, जो ग्रंथियों के बालों के साथ एपिडर्मिस से ढके होते हैं। अमृत या तो एपिडर्मल कोशिकाओं या ग्रंथियों के बालों द्वारा उत्सर्जित होता है। इस मामले में, स्रावित अमृत फ्लोएम सैप से भिन्न होता है। चूंकि इसमें ग्लूकोज, फ्रुक्टोज, सुक्रोज और फ्लोएम जूस - ग्लूकोज का प्रभुत्व है। आयन कम सांद्रता में मौजूद हैं। आयन कम सांद्रता में मौजूद हैं। परागणकों को आकर्षित करने के लिए अमृत में शामिल हो सकता है स्टेरॉयड हार्मोनजो कीड़ों के लिए आवश्यक हैं।
कीटभक्षी पौधों (सूंड्यू, नेपेंथिस, आदि) में पाचन ग्रंथियां मौजूद होती हैं। छोटे अकशेरूकीय के स्पर्श के जवाब में, वे हाइड्रोलाइटिक एंजाइम, फॉर्मिक, ब्यूटिरिक और प्रोपियोनिक एसिड युक्त एक रहस्य का स्राव करते हैं। नतीजतन, बायोपॉलिमर्स मोनोमर्स में टूट जाते हैं, जिनका उपयोग पौधे अपनी जरूरतों के लिए करते हैं।
लवणीय मिट्टी में उगने वाले पौधों में लवण ग्रंथियाँ विकसित होती हैं। ये ग्रंथियां पत्तियों पर स्थित होती हैं और घुले हुए रूप में, अतिरिक्त आयन सतह पर लाए जाते हैं। नमक पहले छल्ली पर जमा होता है, फिर इसे बारिश से धोया जाता है (चित्र 22)। इसके अलावा, कई पौधों की पत्तियों पर नमक के बाल होते हैं। प्रत्येक बाल में दो कोशिकाएँ होती हैं: एक सिर बनाती है, दूसरी - तना। सिर की कोशिका की रसधानी में लवण जमा हो जाते हैं, और जब उनकी सघनता अधिक हो जाती है, तो सिर गिर जाता है। इसके स्थान पर एक नई कोशिका विकसित होती है, जो लवण भी जमा करती है। पत्ती के विकास के दौरान, सिर गिर सकता है और कई बार बन सकता है। नमक बालों का लाभ यह है कि नमक छोड़ने से वे थोड़ा पानी खो देते हैं। खारी मिट्टी पर उगने वाले पौधों के लिए यह घटना बहुत महत्वपूर्ण है, जहां ताजे पानी की कमी है।
आंतरिक उत्सर्जक ऊतक। इडियोब्लास्ट्स के रूप में पौधे के पूरे शरीर में बिखरी हुई आंतरिक उत्सर्जन संरचनाएं, शरीर के बाहर पदार्थों को नहीं हटाती हैं, उन्हें स्वयं में जमा करती हैं। इडियोब्लास्ट पादप कोशिकाएं हैं जो एक ही ऊतक में अन्य कोशिकाओं से आकार, संरचना या सामग्री में भिन्न होती हैं। उत्पत्ति के आधार पर, स्रावित पदार्थों के स्किज़ोजेनिक (ग्रीक स्किसो - स्प्लिट) और लाइसोजेनिक (ग्रीक लिसीज़ - विघटन) रिसेप्टेकल्स को प्रतिष्ठित किया जाता है (चित्र 23)।
स्किज़ोजेनिक रिसेप्टेकल्स। ये कमोबेश व्यापक अंतरकोशिकीय स्थान हैं जो उत्सर्जित पदार्थों से भरे हुए हैं। उदाहरण के लिए, कोनिफर्स, अरालिया, छाता और अन्य के राल मार्ग। माना जाता है कि राल है जीवाणुनाशक गुण, शाकाहारियों को पीछे हटाता है, पौधों को उनके लिए अखाद्य बनाता है।
लाइसजेनिक रिसेप्टेकल्स जीवित कोशिकाओं के स्थान पर बनते हैं, जो मर जाते हैं और उनमें पदार्थों के जमा होने के बाद नष्ट हो जाते हैं। खट्टे फलों के छिलके में इस तरह की संरचनाएँ देखी जा सकती हैं।
मुख्य प्रकार के अलावा, मध्यवर्ती रूप भी हैं। दुहने वाले दो प्रकार के होते हैं: मुखर और गैर-मुखर। पूर्व का निर्माण जंजीरों में व्यवस्थित कई जीवित कोशिकाओं से होता है। संपर्क के बिंदु पर, गोले नष्ट हो जाते हैं, प्रोटोप्लास्ट विलीन हो जाते हैं, और परिणामस्वरूप एक एकल नेटवर्क बनता है। ऐसे दूधिया कम्पोजिट में पाए जाते हैं। गैर-खंडित लैक्टिक कोशिकाएं एक विशाल बहुकेन्द्रीय कोशिका द्वारा बनाई जाती हैं। यह विकास के प्रारंभिक चरण में उत्पन्न होता है, लंबा होता है और पौधे के बढ़ने पर शाखाएं, एक जटिल नेटवर्क बनाते हैं, उदाहरण के लिए, यूफोरबिया पौधों (चित्र 25) में।
लैक्टिफेरस की कोशिका भित्ति अत्यधिक जलयुक्त होती है। वे लिग्निफाई नहीं करते हैं और इसलिए प्लास्टिक हैं। लैक्टिस के मध्य भाग में लेटेक्स के साथ एक रिक्तिका होती है, और प्रोटोप्लास्ट एक निकट-दीवार की स्थिति में होता है। प्रोटोप्लास्ट और रसधानी के बीच की सीमा हमेशा निर्धारित नहीं की जा सकती है। लैक्टिफेर मेरिस्टेम में प्रवेश करते हैं और पौधे के अंग के साथ बढ़ते हैं।
प्रवाहकीय ऊतक
यह प्रकार जटिल ऊतकों को संदर्भित करता है, इसमें अलग-अलग विभेदित कोशिकाएं होती हैं। वास्तविक प्रवाहकीय तत्वों के अलावा, ऊतक में यांत्रिक, उत्सर्जन और भंडारण तत्व होते हैं (चित्र 26)। प्रवाहकीय ऊतक सभी पौधों के अंगों को एक प्रणाली में एकजुट करते हैं। प्रवाहकीय ऊतक दो प्रकार के होते हैं: जाइलम और फ्लोएम (यूनानी ज़ाइलॉन - पेड़; फ्लोयोस - छाल, बस्ट)। उनके पास संरचनात्मक और कार्यात्मक दोनों अंतर हैं।
जाइलम के संवाहक तत्व मृत कोशिकाओं द्वारा बनते हैं। वे जड़ से पत्तियों तक पानी और उसमें घुले पदार्थों का लंबी दूरी तक परिवहन करते हैं। फ्लोएम के प्रवाहकीय तत्व प्रोटोप्लास्ट को जीवित रखते हैं। वे प्रकाश संश्लेषक पत्तियों से जड़ तक लंबी दूरी का परिवहन करते हैं।
आमतौर पर जाइलम और फ्लोएम पौधे के शरीर में स्थित होते हैं निश्चित आदेश, परतें या प्रवाहकीय बंडल बनाते हैं। संरचना के आधार पर, कई प्रकार के प्रवाहकीय बंडल प्रतिष्ठित होते हैं, जिनकी विशेषता होती है कुछ समूहपौधे। जाइलम और फ्लोएम के बीच संपार्श्विक खुले बंडल में, एक कैम्बियम होता है, जो द्वितीयक वृद्धि प्रदान करता है (चित्र 27-ए, 28)। बाइकोलेटरल ओपन बंडल में, फ्लोएम जाइलम के दोनों किनारों पर स्थित होता है (चित्र 27-बी, 29)। बंद बंडलों में कैम्बियम नहीं होता है, और इसलिए वे द्वितीयक गाढ़ा करने में सक्षम नहीं होते हैं (चित्र 27-बी, 27-डी, 30.31)। दो और प्रकार के संकेंद्रित बंडल पाए जा सकते हैं, जहाँ या तो फ्लोएम जाइलम (चित्र 27-ई, 32) को घेरता है, या जाइलम फ्लोएम को घेरता है (चित्र 27-ई)।
जाइलम (लकड़ी)। उच्च पौधों में जाइलम का विकास जल विनिमय के प्रावधान से जुड़ा हुआ है। चूंकि एपिडर्मिस के माध्यम से पानी लगातार उत्सर्जित होता है, नमी की समान मात्रा को पौधे द्वारा अवशोषित किया जाना चाहिए और उन अंगों में जोड़ा जाना चाहिए जो वाष्पोत्सर्जन करते हैं। यह ध्यान में रखा जाना चाहिए कि जल-संचालन कोशिकाओं में एक जीवित प्रोटोप्लास्ट की उपस्थिति परिवहन को बहुत धीमा कर देगी, और मृत कोशिकाएं यहां अधिक कार्यात्मक हैं। हालांकि, एक मृत कोशिका में स्फीति नहीं होती है, इसलिए झिल्ली में यांत्रिक गुण होने चाहिए। नोट: मरोड़ पौधों की कोशिकाओं, ऊतकों और अंगों की अवस्था है, जिसमें वे अपनी लोचदार झिल्लियों पर कोशिकाओं की सामग्री के दबाव के कारण लोचदार हो जाते हैं। दरअसल, जाइलम के संवाहक तत्वों में मोटे लिग्निफाइड गोले के साथ अंग की धुरी के साथ फैली हुई मृत कोशिकाएं होती हैं।
प्रारंभ में, जाइलम प्राथमिक मेरिस्टेम - प्रोकैम्बियम से बनता है, जो अक्षीय अंगों के शीर्ष पर स्थित होता है। प्रोटोजाइलम पहले विभेदित करता है, उसके बाद मेटाजाइलम। तीन प्रकार के जाइलम गठन ज्ञात हैं। एक्सार्क प्रकार में, प्रोटोजाइलम के तत्व पहले प्रोकैम्बियल बंडल की परिधि पर दिखाई देते हैं, फिर मेटाजाइलम के तत्व केंद्र में दिखाई देते हैं। यदि प्रक्रिया विपरीत दिशा में जाती है (अर्थात केंद्र से परिधि तक), तो यह एक स्थायी प्रकार है। मेसार्चिक प्रकार में, जाइलम को प्रोकैम्बियल बंडल के केंद्र में रखा जाता है, जिसके बाद इसे केंद्र की ओर और परिधि की ओर जमा किया जाता है।
जड़ की विशेषता एक एक्सार्क प्रकार के जाइलम गठन से होती है, जबकि तनों में एक एंडार्क प्रकार होता है। कम संगठित पौधों में, जाइलम निर्माण के तरीके बहुत विविध होते हैं और व्यवस्थित विशेषताओं के रूप में काम कर सकते हैं।
कुछ पौधों में (उदाहरण के लिए, मोनोकॉट्स), सभी प्रकोष्ठीय कोशिकाएं प्रवाहकीय ऊतकों में अंतर करती हैं जो द्वितीयक गाढ़ा करने में सक्षम नहीं होते हैं। अन्य रूपों में (उदाहरण के लिए, वुडी), पार्श्व विभज्योतक (केंबियम) जाइलम और फ्लोएम के बीच रहते हैं। ये कोशिकाएँ जाइलम और फ्लोएम को विभाजित करने, नवीनीकृत करने में सक्षम हैं। इस प्रक्रिया को द्वितीयक वृद्धि कहते हैं। अपेक्षाकृत स्थिर जलवायु परिस्थितियों में उगने वाले कई पौधे लगातार बढ़ते हैं। मौसमी जलवायु परिवर्तन के अनुकूल रूपों में - समय-समय पर। नतीजतन, अच्छी तरह से परिभाषित वार्षिक वृद्धि के छल्ले बनते हैं।
प्रकोष्ठीय कोशिकाओं के विभेदन के मुख्य चरण। इसकी कोशिकाओं में पतली झिल्लियां होती हैं जो अंग के विकास के दौरान उन्हें फैलने से नहीं रोकती हैं। प्रोटोप्लास्ट तब द्वितीयक झिल्ली बिछाना शुरू कर देता है। लेकिन इस प्रक्रिया में स्पष्ट विशेषताएं हैं। द्वितीयक झिल्ली एक सतत परत में जमा नहीं होती है, जो कोशिका को फैलने नहीं देती, बल्कि छल्ले या सर्पिल के रूप में होती है। सेल बढ़ाव मुश्किल नहीं है। युवा कोशिकाओं में, हेलिक्स के छल्ले या कुंडली एक दूसरे के करीब स्थित होते हैं। परिपक्व कोशिकाओं में, वे कोशिका खिंचाव (चित्र 33) के परिणामस्वरूप विचलन करते हैं। खोल के चक्राकार और सर्पिल मोटा होना विकास को नहीं रोकता है, हालांकि, यंत्रवत् वे गोले से नीच होते हैं, जहां द्वितीयक मोटा होना एक सतत परत बनाता है। इसलिए, जाइलम में वृद्धि की समाप्ति के बाद, एक सतत लिग्नीफाइड शेल (मेटाक्साइलम) वाले तत्व बनते हैं। यह ध्यान दिया जाना चाहिए कि यहां द्वितीयक मोटा होना कुंडलाकार या सर्पिल नहीं है, बल्कि पंचर, सीढ़ी जैसा, जालीदार (चित्र 34) है। इसकी कोशिकाएं खिंचने में सक्षम नहीं होती हैं और कुछ ही घंटों में मर जाती हैं। आस-पास की कोशिकाओं में यह प्रक्रिया समन्वित तरीके से होती है। साइटोप्लाज्म में बड़ी संख्या में लाइसोसोम दिखाई देते हैं। फिर वे विघटित हो जाते हैं, और उनमें मौजूद एंजाइम प्रोटोप्लास्ट को नष्ट कर देते हैं। जब अनुप्रस्थ दीवारें नष्ट हो जाती हैं, तो एक के ऊपर एक श्रृंखला में स्थित कोशिकाएं एक खोखले बर्तन (चित्र 35) का निर्माण करती हैं। बहुमत आवृतबीजीऔर कुछ फर्न में बर्तन होते हैं।
एक संवाहक कोशिका जो इसकी दीवार में छिद्रों के माध्यम से नहीं बनती है, ट्रेकिड कहलाती है। वाहिकाओं के माध्यम से ट्रेकिड्स के माध्यम से पानी की गति धीमी होती है। तथ्य यह है कि ट्रेकिड्स में प्राथमिक झिल्ली कहीं भी बाधित नहीं होती है। ट्रेकिड्स एक दूसरे के साथ छिद्रों के माध्यम से संवाद करते हैं। यह स्पष्ट किया जाना चाहिए कि पौधों में, छिद्र प्राथमिक झिल्ली तक द्वितीयक झिल्ली में केवल एक अवसाद है, और ट्रेकिड्स के बीच छिद्रों के माध्यम से नहीं होते हैं।
सबसे आम बॉर्डर वाले छिद्र हैं (चित्र 35-1)। उनका चैनल, कोशिका की गुहा का सामना करना पड़ रहा है, एक विस्तार बनाता है - ताकना कक्ष। प्राथमिक खोल पर अधिकांश शंकुधारी पौधों के छिद्रों में एक मोटा होना होता है - एक टोरस, जो एक प्रकार का वाल्व होता है और जल परिवहन की तीव्रता को नियंत्रित करने में सक्षम होता है। स्थानांतरित करके, टोरस छिद्र के माध्यम से पानी के प्रवाह को अवरुद्ध करता है, लेकिन उसके बाद यह एक बार की कार्रवाई करते हुए अपनी पिछली स्थिति में वापस नहीं आ सकता है।
छिद्र अधिक या कम गोल होते हैं, लम्बी धुरी के लंबवत होते हैं (इन छिद्रों का एक समूह सीढ़ी जैसा दिखता है, इसलिए इस छिद्र को सीढ़ी छिद्र कहा जाता है)। छिद्रों के माध्यम से परिवहन अनुदैर्ध्य और अनुप्रस्थ दोनों दिशाओं में किया जाता है। छिद्र न केवल ट्रेकिड्स में मौजूद होते हैं, बल्कि एक पोत बनाने वाली व्यक्तिगत संवहनी कोशिकाओं में भी मौजूद होते हैं।
विकासवादी सिद्धांत के दृष्टिकोण से, ट्रेकिड्स पहली और मुख्य संरचना है जो उच्च पौधों के शरीर में पानी का संचालन करती है। ऐसा माना जाता है कि जहाजों की उत्पत्ति उनके बीच की अनुप्रस्थ दीवारों (चित्र 36) के लसीका के कारण ट्रेकिड्स से हुई थी। अधिकांश फ़र्न और जिम्नोस्पर्म में नहीं होता है। वे ट्रेकिड्स के माध्यम से पानी ले जाते हैं।
विकासवादी विकास के क्रम में, जहाजों का उदय हुआ विभिन्न समूहपौधे बार-बार, लेकिन उन्होंने एंजियोस्पर्म में सबसे महत्वपूर्ण कार्यात्मक महत्व हासिल किया, जो उन्हें ट्रेकिड्स के साथ मिला है। यह माना जाता है कि परिवहन के अधिक उन्नत तंत्र के कब्जे ने उन्हें न केवल जीवित रहने में मदद की, बल्कि महत्वपूर्ण रूपों को प्राप्त करने में भी मदद की।
जाइलम एक जटिल ऊतक है, इसमें जल-संवाहक तत्वों के अलावा अन्य तत्व भी होते हैं। यांत्रिक कार्य लाइब्रिफॉर्म फाइबर (अव्य। लिबर - बास्ट, फॉर्मा - फॉर्म) द्वारा किए जाते हैं। अतिरिक्त यांत्रिक संरचनाओं की उपस्थिति महत्वपूर्ण है, क्योंकि पानी ले जाने वाले तत्वों की दीवारें मोटी होने के बावजूद अभी भी बहुत पतली हैं। वे खुद को बनाए रखने में असमर्थ हैं एक बड़ा द्रव्यमानबारहमासी पौधा। ट्रेकिड्स से तंतु विकसित होते हैं। वे छोटे आकार, लिग्नीफाइड (लिग्नीफाइड) गोले और संकीर्ण गुहाओं की विशेषता हैं। दीवार पर किनारों से रहित छिद्र पाए जा सकते हैं। ये रेशे पानी का संचालन नहीं कर सकते, इनका मुख्य कार्य आधार है।
जाइलम में जीवित कोशिकाएं भी होती हैं। उनका द्रव्यमान लकड़ी की कुल मात्रा का 25% तक पहुंच सकता है। चूँकि इन कोशिकाओं का आकार गोल होता है, इसलिए इन्हें काष्ठ पैरेन्काइमा कहा जाता है। पैरेन्काइमा पौधे के शरीर में दो तरह से स्थित होता है। पहले मामले में, कोशिकाओं को ऊर्ध्वाधर डोरियों के रूप में व्यवस्थित किया जाता है - यह कॉर्ड पैरेन्काइमा है। एक अन्य मामले में, पैरेन्काइमा क्षैतिज किरणें बनाता है। उन्हें कोर किरणें कहा जाता है, क्योंकि वे कोर और कॉर्टेक्स को जोड़ती हैं। कोर पदार्थों के भंडारण सहित कई कार्य करता है।
फ्लोएम (बस्ट)। यह एक जटिल ऊतक है, क्योंकि यह विषम कोशिकाओं द्वारा बनता है। मुख्य संवाहक कोशिकाओं को छलनी तत्व कहा जाता है (चित्र 37)। जाइलम के संवाहक तत्व मृत कोशिकाओं द्वारा बनते हैं, जबकि फ्लोएम में वे कार्य करने की अवधि के दौरान जीवित रहते हैं, हालांकि बहुत बदल जाते हैं, प्रोटोप्लास्ट। फ्लोएम प्रकाश संश्लेषक अंगों से प्लास्टिक पदार्थों का बहिर्वाह करता है। पौधों की सभी जीवित कोशिकाओं में कार्बनिक पदार्थों के संचालन की क्षमता होती है। और इसलिए, यदि जाइलम केवल उच्च पौधों में पाया जा सकता है, तो कोशिकाओं के बीच कार्बनिक पदार्थों का परिवहन भी किसमें होता है? निचले पौधे.
जाइलम और फ्लोएम एपिकल मेरिस्टेम से विकसित होते हैं। पहले चरण में प्रोटोफ्लोएम प्रोकैम्बियल कॉर्ड में बनता है। जैसे-जैसे आसपास के ऊतक बढ़ते हैं, यह फैलता है, और जब विकास पूरा हो जाता है, तो प्रोटोफ्लोएम के बजाय मेटाफ्लोएम बनता है।
पर विभिन्न समूहउच्च पौधों में दो प्रकार के छलनी तत्व पाए जाते हैं। फ़र्न और जिम्नोस्पर्म में, यह छलनी कोशिकाओं द्वारा दर्शाया जाता है। कोशिकाओं में छलनी के क्षेत्र बगल की दीवारों के साथ बिखरे हुए हैं। प्रोटोप्लास्ट थोड़ा नष्ट हुए नाभिक को बरकरार रखता है।
एंजियोस्पर्म में, छलनी तत्वों को चालनी ट्यूब कहा जाता है। वे चलनी प्लेटों के माध्यम से एक दूसरे के साथ संवाद करते हैं। परिपक्व कोशिकाओं में केन्द्रक का अभाव होता है। हालाँकि, एक साथी कोशिका छलनी नली के बगल में स्थित होती है, जो सामान्य माँ कोशिका (चित्र 38) के माइटोटिक विभाजन के परिणामस्वरूप छलनी नली के साथ मिलकर बनती है। साथी कोशिका में सघन साइटोप्लाज्म होता है जिसमें बड़ी संख्या में सक्रिय माइटोकॉन्ड्रिया होते हैं, साथ ही एक पूरी तरह से काम करने वाला नाभिक भी होता है। बड़ी राशिप्लास्मोडेस्माटा (अन्य कोशिकाओं की तुलना में दस गुना अधिक)। साथी कोशिकाएं गैर-परमाणु चलनी ट्यूब कोशिकाओं की कार्यात्मक गतिविधि को प्रभावित करती हैं।
परिपक्व छलनी कोशिकाओं की संरचना में कुछ ख़ासियतें होती हैं। कोई रिक्तिका नहीं है, इसलिए साइटोप्लाज्म अत्यधिक द्रवीभूत होता है। केंद्रक अनुपस्थित हो सकता है (एंजियोस्पर्म में) या झुर्रीदार, कार्यात्मक रूप से निष्क्रिय अवस्था में हो सकता है। राइबोसोम और गोल्गी कॉम्प्लेक्स भी अनुपस्थित हैं, लेकिन एंडोप्लाज्मिक रेटिकुलम अच्छी तरह से विकसित है, जो न केवल साइटोप्लाज्म में प्रवेश करता है, बल्कि छलनी क्षेत्रों के छिद्रों के माध्यम से पड़ोसी कोशिकाओं में भी जाता है। अच्छी तरह से विकसित माइटोकॉन्ड्रिया और प्लास्टिड बहुतायत में पाए जाते हैं।
कोशिकाओं के बीच, पदार्थों का परिवहन कोशिका झिल्लियों पर स्थित छिद्रों से होता है। इस तरह के उद्घाटन को छिद्र कहा जाता है, लेकिन ट्रेकिड छिद्रों के विपरीत, वे होते हैं। यह माना जाता है कि वे अत्यधिक विस्तारित प्लास्मोडेस्माटा हैं, जिनकी दीवारों पर कॉलोस पॉलीसेकेराइड जमा होता है। छिद्रों को समूहों में व्यवस्थित किया जाता है, छलनी क्षेत्र बनाते हैं। आदिम रूपों में, छलनी के क्षेत्र खोल की पूरी सतह पर बेतरतीब ढंग से बिखरे हुए हैं, अधिक उन्नत एंजियोस्पर्म में वे एक दूसरे से सटे पड़ोसी कोशिकाओं के सिरों पर स्थित होते हैं, जो एक चलनी प्लेट (चित्र 39) बनाते हैं। यदि उस पर एक छलनी का क्षेत्र है, तो इसे सरल कहा जाता है, यदि कई - जटिल हैं।
चलनी तत्वों के माध्यम से समाधानों की गति की गति 150 सेमी ∕ घंटे तक है। यह मुक्त प्रसार की दर का एक हजार गुना है। संभवतः, सक्रिय परिवहन होता है, और छलनी तत्वों और उपग्रह कोशिकाओं के कई माइटोकॉन्ड्रिया इसके लिए आवश्यक एटीपी की आपूर्ति करते हैं।
फ्लोएम के छलनी तत्वों का जीवन पार्श्व विभज्योतकों की उपस्थिति पर निर्भर करता है। यदि वे मौजूद हैं, तो छलनी तत्व पौधे के जीवन भर काम करते हैं।
छलनी तत्वों और उपग्रह कोशिकाओं के अलावा, फ्लोएम में बस्ट फाइबर, स्केलेरिड्स और पैरेन्काइमा होते हैं।
नियंत्रण और पुनरावृत्ति के लिए प्रश्न
उच्च पौधों के लक्षण? उच्च पौधों की किस पीढ़ी को गैमेटोफाइट कहा जाता है? स्पोरोफाइट?
वे कौन से कारण हैं जो स्थलीय पौधों में विभेदित ऊतकों के विकास में योगदान करते हैं।
पौधों के प्रकारों की सूची बनाइए।
आप विभज्योतकों के किन समूहों को जानते हैं? वे पौधे के शरीर में कहाँ स्थित हैं?
पूर्णांक ऊतक, उनके कार्य, महत्व।
संगठन विभिन्न प्रकार केऊतकों को ढंकना।
रंध्र तंत्र की संरचना। आपको ज्ञात रंध्र उपकरण के प्रकारों की सूची बनाएं। रंध्र कैसे कार्य करते हैं?
पैरेन्काइमल ऊतकों की विशेषताएं, पैरेन्काइमा के प्रकार।
पैरेन्काइमा और स्क्लेरेन्काइमा के लक्षण।
पौधों की उत्सर्जी संरचनाओं के प्रकारों की सूची बनाइए। उनका क्या महत्व है?
उत्सर्जित पदार्थों के लिए पात्र बनाने की विधियाँ।
दूधवाले क्या होते हैं? वे किस प्रकार में विभाजित हैं?
प्रवाहकीय ऊतक कैसे व्यवस्थित होते हैं?
प्रवाहकीय बंडल क्या हैं? उनके प्रकार।
जाइलम में पदार्थों की गति की दिशा? फ्लोएम? क्या परिवहन किए गए पदार्थों की संरचना जाइलम और फ्लोएम में भिन्न होती है?
जाइलम, फ्लोएम की संरचना और कार्य।
विभिन्न से पौधों में फ्लोएम के संचालन तत्वों का संगठन व्यवस्थित समूह.
फ्लोएम में पदार्थों का परिवहन कैसे होता है?
उनसे जुड़े अतिरिक्त प्रोटीन, तथाकथित एमएपी प्रोटीन। ये प्रोटीन, सूक्ष्मनलिकाएं को स्थिर करके, ट्यूबुलिन पोलीमराइजेशन की प्रक्रिया को तेज करते हैं। माइक्रोफिलामेंट्स की संरचना और कार्य माइक्रोफिलामेंट्स बहुत पतली और लंबी फिलामेंटस प्रोटीन संरचनाएं हैं जो पूरे साइटोप्लाज्म में पाई जाती हैं। प्लाज्मा झिल्ली के नीचे, माइक्रोफ़िल्मेंट्स एक सतत प्लेक्सस बनाते हैं, जिससे ...

जैसे, उदाहरण के लिए, डिल, गाजर, शलजम, आदि, ज्यादातर मामलों में इसमें उसी तरह का ऊतक होता है जैसा मैंने अभी कॉर्क में दिखाया था। इस प्रकार पहली बार पादप कोशिका की खोज हुई। लेकिन हूक अपने माइक्रोस्कोप के साथ लंबे समय तक नहीं निपट सका: अन्य आविष्कारों के विचार उसके सिर (वसंत घड़ियां, बेहतर कम्पास, आदि) में तैर गए, और उसने स्वेच्छा से सूक्ष्मदर्शी के आगे के रखरखाव को सौंप दिया ...
सेल अनुसंधान के उपयुक्त तरीकों का कम से कम कुछ विचार। प्रकाश माइक्रोस्कोपी सबसे प्राचीन और, एक ही समय में, कोशिकाओं का अध्ययन करने का सबसे आम तरीका माइक्रोस्कोपी है। हम कह सकते हैं कि प्रकाश प्रकाशीय सूक्ष्मदर्शी के आविष्कार से कोशिका के अध्ययन की शुरुआत हुई। निहत्था मनुष्य की आंखलगभग 1/10 मिमी का संकल्प है। यह...
कई सेल ऑर्गेनेल के साथ, यह व्यक्तिगत इंट्रासेल्युलर संरचनाओं के बीच एक संबंध प्रदान करता है। लाइसोसोम लाइसोसोम (लिस- और ग्रीक से। सोमा - शरीर), जानवरों और पौधों के जीवों की कोशिकाओं में संरचनाएं जिनमें एंजाइम होते हैं जो टूट सकते हैं (यानी लाइसे - इसलिए नाम) प्रोटीन, पॉलीसेकेराइड, पेप्टाइड्स, न्यूक्लिक एसिड। यह 0.1-0.4 माइक्रोन के आकार के बुलबुले का एक बहुत ही भिन्न वर्ग है, ...
पादप कोशिका की संरचना। पौधे के ऊतक
प्लांट सेल की संरचना
एक पादप कोशिका में कम या ज्यादा कठोर कोशिका भित्ति और एक प्रोटोप्लास्ट होता है। कोशिका भित्ति कोशिका भित्ति और साइटोप्लाज्मिक झिल्ली है। प्रोटोप्लास्ट शब्द प्रोटोप्लाज्म शब्द से आया है, जिसका उपयोग लंबे समय से सभी जीवित चीजों को संदर्भित करने के लिए किया जाता है। प्रोटोप्लास्ट एक व्यक्तिगत कोशिका का प्रोटोप्लाज्म है।
प्रोटोप्लास्ट में साइटोप्लाज्म और न्यूक्लियस होते हैं। साइटोप्लाज्म में ऑर्गेनेल (राइबोसोम, माइक्रोट्यूबुल्स, प्लास्टिड्स, माइटोकॉन्ड्रिया) और मेम्ब्रेन सिस्टम (एंडोप्लाज्मिक रेटिकुलम, डिक्टीयोसोम) होते हैं। साइटोप्लाज्म में साइटोप्लाज्मिक मैट्रिक्स (मूल पदार्थ) भी शामिल होता है जिसमें ऑर्गेनेल और मेम्ब्रेन सिस्टम डूबे रहते हैं। साइटोप्लाज्म को कोशिका भित्ति से प्लाज्मा झिल्ली द्वारा अलग किया जाता है, जो एक प्राथमिक झिल्ली है। अधिकांश पशु कोशिकाओं के विपरीत, पादप कोशिकाओं में एक या एक से अधिक रिक्तिकाएँ होती हैं। ये पुटिकाएं तरल से भरी होती हैं और एक प्रारंभिक झिल्ली (टोनोप्लास्ट) से घिरी होती हैं।
एक जीवित पादप कोशिका में, जमीनी पदार्थ निरंतर गति में रहता है। ऑर्गेनेल आंदोलन में शामिल होते हैं, जिसे साइटोप्लाज्म या साइक्लोसिस का प्रवाह कहा जाता है। साइक्लोसिस कोशिका में पदार्थों की आवाजाही और कोशिका और पर्यावरण के बीच उनके आदान-प्रदान की सुविधा प्रदान करता है।
प्लाज्मा झिल्ली।यह एक बाइलेयर फॉस्फोलिपिड संरचना है। पादप कोशिकाओं की विशेषता प्लाज्मा झिल्ली के अंतर्वलन से होती है।
प्लाज्मा झिल्ली निम्नलिखित कार्य करती है:
कोशिका और पर्यावरण के बीच पदार्थों के आदान-प्रदान में भाग लेता है;
कोशिका भित्ति के सेल्युलोज सूक्ष्मतंतुओं के संश्लेषण और संयोजन का समन्वय करता है;
हार्मोनल और बाहरी संकेतों को प्रसारित करता है जो सेल के विकास और भेदभाव को नियंत्रित करते हैं।
मुख्य।यह यूकेरियोटिक कोशिका के साइटोप्लाज्म में सबसे प्रमुख संरचना है। कर्नेल दो महत्वपूर्ण कार्य करता है:
कोशिका की महत्वपूर्ण गतिविधि को नियंत्रित करता है, यह निर्धारित करता है कि कौन से प्रोटीन और किस समय संश्लेषित किए जाने चाहिए;
आनुवंशिक जानकारी को संग्रहीत करता है और इसे कोशिका विभाजन के दौरान संतति कोशिकाओं तक पहुंचाता है।
एक यूकेरियोटिक कोशिका का केंद्रक दो प्राथमिक झिल्लियों से घिरा होता है जो परमाणु आवरण बनाती हैं। यह 30 से 100 एनएम के व्यास के साथ कई छिद्रों से व्याप्त है, जो केवल एक इलेक्ट्रॉन माइक्रोस्कोप में दिखाई देता है। छिद्रों की एक जटिल संरचना होती है। कुछ स्थानों पर परमाणु झिल्ली की बाहरी झिल्ली को एंडोप्लाज्मिक रेटिकुलम के साथ जोड़ा जाता है। परमाणु लिफाफे को एंडोप्लाज्मिक रेटिकुलम (ईआर) के एक विशेष, स्थानीय रूप से विभेदित भाग के रूप में देखा जा सकता है।
विशेष रंगों से सना हुआ नाभिक में, क्रोमैटिन और न्यूक्लियोप्लाज्म (नाभिक का मुख्य पदार्थ) के पतले तंतुओं और गुच्छों को प्रतिष्ठित किया जा सकता है। क्रोमेटिन हिस्टोन नामक विशेष प्रोटीन से बंधे डीएनए से बना होता है। कोशिका विभाजन की प्रक्रिया में, क्रोमैटिन अधिक से अधिक संकुचित हो जाता है और गुणसूत्रों में इकट्ठा हो जाता है। डीएनए आनुवंशिक जानकारी को एनकोड करता है।
दैहिक कोशिकाओं में जीव गुणसूत्रों की संख्या में भिन्न होते हैं। उदाहरण के लिए, गोभी में - 20 गुणसूत्र होते हैं; सूरजमुखी - 34; गेहूँ - 42; मनुष्य - 46, और फ़र्न ओफ़ियोग्लोसम की प्रजातियों में से एक - 1250। सेक्स कोशिकाओं (युग्मकों) में शरीर की दैहिक कोशिकाओं के गुणसूत्रों की संख्या का केवल आधा हिस्सा होता है। युग्मकों में गुणसूत्रों की संख्या को अगुणित (एकल) कहा जाता है, दैहिक कोशिकाओं में - द्विगुणित (डबल)। जिन कोशिकाओं में गुणसूत्रों के दो से अधिक सेट होते हैं उन्हें पॉलीप्लोइड कहा जाता है।
एक प्रकाश सूक्ष्मदर्शी के तहत, गोलाकार संरचनाएं - न्यूक्लियोली - देखी जा सकती हैं। प्रत्येक नाभिक में एक या एक से अधिक नाभिक होते हैं, जो अविभाजित नाभिक में दिखाई देते हैं। राइबोसोमल आरएनए को न्यूक्लियोलस में संश्लेषित किया जाता है। आमतौर पर, द्विगुणित जीवों के नाभिक में दो नाभिक होते हैं, गुणसूत्रों के प्रत्येक अगुणित सेट के लिए एक। न्यूक्लियोली की अपनी झिल्ली नहीं होती है। बायोकेमिकल रूप से, न्यूक्लियोली को आरएनए की उच्च सांद्रता की विशेषता होती है, जो यहां फॉस्फोप्रोटीन के साथ जुड़ा हुआ है। नाभिक का आकार कोशिका की कार्यात्मक अवस्था पर निर्भर करता है। यह देखा गया है कि एक तेजी से बढ़ने वाली कोशिका में, जिसमें प्रोटीन संश्लेषण की गहन प्रक्रियाएँ हो रही होती हैं, नाभिक का आकार बढ़ जाता है। न्यूक्लियोली में, एमआरएनए और राइबोसोम उत्पन्न होते हैं, जो केवल न्यूक्लियस में सिंथेटिक कार्य करते हैं।
न्यूक्लियोप्लाज्म (कार्योप्लाज्म) एक सजातीय तरल द्वारा दर्शाया जाता है जिसमें एंजाइम सहित विभिन्न प्रोटीन घुल जाते हैं।
प्लास्टिड्स।रिक्तिकाएं, सेल्युलोज कोशिका भित्ति और प्लास्टिड पादप कोशिकाओं के विशिष्ट घटक हैं। प्रत्येक प्लास्टिड का अपना खोल होता है, जिसमें दो प्राथमिक झिल्लियाँ होती हैं। प्लास्टिड के अंदर, एक झिल्ली प्रणाली और एक सजातीय पदार्थ की अलग-अलग डिग्री, स्ट्रोमा, प्रतिष्ठित हैं। परिपक्व प्लास्टिड्स को उनमें मौजूद पिगमेंट के आधार पर वर्गीकृत किया जाता है।
क्लोरोप्लास्ट, जिसमें प्रकाश संश्लेषण होता है, में क्लोरोफिल और कैरोटीनॉयड होते हैं। आमतौर पर 4 - 5 माइक्रोन के व्यास वाली डिस्क का रूप होता है। मेसोफिल (पत्ती के मध्य) की एक कोशिका में 40-50 क्लोरोप्लास्ट हो सकते हैं; पत्ती के मिमी 2 में - लगभग 500,000। साइटोप्लाज्म में, क्लोरोप्लास्ट आमतौर पर कोशिका झिल्ली के समानांतर स्थित होते हैं।
क्लोरोप्लास्ट की आंतरिक संरचना जटिल है। स्ट्रोमा को पुटिकाओं - थायलाकोइड्स के रूप में झिल्लियों की एक विकसित प्रणाली द्वारा अनुमति दी जाती है। प्रत्येक थायलाकोइड में दो झिल्ली होते हैं। थायलाकोइड्स एकल प्रणाली बनाते हैं। एक नियम के रूप में, वे ढेर में एकत्र किए जाते हैं - अनाज, सिक्कों के स्तंभों जैसा। अलग-अलग ग्रैने के थायलाकोइड्स स्ट्रोमा थायलाकोइड्स, या इंटरग्रेनल थायलाकोइड्स द्वारा परस्पर जुड़े होते हैं। क्लोरोफिल और कैरोटीनॉयड थायलाकोइड झिल्ली में एम्बेडेड होते हैं। हरे पौधों और शैवाल में क्लोरोप्लास्ट में अक्सर स्टार्च के दाने और छोटे लिपिड (वसा) की बूंदें होती हैं। स्टार्च के दाने प्रकाश संश्लेषण उत्पादों के अस्थायी भंडार होते हैं। वे केवल 24 घंटों के लिए अंधेरे में रहने वाले पौधे के क्लोरोप्लास्ट से गायब हो सकते हैं, और पौधे को प्रकाश में स्थानांतरित करने के 3-4 घंटे बाद फिर से प्रकट हो सकते हैं।
क्लोरोप्लास्ट अर्ध-स्वायत्त अंगक होते हैं और बैक्टीरिया के समान होते हैं। उदाहरण के लिए, बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम में काफी उच्च समानता होती है। वे यूकेरियोटिक राइबोसोम से छोटे होते हैं। बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम पर प्रोटीन संश्लेषण क्लोरैम्फेनिकॉल द्वारा बाधित होता है, जिसका यूकेरियोटिक कोशिकाओं पर कोई प्रभाव नहीं पड़ता है। इसके अलावा, बैक्टीरिया और क्लोरोप्लास्ट दोनों में एक समान प्रकार का न्यूक्लियॉइड होता है, जो समान तरीके से व्यवस्थित होता है। इस तथ्य के बावजूद कि क्लोरोप्लास्ट का निर्माण और उनमें निहित पिगमेंट का संश्लेषण काफी हद तक कोशिका के क्रोमोसोमल डीएनए द्वारा नियंत्रित होता है, फिर भी, अपने स्वयं के डीएनए की अनुपस्थिति में, क्लोरोप्लास्ट नहीं बनते हैं।
क्लोरोप्लास्ट को मुख्य सेलुलर अंग माना जा सकता है, क्योंकि वे सौर ऊर्जा के परिवर्तन की श्रृंखला में पहले हैं, जिसके परिणामस्वरूप मानवता को भोजन और ईंधन दोनों प्राप्त होते हैं। क्लोरोप्लास्ट में न केवल प्रकाश संश्लेषण होता है। वे अमीनो एसिड और फैटी एसिड के संश्लेषण में भी शामिल हैं, स्टार्च के अस्थायी भंडार के भंडार के रूप में काम करते हैं।
क्रोमोप्लास्ट रंजित प्लास्टिड हैं। विविध रूप में, उनके पास क्लोरोफिल नहीं है, लेकिन कैरोटीनॉयड को संश्लेषित और संचित करते हैं, जो फूलों, पुरानी पत्तियों, फलों और जड़ों को पीला, नारंगी, लाल रंग देते हैं। क्रोमोप्लास्ट क्लोरोप्लास्ट से विकसित हो सकते हैं, जो इस मामले में क्लोरोफिल और आंतरिक झिल्ली संरचनाओं को खो देते हैं, कैरोटीनॉयड जमा करते हैं। यह कई फलों के पकने के दौरान होता है। क्रोमोप्लास्ट कीड़ों और अन्य जानवरों को आकर्षित करते हैं जिनके साथ वे विकसित हुए हैं।
ल्यूकोप्लास्ट गैर-रंजित प्लास्टिड हैं। उनमें से कुछ स्टार्च (एमाइलोप्लास्ट) को संश्लेषित करते हैं, अन्य लिपिड और प्रोटीन सहित विभिन्न पदार्थ बनाने में सक्षम हैं। प्रकाश में, ल्यूकोप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं।
प्रोप्लास्टिड्स छोटे, रंगहीन या हल्के हरे रंग के, अविभाजित प्लास्टिड्स होते हैं जो जड़ों और अंकुरों की मेरिस्टेमेटिक (विभाजित) कोशिकाओं में पाए जाते हैं। वे अन्य, अधिक विभेदित प्लास्टिड्स - क्लोरोप्लास्ट्स, क्रोमोप्लास्ट्स और एमिनोप्लास्ट्स के पूर्ववर्ती हैं। यदि प्रकाश की कमी के कारण प्रोटोप्लास्टिड्स के विकास में देरी हो रही है, तो उनमें एक या एक से अधिक प्रोलामेलर निकाय दिखाई दे सकते हैं, जो ट्यूबलर झिल्ली के अर्ध-क्रिस्टलीय संचय हैं। प्रोलामेलर बॉडी वाले प्लास्टिड्स को इटियोप्लास्ट्स कहा जाता है। प्रकाश में, एटिओप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं, जबकि प्रोलामेलर निकायों की झिल्लियां थायलाकोइड्स बनाती हैं। इटियोप्लास्ट पौधों की पत्तियों में बनते हैं जो अंधेरे में होते हैं। बीज भ्रूण के प्रोटोप्लास्ट पहले इटियोप्लास्ट में बदलते हैं, जिससे क्लोरोप्लास्ट प्रकाश में विकसित होते हैं। प्लास्टिड्स को एक प्रकार से दूसरे प्रकार में अपेक्षाकृत आसान संक्रमणों की विशेषता है। प्लास्टिड्स, बैक्टीरिया की तरह, दो में विभाजित करके प्रजनन करते हैं। मेरिस्टेमेटिक कोशिकाओं में, प्रोटोप्लास्टिड्स के विभाजन का समय लगभग कोशिका विभाजन के समय के साथ मेल खाता है। हालांकि, परिपक्व कोशिकाओं में, अधिकांश प्लास्टिड परिपक्व प्लास्टिड्स के विभाजन के परिणामस्वरूप बनते हैं।
माइटोकॉन्ड्रिया।क्लोरोप्लास्ट की तरह, माइटोकॉन्ड्रिया दो प्राथमिक झिल्लियों से घिरा होता है। आंतरिक झिल्ली कई सिलवटों और प्रोट्रूशियंस - cristae का निर्माण करती है, जो माइटोकॉन्ड्रिया की आंतरिक सतह को काफी बढ़ा देती है। वे प्लास्टिड्स से काफी छोटे हैं, व्यास में लगभग 0.5 माइक्रोन और लंबाई और आकार में परिवर्तनशील हैं।
माइटोकॉन्ड्रिया में, श्वसन की प्रक्रिया होती है, जिसके परिणामस्वरूप कार्बनिक अणु ऊर्जा की रिहाई के साथ टूट जाते हैं और एटीपी अणुओं में स्थानांतरित हो जाते हैं, जो सभी यूकेरियोटिक कोशिकाओं का मुख्य ऊर्जा भंडार है। अधिकांश पादप कोशिकाओं में सैकड़ों या हजारों माइटोकॉन्ड्रिया होते हैं। एक सेल में उनकी संख्या सेल की एटीपी की आवश्यकता से निर्धारित होती है। माइटोकॉन्ड्रिया निरंतर गति में हैं, कोशिका के एक भाग से दूसरे भाग में जाते हैं, एक दूसरे के साथ विलय और विभाजित होते हैं। माइटोकॉन्ड्रिया आमतौर पर वहां इकट्ठा होते हैं जहां ऊर्जा की जरूरत होती है। यदि प्लाज़्मा झिल्ली सक्रिय रूप से कोशिका से कोशिका में पदार्थों को स्थानांतरित करती है, तो माइटोकॉन्ड्रिया झिल्ली की सतह के साथ स्थित होते हैं। मोटिव एककोशिकीय शैवाल में, माइटोकॉन्ड्रिया फ्लैगेल्ला के आधार पर जमा होता है, जो उनके आंदोलन के लिए आवश्यक ऊर्जा की आपूर्ति करता है।
माइटोकॉन्ड्रिया, प्लास्टिड्स की तरह, अर्ध-स्वायत्त अंग होते हैं जिनमें अपने स्वयं के प्रोटीन के संश्लेषण के लिए आवश्यक घटक होते हैं। आंतरिक झिल्ली एक द्रव मैट्रिक्स को घेरती है जिसमें प्रोटीन, आरएनए, डीएनए, बैक्टीरिया जैसे राइबोसोम और विभिन्न विलेय होते हैं। डीएनए एक या एक से अधिक न्यूक्लियॉइड्स में स्थित गोलाकार अणुओं के रूप में मौजूद होता है।
माइटोकॉन्ड्रिया और यूकेरियोटिक कोशिकाओं के क्लोरोप्लास्ट के साथ बैक्टीरिया की समानता के आधार पर, यह माना जा सकता है कि माइटोकॉन्ड्रिया और क्लोरोप्लास्ट बैक्टीरिया से उत्पन्न हुए हैं जो बड़े हेटरोट्रॉफ़िक कोशिकाओं में "शरण" पाए गए - यूकेरियोट्स के अग्रदूत।
माइक्रोबॉडीज।प्लास्टिड्स और माइटोकॉन्ड्रिया के विपरीत, जो दो झिल्लियों द्वारा सीमांकित होते हैं, माइक्रोबॉडी एक झिल्ली से घिरे गोलाकार अंग होते हैं। माइक्रोबॉडी में एक दानेदार (दानेदार) सामग्री होती है, कभी-कभी उनमें क्रिस्टलीय प्रोटीन का समावेश भी होता है। माइक्रोबॉडी एंडोप्लाज्मिक रेटिकुलम के एक या दो क्षेत्रों से जुड़े होते हैं।
कुछ माइक्रोबॉडी, जिन्हें प्रॉक्सिसोम कहा जाता है, ग्लाइकोलिक एसिड के चयापचय में एक महत्वपूर्ण भूमिका निभाते हैं, जो सीधे तौर पर प्रकाश श्वसन से संबंधित है। हरी पत्तियों में, वे माइटोकॉन्ड्रिया और क्लोरोप्लास्ट से जुड़े होते हैं। अन्य माइक्रोबॉडीज, जिन्हें ग्लायॉक्सिसोम कहा जाता है, में वसा को कार्बोहाइड्रेट में बदलने के लिए आवश्यक एंजाइम होते हैं। यह कई बीजों में अंकुरण के दौरान होता है।
रिक्तिकाएं -ये झिल्ली द्वारा परिसीमित कोशिका के क्षेत्र हैं, जो तरल - कोशिका रस से भरे होते हैं। वे एक टोनोप्लास्ट (वैक्यूलर मेम्ब्रेन) से घिरे होते हैं।
एक युवा पादप कोशिका में कई छोटे रिक्तिकाएँ होती हैं, जो कोशिका आयु के रूप में एक बड़ी रसधानी में विलीन हो जाती हैं। एक परिपक्व कोशिका में, इसकी मात्रा का 90% तक रिक्तिका द्वारा कब्जा किया जा सकता है। इस मामले में, साइटोप्लाज्म कोशिका झिल्ली को एक पतली परिधीय परत के रूप में दबाया जाता है। कोशिका आकार में वृद्धि मुख्य रूप से रसधानी की वृद्धि के कारण होती है। नतीजतन, टर्गर दबाव उत्पन्न होता है और ऊतक लोच बनाए रखा जाता है। यह रिक्तिका और टोनोप्लास्ट के मुख्य कार्यों में से एक है।
रस का मुख्य घटक पानी है, बाकी पौधे के प्रकार और उसकी शारीरिक अवस्था के आधार पर भिन्न होता है। रिक्तिका में लवण, शर्करा, कम अक्सर प्रोटीन होते हैं। टोनोप्लास्ट रिक्तिका में कुछ आयनों के परिवहन और संचय में सक्रिय भूमिका निभाता है। सेल सैप में आयनों की सांद्रता पर्यावरण में इसकी सांद्रता से काफी अधिक हो सकती है। कुछ पदार्थों की उच्च सामग्री के साथ, रिक्तिका में क्रिस्टल बनते हैं। सबसे आम कैल्शियम ऑक्सालेट क्रिस्टल होते हैं, जिनका एक अलग आकार होता है।
रिक्तिकाएं चयापचय उत्पादों (चयापचय) के संचय के स्थान हैं। ये प्रोटीन, अम्ल और यहां तक कि मनुष्य के लिए विषाक्त पदार्थ (एल्कलॉइड) भी हो सकते हैं। पिगमेंट अक्सर जमा होते हैं। नीला, बैंगनी, बैंगनी, गहरा लाल, किरमिजी पौधे की कोशिकाओं को एंथोसायनिन समूह से वर्णक देते हैं। अन्य पिगमेंट के विपरीत, वे पानी में अच्छी तरह से घुल जाते हैं और सेल सैप में पाए जाते हैं। वे कई सब्जियों (मूली, शलजम, गोभी), फल (अंगूर, आलूबुखारा, चेरी), फूल (कॉर्नफ्लॉवर, जेरेनियम, डेल्फीनियम, गुलाब, peonies) के लाल और नीले रंग का निर्धारण करते हैं। कभी-कभी ये वर्णक पत्तियों में क्लोरोफिल को ढक लेते हैं, उदाहरण के लिए, सजावटी लाल मेपल में। एंथोसायनिन रंग शरद ऋतु चमकदार लाल छोड़ देता है। वे ठंडी धूप के मौसम में बनते हैं, जब पत्तियों में क्लोरोफिल का संश्लेषण बंद हो जाता है। पत्तियों में, जब एंथोसायनिन नहीं बनते हैं, क्लोरोफिल के विनाश के बाद, क्लोरोप्लास्ट के पीले-नारंगी कैरोटीनॉयड ध्यान देने योग्य हो जाते हैं। ठंडे, स्पष्ट शरद ऋतु में पत्तियाँ सबसे चमकीले रंग की होती हैं।
कोशिका में उनके घटकों के संचलन में, रिक्तिकाएं मैक्रोमोलेक्युलस के विनाश में शामिल होती हैं। राइबोसोम, माइटोकॉन्ड्रिया, प्लास्टिड, रिक्तिका में गिरकर नष्ट हो जाते हैं। इस पाचन गतिविधि के अनुसार, उनकी तुलना लाइसोसोम - पशु कोशिकाओं के अंग से की जा सकती है।
एंडोप्लाज्मिक रेटिकुलम (रेटिकुलम) से रिक्तिकाएं बनती हैं
राइबोसोम।छोटे कण (17 - 23nm), जिसमें प्रोटीन और RNA की लगभग समान मात्रा होती है। राइबोसोम में, अमीनो एसिड मिलकर प्रोटीन बनाते हैं। सक्रिय चयापचय वाले कोशिकाओं में उनमें से अधिक हैं। राइबोसोम कोशिका के साइटोप्लाज्म में स्वतंत्र रूप से स्थित होते हैं या एंडोप्लाज्मिक रेटिकुलम (80S) से जुड़े होते हैं। वे नाभिक (80S), माइटोकॉन्ड्रिया (70S), प्लास्टिड्स (70S) में भी पाए जाते हैं।
राइबोसोम एक जटिल बना सकते हैं जिस पर समान पॉलीपेप्टाइड्स का एक साथ संश्लेषण होता है, जिसके बारे में जानकारी एक अणु और आरएनए से ली जाती है। इस तरह के एक जटिल को पॉलीराइबोसोम (पॉलीसोम) कहा जाता है। बड़ी मात्रा में प्रोटीन को संश्लेषित करने वाली कोशिकाओं में पॉलीसोम्स की एक व्यापक प्रणाली होती है, जो अक्सर परमाणु लिफाफे की बाहरी सतह से जुड़ी होती हैं।
अन्तः प्रदव्ययी जलिका।यह अनिश्चितकालीन सीमा की एक जटिल त्रि-आयामी झिल्ली प्रणाली है। ईआर के संदर्भ में उनके बीच एक संकीर्ण पारदर्शी स्थान के साथ दो प्राथमिक झिल्लियों की तरह दिखता है। ईआर का आकार और लंबाई सेल प्रकार, इसकी चयापचय गतिविधि और भेदभाव के चरण पर निर्भर करती है। कोशिकाओं में जो प्रोटीन को स्रावित या संग्रहीत करते हैं, ईआर को सपाट थैली या सिस्टर्न के आकार का होता है, जिसकी बाहरी सतह पर कई राइबोसोम जुड़े होते हैं। इस रेटिकुलम को रफ एंडोप्लाज्मिक रेटिकुलम कहा जाता है। एक चिकनी ईआर आमतौर पर आकार में ट्यूबलर होती है। खुरदरी और चिकनी अंतर्द्रव्यी जालिका एक ही कोशिका में मौजूद हो सकती है। एक नियम के रूप में, उनके बीच कई संबंध हैं।
एंडोप्लाज्मिक रेटिकुलम सेल की संचार प्रणाली के रूप में कार्य करता है। यह कर्नेल के बाहरी आवरण से जुड़ा होता है। वास्तव में, ये दो संरचनाएं एकल झिल्ली प्रणाली बनाती हैं। जब कोशिका विभाजन के दौरान परमाणु लिफाफा फट जाता है, तो इसके टुकड़े ER अंशों के समान हो जाते हैं। एंडोप्लाज्मिक रेटिकुलम पदार्थों के परिवहन के लिए एक प्रणाली है: प्रोटीन, लिपिड, कार्बोहाइड्रेट, कोशिका के विभिन्न भागों में। पड़ोसी कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम साइटोप्लाज्मिक स्ट्रैंड्स - प्लास्मोडेस्मेटा - से जुड़े होते हैं जो कोशिका झिल्ली से गुजरते हैं।
कोशिका झिल्लियों के संश्लेषण के लिए एंडोप्लाज्मिक रेटिकुलम मुख्य स्थल है। कुछ पादप कोशिकाओं में, रसधानियों और सूक्ष्म निकायों की झिल्लियों, तानाशाहों के गढ्ढों का निर्माण होता है।
गॉल्जीकाय।इस शब्द का प्रयोग एक सेल में सभी तानाशाही, या गोल्गी निकायों को संदर्भित करने के लिए किया जाता है। डिक्टियोसोम फ्लैट, डिस्क के आकार के पुटिकाओं या सिस्टर्न के समूह हैं, जो किनारों के साथ नलिकाओं की एक जटिल प्रणाली में शाखा करते हैं। उच्च पौधों में डिक्टायोसोम में एक साथ एकत्रित 4 - 8 कुंड होते हैं।
आमतौर पर, टैंकों के एक पैकेट में, एक विकासशील और परिपक्व पक्ष प्रतिष्ठित होता है। बनने वाले गढ्ढों की झिल्लियां संरचना में ईआर की झिल्लियों से मिलती-जुलती हैं, और परिपक्व हौदों की झिल्लियां प्लाज्मा झिल्ली से मिलती-जुलती हैं।
डिक्टायोसोम स्राव में शामिल होते हैं और अधिकांश उच्च पौधों में कोशिका झिल्लियों के निर्माण में शामिल होते हैं। डिक्टायोसोम द्वारा संश्लेषित कोशिका भित्ति पॉलीसेकेराइड पुटिकाओं में जमा हो जाते हैं, जो तब परिपक्व होने वाले सिस्टर्न से अलग हो जाते हैं। ये स्रावी पुटिकाएं सीएमपी के साथ माइग्रेट और फ्यूज हो जाती हैं; जबकि उनमें निहित पॉलीसेकेराइड कोशिका झिल्ली में निर्मित होते हैं। कुछ पदार्थ जो तानाशाहों में जमा होते हैं, अन्य संरचनाओं में बनते हैं, उदाहरण के लिए, ईआर में, और फिर तानाशाहों में ले जाया जाता है, जहां स्राव से पहले उन्हें संशोधित (संशोधित) किया जाता है। उदाहरण के लिए, ग्लाइकोप्रोटीन कोशिका झिल्ली की एक महत्वपूर्ण निर्माण सामग्री है। प्रोटीन भाग को किसी न किसी ईआर के पॉलीसोम द्वारा संश्लेषित किया जाता है, कार्बोहाइड्रेट भाग को डिक्टायोसोम में संश्लेषित किया जाता है, जहां दोनों भाग ग्लाइकोप्रोटीन बनाने के लिए संयोजित होते हैं।
झिल्लियां गतिशील, मोबाइल संरचनाएं हैं जो लगातार अपना आकार और क्षेत्र बदलती रहती हैं। एंडोप्लाज्मिक सिस्टम की अवधारणा झिल्ली की गतिशीलता पर आधारित है। इस अवधारणा के अनुसार, माइटोकॉन्ड्रिया और प्लास्टिड्स की झिल्लियों को छोड़कर, साइटोप्लाज्म की आंतरिक झिल्लियां एक ही पूरी होती हैं और एंडोप्लाज्मिक रेटिकुलम से उत्पन्न होती हैं। मध्यवर्ती पुटिका चरण के माध्यम से एंडोप्लाज्मिक रेटिकुलम से डिक्टायोसोम के नए कुंड बनते हैं, और स्रावी पुटिकाएं जो डिक्टीयोसोम से अलग होती हैं, अंततः प्लाज्मा झिल्ली के निर्माण में योगदान करती हैं। इस प्रकार, एंडोप्लाज्मिक रेटिकुलम और डिक्टियोसोम एक कार्यात्मक इकाई बनाते हैं जिसमें डिक्टायोसोम प्लाज्मा जैसी झिल्ली में एंडोप्लाज्मिक रेटिकुलम जैसी झिल्लियों को परिवर्तित करने की प्रक्रिया में मध्यवर्ती संरचनाओं की भूमिका निभाते हैं। उन ऊतकों में जिनकी कोशिकाएं बढ़ती हैं और कमजोर रूप से विभाजित होती हैं, झिल्ली के घटकों को लगातार अद्यतन किया जाता है।
सूक्ष्मनलिकाएंलगभग सभी यूकेरियोटिक कोशिकाओं में पाया जाता है। वे लगभग 24 एनएम के व्यास के साथ बेलनाकार संरचनाएं हैं। इनकी लंबाई अलग-अलग होती है। प्रत्येक नलिका ट्यूबुलिन नामक प्रोटीन की उपइकाइयों से बनी होती है। उपइकाइयां केंद्रीय गुहा के चारों ओर 13 अनुदैर्ध्य तंतु बनाती हैं। सूक्ष्मनलिकाएं गतिशील संरचनाएं हैं, वे नियमित रूप से नष्ट हो जाती हैं और कोशिका चक्र के कुछ चरणों में बनती हैं। इनका संयोजन विशेष स्थानों पर होता है, जिन्हें सूक्ष्मनलिकाय संगठन केंद्र कहते हैं। पादप कोशिकाओं में, उनके पास कमजोर व्यक्त अनाकार संरचना होती है।
सूक्ष्मनलिकाएं के कार्य: कोशिका झिल्ली के निर्माण में भाग लेते हैं; बनाने वाली झिल्ली के लिए तानाशाहों के सीधे बुलबुले, जैसे धुरी के धागे जो एक विभाजित कोशिका में बनते हैं; सेल प्लेट (बेटी कोशिकाओं के बीच प्रारंभिक सीमा) के निर्माण में भूमिका निभाते हैं। इसके अलावा, सूक्ष्मनलिकाएं फ्लैगेल्ला और सिलिया का एक महत्वपूर्ण घटक हैं, जिसके संचलन में वे महत्वपूर्ण भूमिका निभाते हैं।
सूक्ष्म तंतु,सूक्ष्मनलिकाएं की तरह, वे लगभग सभी यूकेरियोटिक कोशिकाओं में पाए जाते हैं। वे 5-7 एनएम मोटे लंबे तंतु होते हैं, जिनमें सिकुड़ा हुआ प्रोटीन एक्टिन होता है। उच्च पौधों की कई कोशिकाओं में माइक्रोफ़िल्मेंट्स के बंडल पाए जाते हैं। जाहिर है, वे साइटोप्लाज्म की धाराओं में एक महत्वपूर्ण भूमिका निभाते हैं। माइक्रोफ़िल्मेंट्स, सूक्ष्मनलिकाएं के साथ मिलकर एक लचीला नेटवर्क बनाते हैं जिसे साइटोस्केलेटन कहा जाता है।
आधार पदार्थलंबे समय तक एक सजातीय (सजातीय) प्रोटीन युक्त समाधान माना जाता था जिसमें कम संख्या में संरचनाएं या यहां तक कि संरचना रहित होती थी। हालांकि, वर्तमान में, एक उच्च-वोल्टेज इलेक्ट्रॉन माइक्रोस्कोप का उपयोग करके, यह स्थापित किया गया है कि जमीनी पदार्थ एक त्रि-आयामी जाली है जो पतली (3–6 एनएम व्यास) किस्में से निर्मित होती है जो पूरे सेल को भरती है। सूक्ष्मनलिकाएं और माइक्रोफ़िल्मेंट्स सहित साइटोप्लाज्म के अन्य घटक इस माइक्रोट्रैब्युलर जाली से निलंबित हैं।
माइक्रोट्राबेकुलर संरचना प्रोटीन स्ट्रैंड्स की एक जाली है, जिसके बीच का स्थान पानी से भरा होता है। पानी के साथ, जाली में जेल की स्थिरता होती है; जेल जिलेटिनस निकायों की तरह दिखता है।
ऑर्गेनेल माइक्रोट्राबेकुलर जाली से जुड़े होते हैं। जाली कोशिका के अलग-अलग हिस्सों के बीच संचार प्रदान करती है और इंट्रासेल्युलर परिवहन को निर्देशित करती है।
लिपिड बूँदें- एक गोलाकार आकार की संरचनाएं, एक प्रकाश सूक्ष्मदर्शी के तहत एक पादप कोशिका के साइटोप्लाज्म को ग्रैन्युलैरिटी देती हैं। इलेक्ट्रॉन माइक्रोग्राफ पर, वे अनाकार दिखते हैं। प्लास्टिड्स में बहुत समान लेकिन छोटी बूंदें पाई जाती हैं।
लिपिड बूँदें, गलती से उन्हें ऑर्गेनेल समझकर, उन्हें स्फेरोसोम्स कहते हैं और मानते हैं कि वे एक या दो परत वाली झिल्ली से घिरे हुए हैं। हालाँकि, हाल के आंकड़े बताते हैं कि लिपिड बूंदों में झिल्ली नहीं होती है, लेकिन प्रोटीन के साथ लेपित हो सकते हैं।
एर्गास्टिक पदार्थ -ये प्रोटोप्लास्ट के "निष्क्रिय उत्पाद" हैं: आरक्षित पदार्थ या अपशिष्ट उत्पाद। वे कोशिका चक्र की विभिन्न अवधियों में प्रकट और गायब हो सकते हैं। स्टार्च अनाज, क्रिस्टल, एंथोसायनिन पिगमेंट और लिपिड बूंदों को छोड़कर। इनमें रेजिन, गोंद, टैनिन और प्रोटीन शामिल हैं। एर्गैस्टिक पदार्थ कोशिका झिल्ली का हिस्सा होते हैं, साइटोप्लाज्म और ऑर्गेनेल का मुख्य पदार्थ, जिसमें रिक्तिकाएं भी शामिल हैं।
फ्लैगेल्ला और सिलिया -वे पतली, बालों जैसी संरचनाएं हैं जो कई यूकेरियोटिक कोशिकाओं की सतह से फैली हुई हैं। उनका एक स्थिर व्यास है, लेकिन लंबाई 2 से 150 माइक्रोन तक भिन्न होती है। परंपरागत रूप से, उनमें से लंबे और कुछ को फ्लैगेल्ला कहा जाता है, और छोटे और अधिक कई को सिलिया कहा जाता है। इन दो प्रकार की संरचनाओं के बीच कोई स्पष्ट अंतर नहीं है, इसलिए फ्लैगेलम शब्द का उपयोग दोनों को संदर्भित करने के लिए किया जाता है।
कुछ शैवालों तथा कवकों में कशाभिका (Flagella) चलन अंग होते हैं, जिनकी सहायता से ये जल में गति करते हैं। पौधों में (उदाहरण के लिए, काई, लिवरवॉर्ट्स, फ़र्न, कुछ जिम्नोस्पर्म), केवल जर्म कोशिकाओं (युग्मक) में फ्लैगेल्ला होता है।
प्रत्येक फ्लैगेलम का एक विशिष्ट संगठन होता है। सूक्ष्मनलिकाएं के 9 जोड़े की एक बाहरी रिंग फ्लैगेलम के केंद्र में स्थित दो अतिरिक्त सूक्ष्मनलिकाएं को घेरे हुए है। एंजाइम युक्त "हैंडल" प्रत्येक बाहरी जोड़े के एक सूक्ष्मनलिका से फैलता है। यह यूकेरियोटिक जीवों में सभी कशाभों में पाया जाने वाला बुनियादी 9+2 संगठन पैटर्न है। यह माना जाता है कि फ्लैगेल्ला की गति सूक्ष्मनलिकाएं के फिसलने पर आधारित होती है, जबकि सूक्ष्मनलिकाएं के बाहरी जोड़े बिना संकुचन के एक दूसरे के साथ चलते हैं। एक दूसरे के सापेक्ष सूक्ष्मनलिकाएं के जोड़े के फिसलने से फ्लैगेलम का स्थानीय झुकाव होता है।
फ्लैगेल्ला साइटोप्लाज्मिक बेलनाकार संरचनाओं से "बढ़ता है" जिसे बेसल बॉडी कहा जाता है, जो फ्लैगेलम का बेसल भाग भी बनाता है। बेसल निकायों में एक फ्लैगेलम जैसी आंतरिक संरचना होती है, सिवाय इसके कि बाहरी नलिकाओं को जोड़े के बजाय ट्रिपल में व्यवस्थित किया जाता है, और केंद्रीय नलिकाएं अनुपस्थित होती हैं।
कोशिका भित्ति।कोशिका भित्ति प्रोटोप्लास्ट के आकार को परिसीमित करती है और रिक्तिका द्वारा पानी के अवशोषण के कारण इसके टूटने को रोकती है।
कोशिका भित्ति में विशिष्ट कार्य होते हैं जो न केवल कोशिका और उस ऊतक के लिए महत्वपूर्ण होते हैं जिसमें कोशिका स्थित होती है, बल्कि पूरे पौधे के लिए भी महत्वपूर्ण होती है। कोशिका भित्ति पदार्थों के अवशोषण, परिवहन और उत्सर्जन में एक आवश्यक भूमिका निभाती है, और इसके अलावा, लाइसोसोमल या पाचन गतिविधि उनमें केंद्रित हो सकती है।
सेल की दीवार के घटक। कोशिका भित्ति का सबसे विशिष्ट घटक सेल्युलोज है, जो काफी हद तक इसकी वास्तुकला को निर्धारित करता है। सेल्युलोज अणु अंत से अंत तक जुड़े ग्लूकोज अणुओं को दोहराते हुए बने होते हैं। लंबे पतले सेल्युलोज अणुओं को 10 - 25 एनएम की मोटाई के साथ माइक्रोफाइब्रिल में जोड़ा जाता है। माइक्रोफ़ाइब्रिल्स आपस में जुड़ते हैं और पतले धागे बनाते हैं, जो बदले में एक दूसरे के चारों ओर लपेटे जा सकते हैं, जैसे कि रस्सी में किस्में। ऐसी प्रत्येक "रस्सी", या मैक्रोफिब्रिल की मोटाई लगभग 0.5 माइक्रोन होती है, जो 4 माइक्रोन की लंबाई तक पहुंचती है। मैक्रोफाइब्रिल समान आकार के स्टील के तार जितने मजबूत होते हैं।
सेल की दीवार का सेल्यूलोज फ्रेम इसके साथ जुड़े सेल्यूलोज मैट्रिक्स अणुओं से भरा होता है। इसमें पॉलीसेकेराइड होते हैं जिन्हें हेमिकेलुलोज कहा जाता है, और पेक्टिन, या पेक्टिन, रासायनिक रूप से हेमिकेलुलोज के बहुत करीब होते हैं।
कोशिका भित्ति का एक अन्य घटक, लिग्निन, सेल्युलोज के बाद पादप कोशिकाओं में सबसे प्रचुर मात्रा में पाया जाने वाला बहुलक है। लिग्निन दीवार की कठोरता को बढ़ाता है और आमतौर पर उन कोशिकाओं में पाया जाता है जो एक सहायक या यांत्रिक कार्य करती हैं।
क्यूटिन, सुबेरिन, मोम - आमतौर पर पौधों के सुरक्षात्मक ऊतकों के गोले में जमा होते हैं। क्यूटिन, उदाहरण के लिए, एपिडर्मिस की कोशिका झिल्लियों में पाया जाता है, और सुबेरिन द्वितीयक सुरक्षात्मक ऊतक, कॉर्क में पाया जाता है। दोनों पदार्थ वैक्स के संयोजन में पाए जाते हैं और पौधे से अत्यधिक पानी के नुकसान को रोकते हैं।
कोशिका भित्ति की परतें। पादप कोशिकाओं की दीवार की मोटाई व्यापक रूप से भिन्न होती है, जो पादप संरचना में कोशिकाओं की भूमिका और स्वयं कोशिका की आयु पर निर्भर करती है। एक इलेक्ट्रॉन माइक्रोस्कोप के तहत, पौधे की कोशिका दीवार में दो परतें दिखाई देती हैं: माध्यिका पटल (जिसे अंतरकोशिकीय पदार्थ भी कहा जाता है), और प्राथमिक कोशिका भित्ति। कई कोशिकाएं एक और परत बिछाती हैं, द्वितीयक कोशिका भित्ति। मीडियन प्लेट आसन्न कोशिकाओं की प्राथमिक दीवारों के बीच स्थित होती है। द्वितीयक दीवार, यदि कोई हो, प्राथमिक सेल दीवार की आंतरिक सतह पर सेल प्रोटोप्लास्ट द्वारा जमा की जाती है।
बीच की थाली। मीडियन लैमिना में मुख्य रूप से पेक्टिन होता है। जहां एक कोशिका भित्ति उत्पन्न होनी चाहिए, दो नवगठित कोशिकाओं के बीच, एंडोप्लाज़मिक रेटिकुलम के नलिकाओं का एक घना जाल और गोल्गी तंत्र (डिक्टायोसोम) का कुंड पहले नोट किया जाता है। फिर इस जगह में पेक्टिन (पॉलीसेकेराइड से) से भरे बुलबुले दिखाई देते हैं। इन बुलबुलों को गॉल्जी उपकरण की टंकियों से अलग किया जाता है। प्रारंभिक कोशिका भित्ति में विभिन्न पॉलीसेकेराइड होते हैं, जिनमें से मुख्य हैं पेक्टिन और हेमिकेलुलोज। बाद में, इसमें सघन पदार्थ - सेल्युलोज और लिग्निन होते हैं।
प्राथमिक कोशिका भित्ति। यह सेल्युलोज आवरण की एक परत है जो कोशिका वृद्धि से पहले या उसके दौरान जमा होती है। सेलूलोज़, हेमिसेल्यूलोज़ और पेक्टिन के अलावा, प्राथमिक झिल्ली में ग्लाइकोप्रोटीन होता है। प्राथमिक गोले लिग्निफाई कर सकते हैं। पेक्टिन घटक नमनीयता प्रदान करता है, जो प्राथमिक खोल को जड़, तना या पत्ती के रूप में विस्तारित करने की अनुमति देता है।
सक्रिय रूप से विभाजित होने वाली कोशिकाओं (प्रकाश संश्लेषण, श्वसन और स्राव की प्रक्रियाओं में शामिल अधिकांश परिपक्व कोशिकाओं) में प्राथमिक झिल्ली होती है। एक प्राथमिक झिल्ली और एक जीवित प्रोटोप्लास्ट वाली ऐसी कोशिकाएं अपने विशिष्ट आकार को खोने, विभाजित करने और एक नए प्रकार की कोशिका में अंतर करने में सक्षम होती हैं। वे पौधों में घाव भरने और ऊतक पुनर्जनन में शामिल हैं।
प्राथमिक कोशिका झिल्लियां अपनी पूरी लंबाई में मोटाई में एक समान नहीं होती हैं, लेकिन पतले खंड होते हैं, जिन्हें प्राथमिक छिद्र क्षेत्र कहा जाता है। साइटोप्लाज्म, या प्लास्मोडेस्माटा की किस्में, जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ती हैं, आमतौर पर प्राथमिक छिद्र क्षेत्रों से गुजरती हैं।
द्वितीयक कोशिका भित्ति। इस तथ्य के बावजूद कि कई पादप कोशिकाओं में केवल एक प्राथमिक दीवार होती है, कुछ में, प्रोटोप्लास्ट कोशिका के केंद्र की ओर एक द्वितीयक दीवार जमा करता है। यह आमतौर पर कोशिका वृद्धि की समाप्ति के बाद होता है और प्राथमिक झिल्ली का क्षेत्र अब नहीं बढ़ता है। इस कारण से, द्वितीयक खोल प्राथमिक से भिन्न होता है। माध्यमिक झिल्लियों की विशेष रूप से विशेष कोशिकाओं द्वारा आवश्यकता होती है जो पौधे को मजबूत करती हैं और पानी का संचालन करती हैं। द्वितीयक झिल्ली के निक्षेपण के बाद, इन कोशिकाओं के प्रोटोप्लास्ट, एक नियम के रूप में, मर जाते हैं। माध्यमिक गोले में प्राथमिक की तुलना में अधिक सेल्यूलोज होता है, और उनमें पेक्टिन पदार्थ और ग्लाइकोप्रोटीन अनुपस्थित होते हैं। द्वितीयक झिल्ली को फैलाना मुश्किल होता है, इसके मैट्रिक्स में हेमिकेलुलोज होता है।
द्वितीयक खोल में, तीन परतों को प्रतिष्ठित किया जा सकता है - बाहरी, मध्य और आंतरिक (S1, S2, S3)। द्वितीयक गोले की स्तरित संरचना उनकी ताकत में काफी वृद्धि करती है। द्वितीयक झिल्ली में सेल्युलोज माइक्रोफाइब्रिल्स प्राथमिक की तुलना में सघन रूप से जमा होते हैं। लिग्निन लकड़ी की द्वितीयक खाल का एक सामान्य घटक है।
संपर्क कोशिकाओं की झिल्लियों में छिद्र एक दूसरे के विपरीत स्थित होते हैं। दो विरोधी छिद्र और एक छिद्र झिल्ली छिद्रों की एक जोड़ी बनाते हैं। द्वितीयक झिल्लियों वाली कोशिकाओं में, दो मुख्य प्रकार के छिद्र होते हैं: सरल और सीमाबद्ध। बॉर्डर वाले छिद्रों में, द्वितीयक दीवार छिद्र गुहा के ऊपर लटकी होती है। सिंपल पोर्स में ऐसा नहीं होता है।
कोशिका भित्ति का विकास। जैसे-जैसे कोशिका बढ़ती है, कोशिका झिल्ली की मोटाई और क्षेत्रफल बढ़ता जाता है। शेल स्ट्रेचिंग एक जटिल प्रक्रिया है। यह प्रोटोप्लास्ट द्वारा नियंत्रित होता है और हार्मोन ऑक्सिन द्वारा नियंत्रित होता है।
सभी दिशाओं में समान रूप से बढ़ने वाली कोशिकाओं में, मायोफिब्रिल्स का जमाव यादृच्छिक होता है। ये मायोफिब्रिल एक अनियमित नेटवर्क बनाते हैं। ऐसी कोशिकाएं तने के मूल भाग, भंडारण के ऊतकों और इन विट्रो में कोशिका संवर्धन के दौरान पाई गईं। लम्बी कोशिकाओं में, पार्श्व आवरण के पेशीतंतुओं को बढ़ाव के अक्ष पर समकोण पर जमा किया जाता है।
मैट्रिक्स पदार्थ - पेक्टिन, हेमिकेलुलोज और ग्लाइकोप्रोटीन को डिक्टीयोसोम के पुटिकाओं में झिल्ली में स्थानांतरित किया जाता है। इसी समय, पेक्टिन बढ़ती कोशिकाओं की अधिक विशेषता है, जबकि हेमिकेलुलोज गैर-बढ़ती कोशिकाओं में प्रबल होते हैं।
सेल्युलोज माइक्रोफिब्रिल को कोशिका की सतह पर प्लाज्मा झिल्ली से जुड़े एक एंजाइम कॉम्प्लेक्स की मदद से संश्लेषित किया जाता है। सूक्ष्मतंतुओं का उन्मुखीकरण प्लाज्मा झिल्ली की आंतरिक सतह के पास स्थित सूक्ष्मनलिकाएं द्वारा नियंत्रित होता है।
प्लाज्मोडेस्मा।ये साइटोप्लाज्म के पतले धागे हैं जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ते हैं। प्लास्मोडेस्माटा या तो कहीं भी कोशिका भित्ति से होकर गुजरता है, या प्राथमिक छिद्र क्षेत्रों में या छिद्रों के जोड़े के बीच झिल्लियों में केंद्रित होता है। एक इलेक्ट्रॉन माइक्रोस्कोप के तहत, प्लास्मोडेमाटा प्लाज्मा झिल्ली के साथ पंक्तिबद्ध संकीर्ण चैनल के रूप में दिखाई देते हैं। नहर की धुरी के साथ, एक छोटी बेलनाकार ट्यूब, डेस्मोटुबुल, एक कोशिका से दूसरी कोशिका तक फैली हुई है, जो दोनों आसन्न कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम के साथ संचार करती है। कोशिका विभाजन के दौरान कई प्लास्मोडेस्माटा बनते हैं, जब ट्यूबलर एंडोप्लाज्मिक रेटिकुलम विकासशील सेल प्लेट द्वारा लिया जाता है। प्लास्मोडेस्माटा अविभाजित कोशिकाओं की झिल्लियों में भी बन सकता है। ये संरचनाएं कोशिका से कोशिका में कुछ पदार्थों का कुशल स्थानांतरण प्रदान करती हैं।
कोशिका विभाजन।बहुकोशिकीय जीवों में, कोशिका विभाजन, उनके आकार में वृद्धि के साथ, पूरे जीव के विकास का एक तरीका है। विभाजन के दौरान बनने वाली नई कोशिकाएं मूल कोशिका और एक दूसरे दोनों की संरचना और कार्य में समान होती हैं। यूकेरियोट्स में विभाजन प्रक्रिया को दो आंशिक रूप से अतिव्यापी चरणों में विभाजित किया जा सकता है: माइटोसिस और साइटोकाइनेसिस।
माइटोसिस एक नाभिक से दो बेटी नाभिक का निर्माण होता है, एक दूसरे के रूपात्मक और आनुवंशिक रूप से समतुल्य। साइटोकिनेसिस बेटी कोशिकाओं के गठन के साथ कोशिका के साइटोप्लाज्मिक भाग का विभाजन है।
कोशिका चक्र।एक जीवित कोशिका क्रमिक घटनाओं की एक श्रृंखला से गुजरती है जो कोशिका चक्र बनाती है। चक्र की अवधि स्वयं कोशिका के प्रकार और बाहरी कारकों जैसे तापमान या पोषक तत्वों की उपलब्धता के आधार पर भिन्न होती है। आमतौर पर चक्र को इंटरपेज़ और माइटोसिस के चार चरणों में विभाजित किया जाता है।
अंतरावस्था।क्रमिक माइटोटिक विभाजनों के बीच की अवधि।
इंटरपेज़ को तीन अवधियों में विभाजित किया गया है, जिसे G1, S, G2 के रूप में दर्शाया गया है।
G1 अवधि के दौरान, जो माइटोसिस के बाद शुरू होती है। इस अवधि के दौरान, विभिन्न जीवों सहित साइटोप्लाज्म की मात्रा बढ़ जाती है। इसके अलावा, वर्तमान परिकल्पना के अनुसार, पदार्थ G1 अवधि के दौरान संश्लेषित होते हैं जो या तो S अवधि और शेष चक्र को उत्तेजित या बाधित करते हैं, इस प्रकार विभाजन प्रक्रिया का निर्धारण करते हैं।
S अवधि G1 अवधि का अनुसरण करती है, जिस समय आनुवंशिक सामग्री (डीएनए) का दोहराव होता है।
G2 अवधि के दौरान, जो S का अनुसरण करता है, संरचनाएं जो सीधे माइटोसिस में शामिल होती हैं, जैसे कि धुरी घटक, बनते हैं।
कुछ कोशिकाएं असीमित संख्या में सेल चक्रों से गुजरती हैं। ये एककोशिकीय जीव और सक्रिय विकास क्षेत्रों (मेरिस्टेम) की कुछ कोशिकाएँ हैं। परिपक्वता के बाद कुछ विशिष्ट कोशिकाएं प्रजनन करने की अपनी क्षमता खो देती हैं। कोशिकाओं का तीसरा समूह, उदाहरण के लिए, घाव के ऊतक (कैलस) बनाने वाले, केवल विशेष परिस्थितियों में विभाजित करने की क्षमता को बरकरार रखते हैं।
सूत्रीविभाजन,या परमाणु विखंडन। यह एक सतत प्रक्रिया है जिसे चार चरणों में बांटा गया है: प्रोफ़ेज़, मेटाफ़ेज़, एनाफ़ेज़, टेलोफ़ेज़। माइटोसिस के परिणामस्वरूप, आनुवंशिक सामग्री जो इंटरपेज़ में दोगुनी हो गई है, दो बेटी नाभिकों के बीच समान रूप से विभाजित हो जाती है।
विभाजन के लिए एक कोशिका के संक्रमण के शुरुआती संकेतों में से एक प्लाज्मा झिल्ली के ठीक नीचे सूक्ष्मनलिकाएं के एक संकीर्ण, कुंडलाकार बैंड की उपस्थिति है। यह अपेक्षाकृत सघन बेल्ट भविष्य के माइटोटिक स्पिंडल के विषुवतीय तल में नाभिक को घेरता है। चूंकि यह प्रोफ़ेज़ से पहले प्रकट होता है, इसलिए इसे प्रीप्रोफ़ेज़ गर्डल कहा जाता है। यह माइटोटिक स्पिंडल के बाद गायब हो जाता है, सेल प्लेट के लेट टेलोफ़ेज़ में प्रकट होने से बहुत पहले, जो केंद्र से परिधि तक बढ़ता है और प्रीप्रोफ़ेज़ बेल्ट द्वारा कब्जा किए गए क्षेत्र में मातृ कोशिका झिल्ली के साथ विलीन हो जाता है।
प्रोफ़ेज़। प्रोफ़ेज़ की शुरुआत में, गुणसूत्र नाभिक के अंदर बिखरे हुए लंबे धागों के समान होते हैं। फिर, जैसे-जैसे तंतु छोटे और मोटे होते जाते हैं, कोई यह देख सकता है कि प्रत्येक गुणसूत्र एक नहीं, बल्कि दो गुंथे हुए तंतुओं से बना होता है जिन्हें क्रोमैटिड कहा जाता है। देर से प्रोफ़ेज़ में, प्रत्येक गुणसूत्र के दो छोटे युग्मित क्रोमैटिड समानांतर में एक साथ होते हैं, जो सेंट्रोमियर नामक एक संकीर्ण खंड से जुड़े होते हैं। यह प्रत्येक गुणसूत्र पर एक विशिष्ट स्थिति रखता है और गुणसूत्र को अलग-अलग लंबाई की दो भुजाओं में विभाजित करता है।
सूक्ष्मनलिकाएं धुरी की धुरी के साथ नाभिक की सतह के समानांतर स्थित होती हैं। यह माइटोटिक स्पिंडल असेंबली की सबसे पहली अभिव्यक्ति है।
प्रोफ़ेज़ के अंत तक, नाभिक धीरे-धीरे अपनी स्पष्ट रूपरेखा खो देता है और अंत में गायब हो जाता है। इसके तुरंत बाद, परमाणु लिफाफा भी बिखर जाता है।
रूपक। मेटाफ़ेज़ की शुरुआत में, स्पिंडल, जो एक त्रि-आयामी संरचना है, मध्य में सबसे चौड़ा और ध्रुवों की ओर पतला होता है, पहले नाभिक द्वारा कब्जा कर लिया गया स्थान लेता है। स्पिंडल फाइबर सूक्ष्मनलिकाएं के बंडल हैं। मेटाफ़ेज़ के दौरान, क्रोमोसोम, जिनमें से प्रत्येक में दो क्रोमैटिड होते हैं, को व्यवस्थित किया जाता है ताकि उनके सेंट्रोमर्स स्पिंडल के इक्वेटोरियल प्लेन में स्थित हों। इसके सेंट्रोमियर के साथ, प्रत्येक गुणसूत्र स्पिंडल थ्रेड्स से जुड़ा होता है। हालाँकि, कुछ किस्में गुणसूत्रों से जुड़े बिना एक ध्रुव से दूसरे ध्रुव तक जाती हैं।
जब सभी गुणसूत्र विषुवतीय तल में स्थित होते हैं, तो मेटाफ़ेज़ पूरा हो जाता है। गुणसूत्र विभाजित होने के लिए तैयार होते हैं।
पश्चावस्था। प्रत्येक गुणसूत्र के क्रोमैटिड अलग हो जाते हैं। अब ये पुत्री गुणसूत्र हैं। सबसे पहले, सेंट्रोमियर विभाजित होता है, और दो बेटी गुणसूत्र विपरीत ध्रुवों पर खींचे जाते हैं। इस मामले में, सेंट्रोमर्स आगे बढ़ते हैं, और गुणसूत्रों की भुजाएँ पीछे की ओर खिंचती हैं। क्रोमोसोम से जुड़े स्पिंडल फिलामेंट्स छोटे हो जाते हैं, क्रोमैटिड्स को अलग करने और विपरीत दिशाओं में बेटी क्रोमोसोम के आंदोलन में योगदान करते हैं।
प्लांट सेल की संरचना
एक पादप कोशिका में कम या ज्यादा कठोर कोशिका भित्ति और एक प्रोटोप्लास्ट होता है। कोशिका भित्ति कोशिका भित्ति और साइटोप्लाज्मिक झिल्ली है। प्रोटोप्लास्ट शब्द प्रोटोप्लाज्म शब्द से आया है, जिसका उपयोग लंबे समय से सभी जीवित चीजों को संदर्भित करने के लिए किया जाता है। प्रोटोप्लास्ट एक व्यक्तिगत कोशिका का प्रोटोप्लाज्म है।
प्रोटोप्लास्ट में साइटोप्लाज्म और न्यूक्लियस होते हैं। साइटोप्लाज्म में ऑर्गेनेल (राइबोसोम, माइक्रोट्यूबुल्स, प्लास्टिड्स, माइटोकॉन्ड्रिया) और मेम्ब्रेन सिस्टम (एंडोप्लाज्मिक रेटिकुलम, डिक्टीयोसोम) होते हैं। साइटोप्लाज्म में साइटोप्लाज्मिक मैट्रिक्स (मूल पदार्थ) भी शामिल होता है जिसमें ऑर्गेनेल और मेम्ब्रेन सिस्टम डूबे रहते हैं। साइटोप्लाज्म को कोशिका भित्ति से प्लाज्मा झिल्ली द्वारा अलग किया जाता है, जो एक प्राथमिक झिल्ली है। अधिकांश पशु कोशिकाओं के विपरीत, पादप कोशिकाओं में एक या एक से अधिक रिक्तिकाएँ होती हैं। ये पुटिकाएं तरल से भरी होती हैं और एक प्रारंभिक झिल्ली (टोनोप्लास्ट) से घिरी होती हैं।
एक जीवित पादप कोशिका में, जमीनी पदार्थ निरंतर गति में रहता है। ऑर्गेनेल आंदोलन में शामिल होते हैं, जिसे साइटोप्लाज्म या साइक्लोसिस का प्रवाह कहा जाता है। साइक्लोसिस कोशिका में पदार्थों की आवाजाही और कोशिका और पर्यावरण के बीच उनके आदान-प्रदान की सुविधा प्रदान करता है।
प्लाज्मा झिल्ली।यह एक बाइलेयर फॉस्फोलिपिड संरचना है। पादप कोशिकाओं की विशेषता प्लाज्मा झिल्ली के अंतर्वलन से होती है।
प्लाज्मा झिल्ली निम्नलिखित कार्य करती है:
कोशिका और पर्यावरण के बीच पदार्थों के आदान-प्रदान में भाग लेता है;
कोशिका भित्ति के सेल्युलोज सूक्ष्मतंतुओं के संश्लेषण और संयोजन का समन्वय करता है;
हार्मोनल और बाहरी संकेतों को प्रसारित करता है जो सेल के विकास और भेदभाव को नियंत्रित करते हैं।
मुख्य।यह यूकेरियोटिक कोशिका के साइटोप्लाज्म में सबसे प्रमुख संरचना है। कर्नेल दो महत्वपूर्ण कार्य करता है:
कोशिका की महत्वपूर्ण गतिविधि को नियंत्रित करता है, यह निर्धारित करता है कि कौन से प्रोटीन और किस समय संश्लेषित किए जाने चाहिए;
आनुवंशिक जानकारी को संग्रहीत करता है और इसे कोशिका विभाजन के दौरान संतति कोशिकाओं तक पहुंचाता है।
एक यूकेरियोटिक कोशिका का केंद्रक दो प्राथमिक झिल्लियों से घिरा होता है जो परमाणु आवरण बनाती हैं। यह 30 से 100 एनएम के व्यास के साथ कई छिद्रों से व्याप्त है, जो केवल एक इलेक्ट्रॉन माइक्रोस्कोप में दिखाई देता है। छिद्रों की एक जटिल संरचना होती है। कुछ स्थानों पर परमाणु झिल्ली की बाहरी झिल्ली को एंडोप्लाज्मिक रेटिकुलम के साथ जोड़ा जाता है। परमाणु लिफाफे को एंडोप्लाज्मिक रेटिकुलम (ईआर) के एक विशेष, स्थानीय रूप से विभेदित भाग के रूप में देखा जा सकता है।
विशेष रंगों से सना हुआ नाभिक में, क्रोमैटिन और न्यूक्लियोप्लाज्म (नाभिक का मुख्य पदार्थ) के पतले तंतुओं और गुच्छों को प्रतिष्ठित किया जा सकता है। क्रोमेटिन हिस्टोन नामक विशेष प्रोटीन से बंधे डीएनए से बना होता है। कोशिका विभाजन की प्रक्रिया में, क्रोमैटिन अधिक से अधिक संकुचित हो जाता है और गुणसूत्रों में इकट्ठा हो जाता है। डीएनए आनुवंशिक जानकारी को एनकोड करता है।
दैहिक कोशिकाओं में जीव गुणसूत्रों की संख्या में भिन्न होते हैं। उदाहरण के लिए, गोभी में - 20 गुणसूत्र होते हैं; सूरजमुखी - 34; गेहूँ - 42; मनुष्य - 46, और फ़र्न ओफ़ियोग्लोसम की प्रजातियों में से एक - 1250। सेक्स कोशिकाओं (युग्मकों) में शरीर की दैहिक कोशिकाओं के गुणसूत्रों की संख्या का केवल आधा हिस्सा होता है। युग्मकों में गुणसूत्रों की संख्या को अगुणित (एकल) कहा जाता है, दैहिक कोशिकाओं में - द्विगुणित (डबल)। जिन कोशिकाओं में गुणसूत्रों के दो से अधिक सेट होते हैं उन्हें पॉलीप्लोइड कहा जाता है।
एक प्रकाश सूक्ष्मदर्शी के तहत, गोलाकार संरचनाएं - न्यूक्लियोली - देखी जा सकती हैं। प्रत्येक नाभिक में एक या एक से अधिक नाभिक होते हैं, जो अविभाजित नाभिक में दिखाई देते हैं। राइबोसोमल आरएनए को न्यूक्लियोलस में संश्लेषित किया जाता है। आमतौर पर, द्विगुणित जीवों के नाभिक में दो नाभिक होते हैं, गुणसूत्रों के प्रत्येक अगुणित सेट के लिए एक। न्यूक्लियोली की अपनी झिल्ली नहीं होती है। बायोकेमिकल रूप से, न्यूक्लियोली को आरएनए की उच्च सांद्रता की विशेषता होती है, जो यहां फॉस्फोप्रोटीन के साथ जुड़ा हुआ है। नाभिक का आकार कोशिका की कार्यात्मक अवस्था पर निर्भर करता है। यह देखा गया है कि एक तेजी से बढ़ने वाली कोशिका में, जिसमें प्रोटीन संश्लेषण की गहन प्रक्रियाएँ हो रही होती हैं, नाभिक का आकार बढ़ जाता है। न्यूक्लियोली में, एमआरएनए और राइबोसोम उत्पन्न होते हैं, जो केवल न्यूक्लियस में सिंथेटिक कार्य करते हैं।
न्यूक्लियोप्लाज्म (कार्योप्लाज्म) एक सजातीय तरल द्वारा दर्शाया जाता है जिसमें एंजाइम सहित विभिन्न प्रोटीन घुल जाते हैं।
प्लास्टिड्स।रिक्तिकाएं, सेल्युलोज कोशिका भित्ति और प्लास्टिड पादप कोशिकाओं के विशिष्ट घटक हैं। प्रत्येक प्लास्टिड का अपना खोल होता है, जिसमें दो प्राथमिक झिल्लियाँ होती हैं। प्लास्टिड के अंदर, एक झिल्ली प्रणाली और एक सजातीय पदार्थ की अलग-अलग डिग्री, स्ट्रोमा, प्रतिष्ठित हैं। परिपक्व प्लास्टिड्स को उनमें मौजूद पिगमेंट के आधार पर वर्गीकृत किया जाता है।
क्लोरोप्लास्ट, जिसमें प्रकाश संश्लेषण होता है, में क्लोरोफिल और कैरोटीनॉयड होते हैं। आमतौर पर 4 - 5 माइक्रोन के व्यास वाली डिस्क का रूप होता है। मेसोफिल (पत्ती के मध्य) की एक कोशिका में 40-50 क्लोरोप्लास्ट हो सकते हैं; पत्ती के मिमी 2 में - लगभग 500,000। साइटोप्लाज्म में, क्लोरोप्लास्ट आमतौर पर कोशिका झिल्ली के समानांतर स्थित होते हैं।
क्लोरोप्लास्ट की आंतरिक संरचना जटिल है। स्ट्रोमा को पुटिकाओं - थायलाकोइड्स के रूप में झिल्लियों की एक विकसित प्रणाली द्वारा अनुमति दी जाती है। प्रत्येक थायलाकोइड में दो झिल्ली होते हैं। थायलाकोइड्स एकल प्रणाली बनाते हैं। एक नियम के रूप में, वे ढेर में एकत्र किए जाते हैं - अनाज, सिक्कों के स्तंभों जैसा। अलग-अलग ग्रैने के थायलाकोइड्स स्ट्रोमा थायलाकोइड्स, या इंटरग्रेनल थायलाकोइड्स द्वारा परस्पर जुड़े होते हैं। क्लोरोफिल और कैरोटीनॉयड थायलाकोइड झिल्ली में एम्बेडेड होते हैं। हरे पौधों और शैवाल में क्लोरोप्लास्ट में अक्सर स्टार्च के दाने और छोटे लिपिड (वसा) की बूंदें होती हैं। स्टार्च के दाने प्रकाश संश्लेषण उत्पादों के अस्थायी भंडार होते हैं। वे केवल 24 घंटों के लिए अंधेरे में रहने वाले पौधे के क्लोरोप्लास्ट से गायब हो सकते हैं, और पौधे को प्रकाश में स्थानांतरित करने के 3-4 घंटे बाद फिर से प्रकट हो सकते हैं।
क्लोरोप्लास्ट अर्ध-स्वायत्त अंगक होते हैं और बैक्टीरिया के समान होते हैं। उदाहरण के लिए, बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम में काफी उच्च समानता होती है। वे यूकेरियोटिक राइबोसोम से छोटे होते हैं। बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम पर प्रोटीन संश्लेषण क्लोरैम्फेनिकॉल द्वारा बाधित होता है, जिसका यूकेरियोटिक कोशिकाओं पर कोई प्रभाव नहीं पड़ता है। इसके अलावा, बैक्टीरिया और क्लोरोप्लास्ट दोनों में एक समान प्रकार का न्यूक्लियॉइड होता है, जो समान तरीके से व्यवस्थित होता है। इस तथ्य के बावजूद कि क्लोरोप्लास्ट का निर्माण और उनमें निहित पिगमेंट का संश्लेषण काफी हद तक कोशिका के क्रोमोसोमल डीएनए द्वारा नियंत्रित होता है, फिर भी, अपने स्वयं के डीएनए की अनुपस्थिति में, क्लोरोप्लास्ट नहीं बनते हैं।
क्लोरोप्लास्ट को मुख्य सेलुलर अंग माना जा सकता है, क्योंकि वे सौर ऊर्जा के परिवर्तन की श्रृंखला में पहले हैं, जिसके परिणामस्वरूप मानवता को भोजन और ईंधन दोनों प्राप्त होते हैं। क्लोरोप्लास्ट में न केवल प्रकाश संश्लेषण होता है। वे अमीनो एसिड और फैटी एसिड के संश्लेषण में भी शामिल हैं, स्टार्च के अस्थायी भंडार के भंडार के रूप में काम करते हैं।
क्रोमोप्लास्ट रंजित प्लास्टिड हैं। विविध रूप में, उनके पास क्लोरोफिल नहीं है, लेकिन कैरोटीनॉयड को संश्लेषित और संचित करते हैं, जो फूलों, पुरानी पत्तियों, फलों और जड़ों को पीला, नारंगी, लाल रंग देते हैं। क्रोमोप्लास्ट क्लोरोप्लास्ट से विकसित हो सकते हैं, जो इस मामले में क्लोरोफिल और आंतरिक झिल्ली संरचनाओं को खो देते हैं, कैरोटीनॉयड जमा करते हैं। यह कई फलों के पकने के दौरान होता है। क्रोमोप्लास्ट कीड़ों और अन्य जानवरों को आकर्षित करते हैं जिनके साथ वे विकसित हुए हैं।
ल्यूकोप्लास्ट गैर-रंजित प्लास्टिड हैं। उनमें से कुछ स्टार्च (एमाइलोप्लास्ट) को संश्लेषित करते हैं, अन्य लिपिड और प्रोटीन सहित विभिन्न पदार्थ बनाने में सक्षम हैं। प्रकाश में, ल्यूकोप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं।
प्रोप्लास्टिड्स छोटे, रंगहीन या हल्के हरे रंग के, अविभाजित प्लास्टिड्स होते हैं जो जड़ों और अंकुरों की मेरिस्टेमेटिक (विभाजित) कोशिकाओं में पाए जाते हैं। वे अन्य, अधिक विभेदित प्लास्टिड्स - क्लोरोप्लास्ट्स, क्रोमोप्लास्ट्स और एमिनोप्लास्ट्स के पूर्ववर्ती हैं। यदि प्रकाश की कमी के कारण प्रोटोप्लास्टिड्स के विकास में देरी हो रही है, तो उनमें एक या एक से अधिक प्रोलामेलर निकाय दिखाई दे सकते हैं, जो ट्यूबलर झिल्ली के अर्ध-क्रिस्टलीय संचय हैं। प्रोलामेलर बॉडी वाले प्लास्टिड्स को इटियोप्लास्ट्स कहा जाता है। प्रकाश में, एटिओप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं, जबकि प्रोलामेलर निकायों की झिल्लियां थायलाकोइड्स बनाती हैं। इटियोप्लास्ट पौधों की पत्तियों में बनते हैं जो अंधेरे में होते हैं। बीज भ्रूण के प्रोटोप्लास्ट पहले इटियोप्लास्ट में बदलते हैं, जिससे क्लोरोप्लास्ट प्रकाश में विकसित होते हैं। प्लास्टिड्स को एक प्रकार से दूसरे प्रकार में अपेक्षाकृत आसान संक्रमणों की विशेषता है। प्लास्टिड्स, बैक्टीरिया की तरह, दो में विभाजित करके प्रजनन करते हैं। मेरिस्टेमेटिक कोशिकाओं में, प्रोटोप्लास्टिड्स के विभाजन का समय लगभग कोशिका विभाजन के समय के साथ मेल खाता है। हालांकि, परिपक्व कोशिकाओं में, अधिकांश प्लास्टिड परिपक्व प्लास्टिड्स के विभाजन के परिणामस्वरूप बनते हैं।
माइटोकॉन्ड्रिया।क्लोरोप्लास्ट की तरह, माइटोकॉन्ड्रिया दो प्राथमिक झिल्लियों से घिरा होता है। आंतरिक झिल्ली कई सिलवटों और प्रोट्रूशियंस - cristae का निर्माण करती है, जो माइटोकॉन्ड्रिया की आंतरिक सतह को काफी बढ़ा देती है। वे प्लास्टिड्स से काफी छोटे हैं, व्यास में लगभग 0.5 माइक्रोन और लंबाई और आकार में परिवर्तनशील हैं।
माइटोकॉन्ड्रिया में, श्वसन की प्रक्रिया होती है, जिसके परिणामस्वरूप कार्बनिक अणु ऊर्जा की रिहाई के साथ टूट जाते हैं और एटीपी अणुओं में स्थानांतरित हो जाते हैं, जो सभी यूकेरियोटिक कोशिकाओं का मुख्य ऊर्जा भंडार है। अधिकांश पादप कोशिकाओं में सैकड़ों या हजारों माइटोकॉन्ड्रिया होते हैं। एक सेल में उनकी संख्या सेल की एटीपी की आवश्यकता से निर्धारित होती है। माइटोकॉन्ड्रिया निरंतर गति में हैं, कोशिका के एक भाग से दूसरे भाग में जाते हैं, एक दूसरे के साथ विलय और विभाजित होते हैं। माइटोकॉन्ड्रिया आमतौर पर वहां इकट्ठा होते हैं जहां ऊर्जा की जरूरत होती है। यदि प्लाज़्मा झिल्ली सक्रिय रूप से कोशिका से कोशिका में पदार्थों को स्थानांतरित करती है, तो माइटोकॉन्ड्रिया झिल्ली की सतह के साथ स्थित होते हैं। मोटिव एककोशिकीय शैवाल में, माइटोकॉन्ड्रिया फ्लैगेल्ला के आधार पर जमा होता है, जो उनके आंदोलन के लिए आवश्यक ऊर्जा की आपूर्ति करता है।
माइटोकॉन्ड्रिया, प्लास्टिड्स की तरह, अर्ध-स्वायत्त अंग होते हैं जिनमें अपने स्वयं के प्रोटीन के संश्लेषण के लिए आवश्यक घटक होते हैं। आंतरिक झिल्ली एक द्रव मैट्रिक्स को घेरती है जिसमें प्रोटीन, आरएनए, डीएनए, बैक्टीरिया जैसे राइबोसोम और विभिन्न विलेय होते हैं। डीएनए एक या एक से अधिक न्यूक्लियॉइड्स में स्थित गोलाकार अणुओं के रूप में मौजूद होता है।
माइटोकॉन्ड्रिया और यूकेरियोटिक कोशिकाओं के क्लोरोप्लास्ट के साथ बैक्टीरिया की समानता के आधार पर, यह माना जा सकता है कि माइटोकॉन्ड्रिया और क्लोरोप्लास्ट बैक्टीरिया से उत्पन्न हुए हैं जो बड़े हेटरोट्रॉफ़िक कोशिकाओं में "शरण" पाए गए - यूकेरियोट्स के अग्रदूत।
माइक्रोबॉडीज।प्लास्टिड्स और माइटोकॉन्ड्रिया के विपरीत, जो दो झिल्लियों द्वारा सीमांकित होते हैं, माइक्रोबॉडी एक झिल्ली से घिरे गोलाकार अंग होते हैं। माइक्रोबॉडी में एक दानेदार (दानेदार) सामग्री होती है, कभी-कभी उनमें क्रिस्टलीय प्रोटीन का समावेश भी होता है। माइक्रोबॉडी एंडोप्लाज्मिक रेटिकुलम के एक या दो क्षेत्रों से जुड़े होते हैं।
कुछ माइक्रोबॉडी, जिन्हें प्रॉक्सिसोम कहा जाता है, ग्लाइकोलिक एसिड के चयापचय में एक महत्वपूर्ण भूमिका निभाते हैं, जो सीधे तौर पर प्रकाश श्वसन से संबंधित है। हरी पत्तियों में, वे माइटोकॉन्ड्रिया और क्लोरोप्लास्ट से जुड़े होते हैं। अन्य माइक्रोबॉडीज, जिन्हें ग्लायॉक्सिसोम कहा जाता है, में वसा को कार्बोहाइड्रेट में बदलने के लिए आवश्यक एंजाइम होते हैं। यह कई बीजों में अंकुरण के दौरान होता है।
रिक्तिकाएं -ये झिल्ली द्वारा परिसीमित कोशिका के क्षेत्र हैं, जो तरल - कोशिका रस से भरे होते हैं। वे एक टोनोप्लास्ट (वैक्यूलर मेम्ब्रेन) से घिरे होते हैं।
एक युवा पादप कोशिका में कई छोटे रिक्तिकाएँ होती हैं, जो कोशिका आयु के रूप में एक बड़ी रसधानी में विलीन हो जाती हैं। एक परिपक्व कोशिका में, इसकी मात्रा का 90% तक रिक्तिका द्वारा कब्जा किया जा सकता है। इस मामले में, साइटोप्लाज्म कोशिका झिल्ली को एक पतली परिधीय परत के रूप में दबाया जाता है। कोशिका आकार में वृद्धि मुख्य रूप से रसधानी की वृद्धि के कारण होती है। नतीजतन, टर्गर दबाव उत्पन्न होता है और ऊतक लोच बनाए रखा जाता है। यह रिक्तिका और टोनोप्लास्ट के मुख्य कार्यों में से एक है।
रस का मुख्य घटक पानी है, बाकी पौधे के प्रकार और उसकी शारीरिक अवस्था के आधार पर भिन्न होता है। रिक्तिका में लवण, शर्करा, कम अक्सर प्रोटीन होते हैं। टोनोप्लास्ट रिक्तिका में कुछ आयनों के परिवहन और संचय में सक्रिय भूमिका निभाता है। सेल सैप में आयनों की सांद्रता पर्यावरण में इसकी सांद्रता से काफी अधिक हो सकती है। कुछ पदार्थों की उच्च सामग्री के साथ, रिक्तिका में क्रिस्टल बनते हैं। सबसे आम कैल्शियम ऑक्सालेट क्रिस्टल होते हैं, जिनका एक अलग आकार होता है।
रिक्तिकाएं चयापचय उत्पादों (चयापचय) के संचय के स्थान हैं। ये प्रोटीन, अम्ल और यहां तक कि मनुष्य के लिए विषाक्त पदार्थ (एल्कलॉइड) भी हो सकते हैं। पिगमेंट अक्सर जमा होते हैं। नीला, बैंगनी, बैंगनी, गहरा लाल, किरमिजी पौधे की कोशिकाओं को एंथोसायनिन समूह से वर्णक देते हैं। अन्य पिगमेंट के विपरीत, वे पानी में अच्छी तरह से घुल जाते हैं और सेल सैप में पाए जाते हैं। वे कई सब्जियों (मूली, शलजम, गोभी), फल (अंगूर, आलूबुखारा, चेरी), फूल (कॉर्नफ्लॉवर, जेरेनियम, डेल्फीनियम, गुलाब, peonies) के लाल और नीले रंग का निर्धारण करते हैं। कभी-कभी ये वर्णक पत्तियों में क्लोरोफिल को ढक लेते हैं, उदाहरण के लिए, सजावटी लाल मेपल में। एंथोसायनिन रंग शरद ऋतु चमकदार लाल छोड़ देता है। वे ठंडी धूप के मौसम में बनते हैं, जब पत्तियों में क्लोरोफिल का संश्लेषण बंद हो जाता है। पत्तियों में, जब एंथोसायनिन नहीं बनते हैं, क्लोरोफिल के विनाश के बाद, क्लोरोप्लास्ट के पीले-नारंगी कैरोटीनॉयड ध्यान देने योग्य हो जाते हैं। ठंडे, स्पष्ट शरद ऋतु में पत्तियाँ सबसे चमकीले रंग की होती हैं।
कोशिका में उनके घटकों के संचलन में, रिक्तिकाएं मैक्रोमोलेक्युलस के विनाश में शामिल होती हैं। राइबोसोम, माइटोकॉन्ड्रिया, प्लास्टिड, रिक्तिका में गिरकर नष्ट हो जाते हैं। इस पाचन गतिविधि के अनुसार, उनकी तुलना लाइसोसोम - पशु कोशिकाओं के अंग से की जा सकती है।
एंडोप्लाज्मिक रेटिकुलम (रेटिकुलम) से रिक्तिकाएं बनती हैं
राइबोसोम।छोटे कण (17 - 23nm), जिसमें प्रोटीन और RNA की लगभग समान मात्रा होती है। राइबोसोम में, अमीनो एसिड मिलकर प्रोटीन बनाते हैं। सक्रिय चयापचय वाले कोशिकाओं में उनमें से अधिक हैं। राइबोसोम कोशिका के साइटोप्लाज्म में स्वतंत्र रूप से स्थित होते हैं या एंडोप्लाज्मिक रेटिकुलम (80S) से जुड़े होते हैं। वे नाभिक (80S), माइटोकॉन्ड्रिया (70S), प्लास्टिड्स (70S) में भी पाए जाते हैं।
राइबोसोम एक जटिल बना सकते हैं जिस पर समान पॉलीपेप्टाइड्स का एक साथ संश्लेषण होता है, जिसके बारे में जानकारी एक अणु और आरएनए से ली जाती है। इस तरह के एक जटिल को पॉलीराइबोसोम (पॉलीसोम) कहा जाता है। बड़ी मात्रा में प्रोटीन को संश्लेषित करने वाली कोशिकाओं में पॉलीसोम्स की एक व्यापक प्रणाली होती है, जो अक्सर परमाणु लिफाफे की बाहरी सतह से जुड़ी होती हैं।
अन्तः प्रदव्ययी जलिका।यह अनिश्चितकालीन सीमा की एक जटिल त्रि-आयामी झिल्ली प्रणाली है। ईआर के संदर्भ में उनके बीच एक संकीर्ण पारदर्शी स्थान के साथ दो प्राथमिक झिल्लियों की तरह दिखता है। ईआर का आकार और लंबाई सेल प्रकार, इसकी चयापचय गतिविधि और भेदभाव के चरण पर निर्भर करती है। कोशिकाओं में जो प्रोटीन को स्रावित या संग्रहीत करते हैं, ईआर को सपाट थैली या सिस्टर्न के आकार का होता है, जिसकी बाहरी सतह पर कई राइबोसोम जुड़े होते हैं। इस रेटिकुलम को रफ एंडोप्लाज्मिक रेटिकुलम कहा जाता है। एक चिकनी ईआर आमतौर पर आकार में ट्यूबलर होती है। खुरदरी और चिकनी अंतर्द्रव्यी जालिका एक ही कोशिका में मौजूद हो सकती है। एक नियम के रूप में, उनके बीच कई संबंध हैं।
एंडोप्लाज्मिक रेटिकुलम सेल की संचार प्रणाली के रूप में कार्य करता है। यह कर्नेल के बाहरी आवरण से जुड़ा होता है। वास्तव में, ये दो संरचनाएं एकल झिल्ली प्रणाली बनाती हैं। जब कोशिका विभाजन के दौरान परमाणु लिफाफा फट जाता है, तो इसके टुकड़े ER अंशों के समान हो जाते हैं। एंडोप्लाज्मिक रेटिकुलम पदार्थों के परिवहन के लिए एक प्रणाली है: प्रोटीन, लिपिड, कार्बोहाइड्रेट, कोशिका के विभिन्न भागों में। पड़ोसी कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम साइटोप्लाज्मिक स्ट्रैंड्स - प्लास्मोडेस्मेटा - से जुड़े होते हैं जो कोशिका झिल्ली से गुजरते हैं।
कोशिका झिल्लियों के संश्लेषण के लिए एंडोप्लाज्मिक रेटिकुलम मुख्य स्थल है। कुछ पादप कोशिकाओं में, रसधानियों और सूक्ष्म निकायों की झिल्लियों, तानाशाहों के गढ्ढों का निर्माण होता है।
गॉल्जीकाय।इस शब्द का प्रयोग एक सेल में सभी तानाशाही, या गोल्गी निकायों को संदर्भित करने के लिए किया जाता है। डिक्टियोसोम फ्लैट, डिस्क के आकार के पुटिकाओं या सिस्टर्न के समूह हैं, जो किनारों के साथ नलिकाओं की एक जटिल प्रणाली में शाखा करते हैं। उच्च पौधों में डिक्टायोसोम में एक साथ एकत्रित 4 - 8 कुंड होते हैं।
आमतौर पर, टैंकों के एक पैकेट में, एक विकासशील और परिपक्व पक्ष प्रतिष्ठित होता है। बनने वाले गढ्ढों की झिल्लियां संरचना में ईआर की झिल्लियों से मिलती-जुलती हैं, और परिपक्व हौदों की झिल्लियां प्लाज्मा झिल्ली से मिलती-जुलती हैं।
डिक्टायोसोम स्राव में शामिल होते हैं और अधिकांश उच्च पौधों में कोशिका झिल्लियों के निर्माण में शामिल होते हैं। डिक्टायोसोम द्वारा संश्लेषित कोशिका भित्ति पॉलीसेकेराइड पुटिकाओं में जमा हो जाते हैं, जो तब परिपक्व होने वाले सिस्टर्न से अलग हो जाते हैं। ये स्रावी पुटिकाएं सीएमपी के साथ माइग्रेट और फ्यूज हो जाती हैं; जबकि उनमें निहित पॉलीसेकेराइड कोशिका झिल्ली में निर्मित होते हैं। कुछ पदार्थ जो तानाशाहों में जमा होते हैं, अन्य संरचनाओं में बनते हैं, उदाहरण के लिए, ईआर में, और फिर तानाशाहों में ले जाया जाता है, जहां स्राव से पहले उन्हें संशोधित (संशोधित) किया जाता है। उदाहरण के लिए, ग्लाइकोप्रोटीन कोशिका झिल्ली की एक महत्वपूर्ण निर्माण सामग्री है। प्रोटीन भाग को किसी न किसी ईआर के पॉलीसोम द्वारा संश्लेषित किया जाता है, कार्बोहाइड्रेट भाग को डिक्टायोसोम में संश्लेषित किया जाता है, जहां दोनों भाग ग्लाइकोप्रोटीन बनाने के लिए संयोजित होते हैं।
झिल्लियां गतिशील, मोबाइल संरचनाएं हैं जो लगातार अपना आकार और क्षेत्र बदलती रहती हैं। एंडोप्लाज्मिक सिस्टम की अवधारणा झिल्ली की गतिशीलता पर आधारित है। इस अवधारणा के अनुसार, माइटोकॉन्ड्रिया और प्लास्टिड्स की झिल्लियों को छोड़कर, साइटोप्लाज्म की आंतरिक झिल्लियां एक ही पूरी होती हैं और एंडोप्लाज्मिक रेटिकुलम से उत्पन्न होती हैं। मध्यवर्ती पुटिका चरण के माध्यम से एंडोप्लाज्मिक रेटिकुलम से डिक्टायोसोम के नए कुंड बनते हैं, और स्रावी पुटिकाएं जो डिक्टीयोसोम से अलग होती हैं, अंततः प्लाज्मा झिल्ली के निर्माण में योगदान करती हैं। इस प्रकार, एंडोप्लाज्मिक रेटिकुलम और डिक्टियोसोम एक कार्यात्मक इकाई बनाते हैं जिसमें डिक्टायोसोम प्लाज्मा जैसी झिल्ली में एंडोप्लाज्मिक रेटिकुलम जैसी झिल्लियों को परिवर्तित करने की प्रक्रिया में मध्यवर्ती संरचनाओं की भूमिका निभाते हैं। उन ऊतकों में जिनकी कोशिकाएं बढ़ती हैं और कमजोर रूप से विभाजित होती हैं, झिल्ली के घटकों को लगातार अद्यतन किया जाता है।
सूक्ष्मनलिकाएंलगभग सभी यूकेरियोटिक कोशिकाओं में पाया जाता है। वे लगभग 24 एनएम के व्यास के साथ बेलनाकार संरचनाएं हैं। इनकी लंबाई अलग-अलग होती है। प्रत्येक नलिका ट्यूबुलिन नामक प्रोटीन की उपइकाइयों से बनी होती है। उपइकाइयां केंद्रीय गुहा के चारों ओर 13 अनुदैर्ध्य तंतु बनाती हैं। सूक्ष्मनलिकाएं गतिशील संरचनाएं हैं, वे नियमित रूप से नष्ट हो जाती हैं और कोशिका चक्र के कुछ चरणों में बनती हैं। इनका संयोजन विशेष स्थानों पर होता है, जिन्हें सूक्ष्मनलिकाय संगठन केंद्र कहते हैं। पादप कोशिकाओं में, उनके पास कमजोर व्यक्त अनाकार संरचना होती है।
सूक्ष्मनलिकाएं के कार्य: कोशिका झिल्ली के निर्माण में भाग लेते हैं; बनाने वाली झिल्ली के लिए तानाशाहों के सीधे बुलबुले, जैसे धुरी के धागे जो एक विभाजित कोशिका में बनते हैं; सेल प्लेट (बेटी कोशिकाओं के बीच प्रारंभिक सीमा) के निर्माण में भूमिका निभाते हैं। इसके अलावा, सूक्ष्मनलिकाएं फ्लैगेल्ला और सिलिया का एक महत्वपूर्ण घटक हैं, जिसके संचलन में वे महत्वपूर्ण भूमिका निभाते हैं।
सूक्ष्म तंतु,सूक्ष्मनलिकाएं की तरह, वे लगभग सभी यूकेरियोटिक कोशिकाओं में पाए जाते हैं। वे 5-7 एनएम मोटे लंबे तंतु होते हैं, जिनमें सिकुड़ा हुआ प्रोटीन एक्टिन होता है। उच्च पौधों की कई कोशिकाओं में माइक्रोफ़िल्मेंट्स के बंडल पाए जाते हैं। जाहिर है, वे साइटोप्लाज्म की धाराओं में एक महत्वपूर्ण भूमिका निभाते हैं। माइक्रोफ़िल्मेंट्स, सूक्ष्मनलिकाएं के साथ मिलकर एक लचीला नेटवर्क बनाते हैं जिसे साइटोस्केलेटन कहा जाता है।
आधार पदार्थलंबे समय तक एक सजातीय (सजातीय) प्रोटीन युक्त समाधान माना जाता था जिसमें कम संख्या में संरचनाएं या यहां तक कि संरचना रहित होती थी। हालांकि, वर्तमान में, एक उच्च-वोल्टेज इलेक्ट्रॉन माइक्रोस्कोप का उपयोग करके, यह स्थापित किया गया है कि जमीनी पदार्थ एक त्रि-आयामी जाली है जो पतली (3–6 एनएम व्यास) किस्में से निर्मित होती है जो पूरे सेल को भरती है। सूक्ष्मनलिकाएं और माइक्रोफ़िल्मेंट्स सहित साइटोप्लाज्म के अन्य घटक इस माइक्रोट्रैब्युलर जाली से निलंबित हैं।
माइक्रोट्राबेकुलर संरचना प्रोटीन स्ट्रैंड्स की एक जाली है, जिसके बीच का स्थान पानी से भरा होता है। पानी के साथ, जाली में जेल की स्थिरता होती है; जेल जिलेटिनस निकायों की तरह दिखता है।
ऑर्गेनेल माइक्रोट्राबेकुलर जाली से जुड़े होते हैं। जाली कोशिका के अलग-अलग हिस्सों के बीच संचार प्रदान करती है और इंट्रासेल्युलर परिवहन को निर्देशित करती है।
लिपिड बूँदें- एक गोलाकार आकार की संरचनाएं, एक प्रकाश सूक्ष्मदर्शी के तहत एक पादप कोशिका के साइटोप्लाज्म को ग्रैन्युलैरिटी देती हैं। इलेक्ट्रॉन माइक्रोग्राफ पर, वे अनाकार दिखते हैं। प्लास्टिड्स में बहुत समान लेकिन छोटी बूंदें पाई जाती हैं।
लिपिड बूँदें, गलती से उन्हें ऑर्गेनेल समझकर, उन्हें स्फेरोसोम्स कहते हैं और मानते हैं कि वे एक या दो परत वाली झिल्ली से घिरे हुए हैं। हालाँकि, हाल के आंकड़े बताते हैं कि लिपिड बूंदों में झिल्ली नहीं होती है, लेकिन प्रोटीन के साथ लेपित हो सकते हैं।
एर्गास्टिक पदार्थ -ये प्रोटोप्लास्ट के "निष्क्रिय उत्पाद" हैं: आरक्षित पदार्थ या अपशिष्ट उत्पाद। वे कोशिका चक्र की विभिन्न अवधियों में प्रकट और गायब हो सकते हैं। स्टार्च अनाज, क्रिस्टल, एंथोसायनिन पिगमेंट और लिपिड बूंदों को छोड़कर। इनमें रेजिन, गोंद, टैनिन और प्रोटीन शामिल हैं। एर्गैस्टिक पदार्थ कोशिका झिल्ली का हिस्सा होते हैं, साइटोप्लाज्म और ऑर्गेनेल का मुख्य पदार्थ, जिसमें रिक्तिकाएं भी शामिल हैं।
फ्लैगेल्ला और सिलिया -वे पतली, बालों जैसी संरचनाएं हैं जो कई यूकेरियोटिक कोशिकाओं की सतह से फैली हुई हैं। उनका एक स्थिर व्यास है, लेकिन लंबाई 2 से 150 माइक्रोन तक भिन्न होती है। परंपरागत रूप से, उनमें से लंबे और कुछ को फ्लैगेल्ला कहा जाता है, और छोटे और अधिक कई को सिलिया कहा जाता है। इन दो प्रकार की संरचनाओं के बीच कोई स्पष्ट अंतर नहीं है, इसलिए फ्लैगेलम शब्द का उपयोग दोनों को संदर्भित करने के लिए किया जाता है।
कुछ शैवालों तथा कवकों में कशाभिका (Flagella) चलन अंग होते हैं, जिनकी सहायता से ये जल में गति करते हैं। पौधों में (उदाहरण के लिए, काई, लिवरवॉर्ट्स, फ़र्न, कुछ जिम्नोस्पर्म), केवल जर्म कोशिकाओं (युग्मक) में फ्लैगेल्ला होता है।
प्रत्येक फ्लैगेलम का एक विशिष्ट संगठन होता है। सूक्ष्मनलिकाएं के 9 जोड़े की एक बाहरी रिंग फ्लैगेलम के केंद्र में स्थित दो अतिरिक्त सूक्ष्मनलिकाएं को घेरे हुए है। एंजाइम युक्त "हैंडल" प्रत्येक बाहरी जोड़े के एक सूक्ष्मनलिका से फैलता है। यह यूकेरियोटिक जीवों में सभी कशाभों में पाया जाने वाला बुनियादी 9+2 संगठन पैटर्न है। यह माना जाता है कि फ्लैगेल्ला की गति सूक्ष्मनलिकाएं के फिसलने पर आधारित होती है, जबकि सूक्ष्मनलिकाएं के बाहरी जोड़े बिना संकुचन के एक दूसरे के साथ चलते हैं। एक दूसरे के सापेक्ष सूक्ष्मनलिकाएं के जोड़े के फिसलने से फ्लैगेलम का स्थानीय झुकाव होता है।
फ्लैगेल्ला साइटोप्लाज्मिक बेलनाकार संरचनाओं से "बढ़ता है" जिसे बेसल बॉडी कहा जाता है, जो फ्लैगेलम का बेसल भाग भी बनाता है। बेसल निकायों में एक फ्लैगेलम जैसी आंतरिक संरचना होती है, सिवाय इसके कि बाहरी नलिकाओं को जोड़े के बजाय ट्रिपल में व्यवस्थित किया जाता है, और केंद्रीय नलिकाएं अनुपस्थित होती हैं।
कोशिका भित्ति।कोशिका भित्ति प्रोटोप्लास्ट के आकार को परिसीमित करती है और रिक्तिका द्वारा पानी के अवशोषण के कारण इसके टूटने को रोकती है।
कोशिका भित्ति में विशिष्ट कार्य होते हैं जो न केवल कोशिका और उस ऊतक के लिए महत्वपूर्ण होते हैं जिसमें कोशिका स्थित होती है, बल्कि पूरे पौधे के लिए भी महत्वपूर्ण होती है। कोशिका भित्ति पदार्थों के अवशोषण, परिवहन और उत्सर्जन में एक आवश्यक भूमिका निभाती है, और इसके अलावा, लाइसोसोमल या पाचन गतिविधि उनमें केंद्रित हो सकती है।
सेल की दीवार के घटक। कोशिका भित्ति का सबसे विशिष्ट घटक सेल्युलोज है, जो काफी हद तक इसकी वास्तुकला को निर्धारित करता है। सेल्युलोज अणु अंत से अंत तक जुड़े ग्लूकोज अणुओं को दोहराते हुए बने होते हैं। लंबे पतले सेल्युलोज अणुओं को 10 - 25 एनएम की मोटाई के साथ माइक्रोफाइब्रिल में जोड़ा जाता है। माइक्रोफ़ाइब्रिल्स आपस में जुड़ते हैं और पतले धागे बनाते हैं, जो बदले में एक दूसरे के चारों ओर लपेटे जा सकते हैं, जैसे कि रस्सी में किस्में। ऐसी प्रत्येक "रस्सी", या मैक्रोफिब्रिल की मोटाई लगभग 0.5 माइक्रोन होती है, जो 4 माइक्रोन की लंबाई तक पहुंचती है। मैक्रोफाइब्रिल समान आकार के स्टील के तार जितने मजबूत होते हैं।
सेल की दीवार का सेल्यूलोज फ्रेम इसके साथ जुड़े सेल्यूलोज मैट्रिक्स अणुओं से भरा होता है। इसमें पॉलीसेकेराइड होते हैं जिन्हें हेमिकेलुलोज कहा जाता है, और पेक्टिन, या पेक्टिन, रासायनिक रूप से हेमिकेलुलोज के बहुत करीब होते हैं।
कोशिका भित्ति का एक अन्य घटक, लिग्निन, सेल्युलोज के बाद पादप कोशिकाओं में सबसे प्रचुर मात्रा में पाया जाने वाला बहुलक है। लिग्निन दीवार की कठोरता को बढ़ाता है और आमतौर पर उन कोशिकाओं में पाया जाता है जो एक सहायक या यांत्रिक कार्य करती हैं।
क्यूटिन, सुबेरिन, मोम - आमतौर पर पौधों के सुरक्षात्मक ऊतकों के गोले में जमा होते हैं। क्यूटिन, उदाहरण के लिए, एपिडर्मिस की कोशिका झिल्लियों में पाया जाता है, और सुबेरिन द्वितीयक सुरक्षात्मक ऊतक, कॉर्क में पाया जाता है। दोनों पदार्थ वैक्स के संयोजन में पाए जाते हैं और पौधे से अत्यधिक पानी के नुकसान को रोकते हैं।
कोशिका भित्ति की परतें। पादप कोशिकाओं की दीवार की मोटाई व्यापक रूप से भिन्न होती है, जो पादप संरचना में कोशिकाओं की भूमिका और स्वयं कोशिका की आयु पर निर्भर करती है। एक इलेक्ट्रॉन माइक्रोस्कोप के तहत, पौधे की कोशिका दीवार में दो परतें दिखाई देती हैं: माध्यिका पटल (जिसे अंतरकोशिकीय पदार्थ भी कहा जाता है), और प्राथमिक कोशिका भित्ति। कई कोशिकाएं एक और परत बिछाती हैं, द्वितीयक कोशिका भित्ति। मीडियन प्लेट आसन्न कोशिकाओं की प्राथमिक दीवारों के बीच स्थित होती है। द्वितीयक दीवार, यदि कोई हो, प्राथमिक सेल दीवार की आंतरिक सतह पर सेल प्रोटोप्लास्ट द्वारा जमा की जाती है।
बीच की थाली। मीडियन लैमिना में मुख्य रूप से पेक्टिन होता है। जहां एक कोशिका भित्ति उत्पन्न होनी चाहिए, दो नवगठित कोशिकाओं के बीच, एंडोप्लाज़मिक रेटिकुलम के नलिकाओं का एक घना जाल और गोल्गी तंत्र (डिक्टायोसोम) का कुंड पहले नोट किया जाता है। फिर इस जगह में पेक्टिन (पॉलीसेकेराइड से) से भरे बुलबुले दिखाई देते हैं। इन बुलबुलों को गॉल्जी उपकरण की टंकियों से अलग किया जाता है। प्रारंभिक कोशिका भित्ति में विभिन्न पॉलीसेकेराइड होते हैं, जिनमें से मुख्य हैं पेक्टिन और हेमिकेलुलोज। बाद में, इसमें सघन पदार्थ - सेल्युलोज और लिग्निन होते हैं।
प्राथमिक कोशिका भित्ति। यह सेल्युलोज आवरण की एक परत है जो कोशिका वृद्धि से पहले या उसके दौरान जमा होती है। सेलूलोज़, हेमिसेल्यूलोज़ और पेक्टिन के अलावा, प्राथमिक झिल्ली में ग्लाइकोप्रोटीन होता है। प्राथमिक गोले लिग्निफाई कर सकते हैं। पेक्टिन घटक नमनीयता प्रदान करता है, जो प्राथमिक खोल को जड़, तना या पत्ती के रूप में विस्तारित करने की अनुमति देता है।
सक्रिय रूप से विभाजित होने वाली कोशिकाओं (प्रकाश संश्लेषण, श्वसन और स्राव की प्रक्रियाओं में शामिल अधिकांश परिपक्व कोशिकाओं) में प्राथमिक झिल्ली होती है। एक प्राथमिक झिल्ली और एक जीवित प्रोटोप्लास्ट वाली ऐसी कोशिकाएं अपने विशिष्ट आकार को खोने, विभाजित करने और एक नए प्रकार की कोशिका में अंतर करने में सक्षम होती हैं। वे पौधों में घाव भरने और ऊतक पुनर्जनन में शामिल हैं।
प्राथमिक कोशिका झिल्लियां अपनी पूरी लंबाई में मोटाई में एक समान नहीं होती हैं, लेकिन पतले खंड होते हैं, जिन्हें प्राथमिक छिद्र क्षेत्र कहा जाता है। साइटोप्लाज्म, या प्लास्मोडेस्माटा की किस्में, जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ती हैं, आमतौर पर प्राथमिक छिद्र क्षेत्रों से गुजरती हैं।
द्वितीयक कोशिका भित्ति। इस तथ्य के बावजूद कि कई पादप कोशिकाओं में केवल एक प्राथमिक दीवार होती है, कुछ में, प्रोटोप्लास्ट कोशिका के केंद्र की ओर एक द्वितीयक दीवार जमा करता है। यह आमतौर पर कोशिका वृद्धि की समाप्ति के बाद होता है और प्राथमिक झिल्ली का क्षेत्र अब नहीं बढ़ता है। इस कारण से, द्वितीयक खोल प्राथमिक से भिन्न होता है। माध्यमिक झिल्लियों की विशेष रूप से विशेष कोशिकाओं द्वारा आवश्यकता होती है जो पौधे को मजबूत करती हैं और पानी का संचालन करती हैं। द्वितीयक झिल्ली के निक्षेपण के बाद, इन कोशिकाओं के प्रोटोप्लास्ट, एक नियम के रूप में, मर जाते हैं। माध्यमिक गोले में प्राथमिक की तुलना में अधिक सेल्यूलोज होता है, और उनमें पेक्टिन पदार्थ और ग्लाइकोप्रोटीन अनुपस्थित होते हैं। द्वितीयक झिल्ली को फैलाना मुश्किल होता है, इसके मैट्रिक्स में हेमिकेलुलोज होता है।
द्वितीयक खोल में, तीन परतों को प्रतिष्ठित किया जा सकता है - बाहरी, मध्य और आंतरिक (S1, S2, S3)। द्वितीयक गोले की स्तरित संरचना उनकी ताकत में काफी वृद्धि करती है। द्वितीयक झिल्ली में सेल्युलोज माइक्रोफाइब्रिल्स प्राथमिक की तुलना में सघन रूप से जमा होते हैं। लिग्निन लकड़ी की द्वितीयक खाल का एक सामान्य घटक है।
संपर्क कोशिकाओं की झिल्लियों में छिद्र एक दूसरे के विपरीत स्थित होते हैं। दो विरोधी छिद्र और एक छिद्र झिल्ली छिद्रों की एक जोड़ी बनाते हैं। द्वितीयक झिल्लियों वाली कोशिकाओं में, दो मुख्य प्रकार के छिद्र होते हैं: सरल और सीमाबद्ध। बॉर्डर वाले छिद्रों में, द्वितीयक दीवार छिद्र गुहा के ऊपर लटकी होती है। सिंपल पोर्स में ऐसा नहीं होता है।
कोशिका भित्ति का विकास। जैसे-जैसे कोशिका बढ़ती है, कोशिका झिल्ली की मोटाई और क्षेत्रफल बढ़ता जाता है। शेल स्ट्रेचिंग एक जटिल प्रक्रिया है। यह प्रोटोप्लास्ट द्वारा नियंत्रित होता है और हार्मोन ऑक्सिन द्वारा नियंत्रित होता है।
सभी दिशाओं में समान रूप से बढ़ने वाली कोशिकाओं में, मायोफिब्रिल्स का जमाव यादृच्छिक होता है। ये मायोफिब्रिल एक अनियमित नेटवर्क बनाते हैं। ऐसी कोशिकाएं तने के मूल भाग, भंडारण के ऊतकों और इन विट्रो में कोशिका संवर्धन के दौरान पाई गईं। लम्बी कोशिकाओं में, पार्श्व आवरण के पेशीतंतुओं को बढ़ाव के अक्ष पर समकोण पर जमा किया जाता है।
मैट्रिक्स पदार्थ - पेक्टिन, हेमिकेलुलोज और ग्लाइकोप्रोटीन को डिक्टीयोसोम के पुटिकाओं में झिल्ली में स्थानांतरित किया जाता है। इसी समय, पेक्टिन बढ़ती कोशिकाओं की अधिक विशेषता है, जबकि हेमिकेलुलोज गैर-बढ़ती कोशिकाओं में प्रबल होते हैं।
सेल्युलोज माइक्रोफिब्रिल को कोशिका की सतह पर प्लाज्मा झिल्ली से जुड़े एक एंजाइम कॉम्प्लेक्स की मदद से संश्लेषित किया जाता है। सूक्ष्मतंतुओं का उन्मुखीकरण प्लाज्मा झिल्ली की आंतरिक सतह के पास स्थित सूक्ष्मनलिकाएं द्वारा नियंत्रित होता है।
प्लाज्मोडेस्मा।ये साइटोप्लाज्म के पतले धागे हैं जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ते हैं। प्लास्मोडेस्माटा या तो कहीं भी कोशिका भित्ति से होकर गुजरता है, या प्राथमिक छिद्र क्षेत्रों में या छिद्रों के जोड़े के बीच झिल्लियों में केंद्रित होता है। एक इलेक्ट्रॉन माइक्रोस्कोप के तहत, प्लास्मोडेमाटा प्लाज्मा झिल्ली के साथ पंक्तिबद्ध संकीर्ण चैनल के रूप में दिखाई देते हैं। नहर की धुरी के साथ, एक छोटी बेलनाकार ट्यूब, डेस्मोटुबुल, एक कोशिका से दूसरी कोशिका तक फैली हुई है, जो दोनों आसन्न कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम के साथ संचार करती है। कोशिका विभाजन के दौरान कई प्लास्मोडेस्माटा बनते हैं, जब ट्यूबलर एंडोप्लाज्मिक रेटिकुलम विकासशील सेल प्लेट द्वारा लिया जाता है। प्लास्मोडेस्माटा अविभाजित कोशिकाओं की झिल्लियों में भी बन सकता है। ये संरचनाएं कोशिका से कोशिका में कुछ पदार्थों का कुशल स्थानांतरण प्रदान करती हैं।
कोशिका विभाजन।बहुकोशिकीय जीवों में, कोशिका विभाजन, उनके आकार में वृद्धि के साथ, पूरे जीव के विकास का एक तरीका है। विभाजन के दौरान बनने वाली नई कोशिकाएं मूल कोशिका और एक दूसरे दोनों की संरचना और कार्य में समान होती हैं। यूकेरियोट्स में विभाजन प्रक्रिया को दो आंशिक रूप से अतिव्यापी चरणों में विभाजित किया जा सकता है: माइटोसिस और साइटोकाइनेसिस।
माइटोसिस एक नाभिक से दो बेटी नाभिक का निर्माण होता है, एक दूसरे के रूपात्मक और आनुवंशिक रूप से समतुल्य। साइटोकिनेसिस बेटी कोशिकाओं के गठन के साथ कोशिका के साइटोप्लाज्मिक भाग का विभाजन है।
कोशिका चक्र।एक जीवित कोशिका क्रमिक घटनाओं की एक श्रृंखला से गुजरती है जो कोशिका चक्र बनाती है। चक्र की अवधि स्वयं कोशिका के प्रकार और बाहरी कारकों जैसे तापमान या पोषक तत्वों की उपलब्धता के आधार पर भिन्न होती है। आमतौर पर चक्र को इंटरपेज़ और माइटोसिस के चार चरणों में विभाजित किया जाता है।
अंतरावस्था।क्रमिक माइटोटिक विभाजनों के बीच की अवधि।
इंटरपेज़ को तीन अवधियों में विभाजित किया गया है, जिसे G1, S, G2 के रूप में दर्शाया गया है।
G1 अवधि के दौरान, जो माइटोसिस के बाद शुरू होती है। इस अवधि के दौरान, विभिन्न जीवों सहित साइटोप्लाज्म की मात्रा बढ़ जाती है। इसके अलावा, वर्तमान परिकल्पना के अनुसार, पदार्थ G1 अवधि के दौरान संश्लेषित होते हैं जो या तो S अवधि और शेष चक्र को उत्तेजित या बाधित करते हैं, इस प्रकार विभाजन प्रक्रिया का निर्धारण करते हैं।
S अवधि G1 अवधि का अनुसरण करती है, जिस समय आनुवंशिक सामग्री (डीएनए) का दोहराव होता है।
G2 अवधि के दौरान, जो S का अनुसरण करता है, संरचनाएं जो सीधे माइटोसिस में शामिल होती हैं, जैसे कि धुरी घटक, बनते हैं।
कुछ कोशिकाएं असीमित संख्या में सेल चक्रों से गुजरती हैं। ये एककोशिकीय जीव और सक्रिय विकास क्षेत्रों (मेरिस्टेम) की कुछ कोशिकाएँ हैं। परिपक्वता के बाद कुछ विशिष्ट कोशिकाएं प्रजनन करने की अपनी क्षमता खो देती हैं। कोशिकाओं का तीसरा समूह, उदाहरण के लिए, घाव के ऊतक (कैलस) बनाने वाले, केवल विशेष परिस्थितियों में विभाजित करने की क्षमता को बरकरार रखते हैं।
सूत्रीविभाजन,या परमाणु विखंडन। यह एक सतत प्रक्रिया है जिसे चार चरणों में बांटा गया है: प्रोफ़ेज़, मेटाफ़ेज़, एनाफ़ेज़, टेलोफ़ेज़। माइटोसिस के परिणामस्वरूप, आनुवंशिक सामग्री जो इंटरपेज़ में दोगुनी हो गई है, दो बेटी नाभिकों के बीच समान रूप से विभाजित हो जाती है।
विभाजन के लिए एक कोशिका के संक्रमण के शुरुआती संकेतों में से एक प्लाज्मा झिल्ली के ठीक नीचे सूक्ष्मनलिकाएं के एक संकीर्ण, कुंडलाकार बैंड की उपस्थिति है। यह अपेक्षाकृत सघन बेल्ट भविष्य के माइटोटिक स्पिंडल के विषुवतीय तल में नाभिक को घेरता है। चूंकि यह प्रोफ़ेज़ से पहले प्रकट होता है, इसलिए इसे प्रीप्रोफ़ेज़ गर्डल कहा जाता है। यह माइटोटिक स्पिंडल के बाद गायब हो जाता है, सेल प्लेट के लेट टेलोफ़ेज़ में प्रकट होने से बहुत पहले, जो केंद्र से परिधि तक बढ़ता है और प्रीप्रोफ़ेज़ बेल्ट द्वारा कब्जा किए गए क्षेत्र में मातृ कोशिका झिल्ली के साथ विलीन हो जाता है।
प्रोफ़ेज़। प्रोफ़ेज़ की शुरुआत में, गुणसूत्र नाभिक के अंदर बिखरे हुए लंबे धागों के समान होते हैं। फिर, जैसे-जैसे तंतु छोटे और मोटे होते जाते हैं, कोई यह देख सकता है कि प्रत्येक गुणसूत्र एक नहीं, बल्कि दो गुंथे हुए तंतुओं से बना होता है जिन्हें क्रोमैटिड कहा जाता है। देर से प्रोफ़ेज़ में, प्रत्येक गुणसूत्र के दो छोटे युग्मित क्रोमैटिड समानांतर में एक साथ होते हैं, जो सेंट्रोमियर नामक एक संकीर्ण खंड से जुड़े होते हैं। यह प्रत्येक गुणसूत्र पर एक विशिष्ट स्थिति रखता है और गुणसूत्र को अलग-अलग लंबाई की दो भुजाओं में विभाजित करता है।
सूक्ष्मनलिकाएं धुरी की धुरी के साथ नाभिक की सतह के समानांतर स्थित होती हैं। यह माइटोटिक स्पिंडल असेंबली की सबसे पहली अभिव्यक्ति है।
प्रोफ़ेज़ के अंत तक, नाभिक धीरे-धीरे अपनी स्पष्ट रूपरेखा खो देता है और अंत में गायब हो जाता है। इसके तुरंत बाद, परमाणु लिफाफा भी बिखर जाता है।
रूपक। मेटाफ़ेज़ की शुरुआत में, स्पिंडल, जो एक त्रि-आयामी संरचना है, मध्य में सबसे चौड़ा और ध्रुवों की ओर पतला होता है, पहले नाभिक द्वारा कब्जा कर लिया गया स्थान लेता है। स्पिंडल फाइबर सूक्ष्मनलिकाएं के बंडल हैं। मेटाफ़ेज़ के दौरान, क्रोमोसोम, जिनमें से प्रत्येक में दो क्रोमैटिड होते हैं, को व्यवस्थित किया जाता है ताकि उनके सेंट्रोमर्स स्पिंडल के इक्वेटोरियल प्लेन में स्थित हों। इसके सेंट्रोमियर के साथ, प्रत्येक गुणसूत्र स्पिंडल थ्रेड्स से जुड़ा होता है। हालाँकि, कुछ किस्में गुणसूत्रों से जुड़े बिना एक ध्रुव से दूसरे ध्रुव तक जाती हैं।
जब सभी गुणसूत्र विषुवतीय तल में स्थित होते हैं, तो मेटाफ़ेज़ पूरा हो जाता है। गुणसूत्र विभाजित होने के लिए तैयार होते हैं।
पश्चावस्था। प्रत्येक गुणसूत्र के क्रोमैटिड अलग हो जाते हैं। अब ये पुत्री गुणसूत्र हैं। सबसे पहले, सेंट्रोमियर विभाजित होता है, और दो बेटी गुणसूत्र विपरीत ध्रुवों पर खींचे जाते हैं। इस मामले में, सेंट्रोमर्स आगे बढ़ते हैं, और गुणसूत्रों की भुजाएँ पीछे की ओर खिंचती हैं। क्रोमोसोम से जुड़े स्पिंडल फिलामेंट्स छोटे हो जाते हैं, क्रोमैटिड्स को अलग करने और विपरीत दिशाओं में बेटी क्रोमोसोम के आंदोलन में योगदान करते हैं।
टेलोफ़ेज़। टेलोफ़ेज़ में, गुणसूत्रों के दो समान समूहों का पृथक्करण पूरा हो जाता है, जबकि उनमें से प्रत्येक के चारों ओर एक परमाणु झिल्ली बन जाती है। रफ रेटिकुलम इसमें सक्रिय भाग लेता है। धुरी तंत्र गायब हो जाता है। टेलोफ़ेज़ के दौरान, गुणसूत्र अपनी रूपरेखा की स्पष्टता खो देते हैं, फैल जाते हैं, पतले धागों में बदल जाते हैं। न्यूक्लियोली को बहाल किया जाता है। जब गुणसूत्र अदृश्य हो जाते हैं, तो माइटोसिस पूरा हो जाता है। दो संतति केंद्रक अंतरावस्था में प्रवेश करते हैं। वे आनुवंशिक रूप से एक दूसरे के और मूल नाभिक के बराबर हैं। यह बहुत महत्वपूर्ण है, क्योंकि अनुवांशिक कार्यक्रम, और इसके साथ सभी संकेतों को बेटी जीवों में स्थानांतरित किया जाना चाहिए।
माइटोसिस की अवधि विभिन्न जीवों में भिन्न होती है और ऊतक के प्रकार पर निर्भर करती है। हालाँकि, प्रोफ़ेज़ सबसे लंबा है और एनाफ़ेज़ सबसे छोटा है। रूट टिप की कोशिकाओं में, प्रोफ़ेज़ की अवधि 1-2 घंटे होती है; रूपक - 5 - 15 मिनट; पश्चावस्था - 2 - 10 मिनट; टेलोफ़ेज़ - 10 - 30 मिनट। इंटरपेज़ की अवधि 12 से 30 घंटे तक है।
कई यूकेरियोटिक कोशिकाओं में, माइटोटिक स्पिंडल के गठन के लिए जिम्मेदार सूक्ष्मनलिका आयोजन केंद्र सेंट्रीओल्स से जुड़े होते हैं।
साइटोकिनेसिस।यह साइटोप्लाज्म के विभाजन की प्रक्रिया है। अधिकांश जीवों में, कोशिकाएँ कोशिका झिल्ली को पीछे खींचकर और एक विखंडन खांचे का निर्माण करके विभाजित होती हैं, जो धीरे-धीरे गहरा होता है, माइटोटिक स्पिंडल के शेष तंतुओं को संकुचित करता है। सभी पौधों (ब्रायोफाइट्स और संवहनी) में और कुछ शैवाल में, कोशिकाएँ किसके गठन के कारण विभाजित होती हैं एक सेल प्लेट।
शुरुआती टेलोफेज में, दो बेटी नाभिकों के बीच एक बैरल के आकार का तंतुओं की प्रणाली जिसे फेटामोप्लास्ट कहा जाता है। फैगमोप्लास्ट फाइबर, माइटोटिक स्पिंडल के तंतुओं की तरह, सूक्ष्मनलिकाएं होते हैं। फ्रैगमोप्लास्ट के विषुवतीय तल में छोटी-छोटी बूंदें दिखाई देती हैं। वे एक सेल प्लेट बनाने के लिए फ्यूज हो जाते हैं जो तब तक बढ़ता है जब तक कि वह विभाजित सेल की दीवार तक नहीं पहुंच जाता। यह दो बेटी कोशिकाओं के अलगाव को पूरा करता है। विलय की बूंदें गोल्गी तंत्र से निकलने वाले बुलबुले हैं। मूल रूप से, उनमें पेक्टिन पदार्थ होते हैं, जिनसे मध्य प्लेट बनती है। प्लेट के दोनों किनारों पर प्लाज्मा झिल्ली के निर्माण में पुटिका झिल्ली शामिल होती है। इसी समय, ट्यूबलर एंडोप्लाज्मिक रेटिकुलम के टुकड़ों से प्लास्मोडेस्मेटा बनता है।
मीडियन प्लेट के बनने के बाद, प्रत्येक प्रोटोप्लास्ट उस पर एक प्राथमिक झिल्ली जमा करता है। इसके अलावा, प्रत्येक बेटी कोशिका पूरे प्रोटोप्लास्ट के चारों ओर झिल्ली की एक नई परत जमा करती है, जो कोशिका प्लेट से उत्पन्न हुई झिल्ली को जारी रखती है। बेटी कोशिकाओं के बढ़ने पर मूल कोशिका का मूल खोल नष्ट हो जाता है।
विभिन्न प्रकार के यूकेरियोटिक माइटोसिस
पौधों और जानवरों में ऊपर वर्णित कोशिका विभाजन अप्रत्यक्ष कोशिका विभाजन का एकमात्र रूप नहीं है। माइटोसिस का सबसे सरल प्रकार है प्लूरोमिटोसिस।यह प्रोकैरियोटिक कोशिकाओं के द्विआधारी विखंडन जैसा दिखता है, जिसमें प्रतिकृति के बाद न्यूक्लियॉइड प्लाज्मा झिल्ली से जुड़े रहते हैं। झिल्ली डीएनए बंधन बिंदुओं के बीच बढ़ने लगती है और इस तरह क्रोमोसोम को कोशिका के विभिन्न भागों में फैला देती है। उसके बाद, एक कोशिका संकुचन के निर्माण के दौरान, प्रत्येक डीएनए अणु एक नई अलग कोशिका में होगा।
यूकेरियोटिक कोशिकाओं के विभाजन के लिए विशेषता सूक्ष्मनलिकाएं से निर्मित एक धुरी का गठन है। पर बंद प्लूरोमिटोसिस(इसे बंद कहा जाता है क्योंकि गुणसूत्रों का विचलन परमाणु लिफाफे को बाधित किए बिना होता है) सेंट्रीओल्स नहीं, बल्कि परमाणु झिल्ली के अंदरूनी हिस्से में स्थित अन्य संरचनाएं सूक्ष्मनलिका आयोजन केंद्र (MCMT) के रूप में भाग लेती हैं। ये अनिश्चित आकृति विज्ञान के तथाकथित ध्रुवीय निकाय हैं, जिनसे सूक्ष्मनलिकाएं फैलती हैं। इनमें से दो शरीर हैं। वे परमाणु लिफाफे से संपर्क खोए बिना एक दूसरे से अलग हो जाते हैं। इसके परिणामस्वरूप, गुणसूत्रों से जुड़े दो अर्ध-स्पिंडल बनते हैं। इस मामले में माइटोटिक स्पिंडल गठन और गुणसूत्र अलगाव की पूरी प्रक्रिया परमाणु लिफाफे के अंतर्गत होती है। इस प्रकार का माइटोसिस प्रोटोजोआ के बीच होता है और व्यापक रूप से कवक (काइट्रिडिया, ज़ाइगोमाइसेट्स, यीस्ट, ओमीसाइकेट्स, एस्कोमाइसेट्स, मायक्सोमाइसेट्स, आदि) में वितरित किया जाता है। रूपों से मिलें अर्ध-बंद प्लूरोमिटोसिसजब गठित धुरी के ध्रुवों पर परमाणु लिफाफा नष्ट हो जाता है।
माइटोसिस का अगला रूप है ऑर्थोमाइटोसिस।इस मामले में, COMTs साइटोप्लाज्म में स्थित होते हैं, शुरुआत से ही, अर्ध-स्पिंडल नहीं बनते हैं, लेकिन एक द्विध्रुवीय स्पिंडल होता है। तीन रूप हैं ऑर्थोमाइटोसिस (सामान्य माइटोसिस) अर्द्ध बंदऔर बंद किया हुआ. अर्ध-बंद ऑर्थोमाइटोसिस में, साइटोप्लाज्म में स्थित TsOMT की मदद से एक बिसिमेट्रिक स्पिंडल बनता है, ध्रुवीय क्षेत्रों के अपवाद के साथ, परमाणु लिफाफा पूरे माइटोसिस में संरक्षित होता है। दानेदार सामग्री, या सेंट्रीओल्स के द्रव्यमान को COMT के रूप में पहचाना जा सकता है। माइटोसिस का यह रूप हरे, भूरे और लाल शैवाल के जूस्पोर्स में पाया जाता है, कुछ निचले कवकों में और ग्रेगरीन में। बंद ऑर्थोमाइटोसिस के साथ, परमाणु झिल्ली पूरी तरह से संरक्षित होती है, जिसमें एक वास्तविक धुरी बनती है। सूक्ष्मनलिकाएं कार्योप्लाज्म में बनती हैं, कम बार वे आंतरिक TsOMT से बढ़ती हैं, जो परमाणु झिल्ली के साथ संबद्ध (प्लुरोमिटोसिस के विपरीत) नहीं होती है। इस प्रकार का माइटोसिस सिलियेट माइक्रोन्यूक्लि के विभाजन के लिए विशिष्ट है, लेकिन प्रोटोजोआ में भी हो सकता है। खुले ऑर्थोमाइटोसिस में, परमाणु लिफाफा पूरी तरह से विघटित हो जाता है। इस प्रकार का कोशिका विभाजन पशु जीवों, कुछ प्रोटोजोआ और उच्च पौधों की कोशिकाओं की विशेषता है। माइटोसिस का यह रूप, बदले में, सूक्ष्म और अनास्ट्रल प्रकारों द्वारा दर्शाया जाता है।
संक्षेप में समीक्षा की गई सामग्री से, यह देखा जा सकता है कि सामान्य रूप से माइटोसिस की मुख्य विशेषता विखंडन धुरी संरचनाओं की उपस्थिति है, जो सीएमटी के संबंध में बनती हैं, जो उनकी संरचना में विविध हैं।
माइटोटिक आकृति की आकृति विज्ञान
माइटोसिस के मेटाफ़ेज़ चरण में माइटोटिक उपकरण विशेष रूप से अच्छी तरह से व्यक्त किया गया है। मेटाफ़ेज़ में, कोशिका के विषुवतीय तल में, गुणसूत्र स्थित होते हैं, जिससे तथाकथित स्पिंडल धागे विपरीत दिशाओं में फैलते हैं, माइटोटिक आकृति के दो अलग-अलग ध्रुवों पर परिवर्तित होते हैं। इस प्रकार, माइटोटिक स्पिंडल गुणसूत्रों, ध्रुवों और तंतुओं का एक संग्रह है। धुरी के तंतु एकल सूक्ष्मनलिकाएं या उनके बंडल होते हैं। सूक्ष्मनलिकाएं धुरी के ध्रुवों से शुरू होती हैं, और उनमें से कुछ जाती हैं सेंट्रोमर्स कोकहाँ स्थित हैं गुणसूत्रबिंदुओंक्रोमोसोम (कीनेटोकोर सूक्ष्मनलिकाएं), कुछ विपरीत ध्रुव की ओर आगे बढ़ते हैं, लेकिन उस तक नहीं पहुंच पाते हैं। उन्हें "इंटरपोलर माइक्रोट्यूबुल्स" कहा जाता है। रेडियल सूक्ष्मनलिकाएं का एक समूह ध्रुवों से निकलता है, जिससे उनके चारों ओर एक "उज्ज्वल चमक" जैसी संरचना बनती है - ये सूक्ष्म सूक्ष्मनलिकाएं हैं।
आकारिकी द्वारा, माइटोटिक आकृतियों को सूक्ष्म और अनास्ट्रल प्रकारों में विभाजित किया जाता है।
सूक्ष्म धुरी प्रकार, या अभिसारी, इस तथ्य की विशेषता है कि इसके ध्रुवों को एक छोटे से क्षेत्र द्वारा दर्शाया जाता है जिसमें सूक्ष्मनलिकाएं अभिसरण (अभिसरण) करती हैं। आमतौर पर, सेंट्रीओल्स वाले सेंट्रोसोम सूक्ष्म स्पिंडल के ध्रुवों पर स्थित होते हैं। हालांकि, सेंट्रीओलर एस्ट्रल माइटोस के मामले ज्ञात हैं (कुछ अकशेरूकीय के अर्धसूत्रीविभाजन के दौरान)। इसके अलावा, ध्रुवों से निकलने वाली रेडियल सूक्ष्मनलिकाएं नोट की जाती हैं, जो धुरी का हिस्सा नहीं हैं, लेकिन तारकीय क्षेत्र - सिटास्टर बनाती हैं। इस प्रकार का माइटोटिक डिवीजन एक डंबल जैसा दिखता है। माइटोटिक आकृति के एनास्ट्रियल प्रकार में ध्रुवों पर सिटास्टर नहीं होते हैं। यहां धुरी के ध्रुवीय क्षेत्र चौड़े हैं, उन्हें ध्रुवीय टोपी कहा जाता है, उनमें सेंट्रीओल्स शामिल हैं। इस मामले में, धुरी के तंतु एक बिंदु से नहीं हटते हैं, लेकिन ध्रुवीय टोपी के पूरे क्षेत्र से एक विस्तृत मोर्चे (विचलन) में विचलन करते हैं। इस प्रकार की धुरी उच्च पौधों की कोशिकाओं को विभाजित करने की विशेषता है, लेकिन उच्च जानवरों में भी पाई जा सकती है। स्तनधारियों के प्रारंभिक भ्रूणजनन में, सेंट्रीओलर (अपसारी) माइटोस डिम्बाणुजनकोशिका परिपक्वता के विभाजन के दौरान और जाइगोट के विभाजन I और II में देखे जाते हैं। लेकिन पहले से ही तीसरे कोशिका विभाजन में और बाद की सभी कोशिकाओं में, वे सूक्ष्म स्पिंडल की भागीदारी के साथ विभाजित होते हैं, जिनके ध्रुवों में सेंट्रीओल्स हमेशा पाए जाते हैं। सामान्य तौर पर, माइटोसिस के सभी रूपों के लिए, उनके किनेटोफ़ोर्स, ध्रुवीय निकायों (सेंट्रोसोम्स) और स्पिंडल फाइबर के साथ गुणसूत्र सामान्य संरचनाएं बने रहते हैं।
सेंट्रोमियर और काइनेटोकोर
क्रोमोसोम की लंबाई के साथ सेंट्रोमर्स का अलग-अलग स्थानीयकरण हो सकता है। Holocentric centromeres तब होते हैं जब सूक्ष्मनलिकाएं पूरे गुणसूत्र (कुछ कीड़े, नेमाटोड, कुछ पौधे) की लंबाई के साथ जुड़ी होती हैं। मोनोसेंट्रिक सेंट्रोमियर - जब सूक्ष्मनलिकाएं एक साइट में गुणसूत्रों से जुड़ी होती हैं। मोनोसेन्ट्रिक सेंट्रोमर्स पंचर हो सकते हैं (उदाहरण के लिए, कुछ नवोदित यीस्ट में), जब केवल एक सूक्ष्मनलिका कीनेटोचोर तक पहुंचती है, और ज़ोनल, जहां सूक्ष्मनलिकाएं का एक बंडल जटिल किनेटोकोर तक पहुंचता है। सेंट्रोमियर ज़ोन की विविधता के बावजूद, ये सभी किनेटोकोर की जटिल संरचना से जुड़े हैं, जिसमें सभी यूकेरियोट्स में संरचना और कार्यों में एक मौलिक समानता है। काइनेटोकोर विशेष प्रोटीन संरचनाएं हैं, जो ज्यादातर क्रोमोसोम के सेंट्रोमियर जोन में स्थित होती हैं। ये कई प्रोटीनों से युक्त जटिल परिसर हैं। रूपात्मक रूप से, वे बहुत समान हैं, समान संरचना है, डायटम से लेकर मनुष्यों तक। वे तीन-परत संरचनाएं हैं: गुणसूत्र के शरीर से सटे आंतरिक घने परत, मध्य ढीली परत और बाहरी घने परत। कई तंतु बाहरी परत से फैलते हैं, जो कीनेटोचोर के तथाकथित रेशेदार मुकुट का निर्माण करते हैं। सामान्य रूप में, कीनेटोकोर्स क्रोमोसोम के प्राथमिक कसना के क्षेत्र में स्थित प्लेट या डिस्क की तरह दिखते हैं, सेंट्रोमियर में। आमतौर पर प्रति क्रोमोसोम या क्रोमैटिड में एक काइनेटोकोर होता है। एनाफेज से पहले, प्रत्येक बहन क्रोमैटिड पर काइनेटोकोर्स स्थित होते हैं, प्रत्येक सूक्ष्मनलिकाएं के अपने स्वयं के बंडल के साथ संचार करता है। पौधों में, काइनेटोकोर प्लेटों की तरह नहीं, बल्कि गोलार्द्धों की तरह दिखता है। कीनेटोकोर्स की कार्यात्मक भूमिका बहन क्रोमैटिड्स को एक दूसरे से बांधना है, माइटोटिक माइक्रोट्यूबुल्स को ठीक करना, क्रोमोसोम पृथक्करण को विनियमित करना और वास्तव में माइटोसिस के दौरान माइक्रोट्यूबुल्स की भागीदारी के साथ क्रोमोसोम को स्थानांतरित करना है। सामान्य तौर पर, प्रोटीन संरचनाएं, किनेटोकोर्स, एस-अवधि में दोगुनी, गुणसूत्र दोहरीकरण के समानांतर। लेकिन उनके प्रोटीन कोशिका चक्र की सभी अवधियों में गुणसूत्रों पर मौजूद होते हैं।
संयंत्र ऊतक
शैक्षिक ऊतक (मेरिस्टेम)
पौधों के शरीर में शैक्षिक ऊतक विभिन्न स्थानों पर स्थित होते हैं, इसलिए उन्हें निम्नलिखित समूहों में विभाजित किया जाता है (चित्र 0;1)।
एपिकल (एपिकल) मेरिस्टेम्स अक्षीय अंगों - तने, जड़ के शीर्ष या एपेक्स पर स्थित होते हैं। इन विभज्योतकों की सहायता से पौधों के कायिक अंग लंबाई में वृद्धि करते हैं।
पार्श्व विभज्योतक अक्षीय अंगों की विशेषता है। वहां वे क्लच के रूप में केंद्रित रूप से स्थित हैं।
इंटरक्लेरी, या इंटरक्लेरी, मेरिस्टेम एपिकल मेरिस्टेम से उत्पन्न होते हैं। ये कोशिकाओं के समूह हैं जो अभी तक गुणा करने में सक्षम नहीं हैं, लेकिन भेदभाव के मार्ग पर चल पड़े हैं। उनमें कोई प्रारंभिक कोशिकाएँ नहीं हैं, लेकिन कई विशिष्ट कोशिकाएँ हैं।
घाव मेरिस्टेम शरीर के क्षतिग्रस्त हिस्से की बहाली प्रदान करते हैं। पुनर्जनन की शुरुआत निर्विभेदीकरण से होती है, यानी विशिष्ट कोशिकाओं से विभज्योतक कोशिकाओं तक का उल्टा विकास। वे फेलोजेन में बदल जाते हैं, जो एक प्लग बनाता है जो घाव की सतह को ढकता है। अलग-अलग विभाजित कोशिकाएं, विभाजित, एक ढीले पैरेन्काइमल ऊतक - कैलस बना सकती हैं। इससे कुछ शर्तों के तहत पौधे के अंगों का निर्माण होता है।
पूर्णांक ऊतक
वे एक सीमा बाधा के रूप में कार्य करते हैं, अंतर्निहित ऊतकों को पर्यावरण से अलग करते हैं। पौधे के प्राथमिक अध्यावरण में केवल जीवित कोशिकाएँ होती हैं। द्वितीयक और तृतीयक अध्यावरण - अधिकतर मृत कोशिका भित्तियों से प्राप्त होता है।
पूर्णांक ऊतकों के मुख्य कार्य:
पौधे को सूखने से बचाना;
हानिकारक सूक्ष्मजीवों के प्रवेश से सुरक्षा;
सनबर्न सुरक्षा;
यांत्रिक क्षति से सुरक्षा;
संयंत्र और पर्यावरण के बीच चयापचय का विनियमन;
जलन की धारणा।
प्राथमिक पूर्णांक ऊतक एपिडर्मिस, एपिडर्मिस है। जीवित कोशिकाओं से मिलकर बनता है। शीर्ष विभज्योतक से निर्मित। युवा बढ़ते तनों और पत्तियों को ढकता है।
एपिडर्मिस का गठन पौधों में जलीय आवास से बाहर निकलने के संबंध में किया गया था ताकि इसे सूखने से रोका जा सके। स्टोमेटा के अलावा, एपिडर्मिस की सभी कोशिकाएं कसकर आपस में जुड़ी होती हैं। मुख्य कोशिकाओं की बाहरी दीवारें बाकी की तुलना में मोटी होती हैं। पूरी सतह क्यूटिन और वेजिटेबल वैक्स की एक परत से ढकी होती है। इस परत को क्यूटिकल (त्वचा) कहते हैं। यह पौधों की बढ़ती जड़ों और पानी के नीचे के हिस्सों पर अनुपस्थित है। सूखने पर छल्ली की पारगम्यता काफी कमजोर हो जाती है।
मुख्य कोशिकाओं के अलावा, एपिडर्मिस में अन्य भी होते हैं, विशेष रूप से बाल या ट्राइकोम्स में। वे एककोशिकीय और बहुकोशिकीय हैं (अंक 2)।कार्यात्मक रूप से, वे एपिडर्मिस की सतह को बढ़ाते हैं, उदाहरण के लिए, जड़ विकास क्षेत्र में, यांत्रिक सुरक्षा के रूप में कार्य करते हैं, एक समर्थन से चिपकते हैं, और पानी के नुकसान को कम करते हैं। कई पौधों में ग्रंथियों के बाल होते हैं, जैसे बिछुआ।
केवल उच्च पौधों में एपिडर्मिस में रंध्र होते हैं, जो पानी और गैसों के आदान-प्रदान को नियंत्रित करते हैं। यदि छल्ली नहीं है, तो रंध्रों की कोई आवश्यकता नहीं है। स्टोमेटा कोशिकाओं का एक समूह है जो रंध्र तंत्र का निर्माण करता है, जिसमें दो अनुगामी कोशिकाएँ और आस-पास के एपिडर्मल कोशिकाएँ - पार्श्व कोशिकाएँ होती हैं। वे मुख्य एपिडर्मल कोशिकाओं से भिन्न होते हैं (चित्र 3). गार्ड कोशिकाएं अपने आसपास की कोशिकाओं से आकार और बड़ी संख्या में क्लोरोप्लास्ट और असमान रूप से मोटी दीवारों की उपस्थिति से भिन्न होती हैं। जो एक दूसरे का सामना करते हैं वे बाकी की तुलना में मोटे होते हैं (अंजीर.4). गार्ड कोशिकाओं के बीच, एक रंध्र अंतर बनता है, जो एक सबस्टोमेटल स्पेस की ओर जाता है जिसे सबस्टोमेटल कैविटी कहा जाता है। गार्ड कोशिकाएं अत्यधिक प्रकाश संश्लेषक होती हैं। उनमें बड़ी मात्रा में आरक्षित स्टार्च और कई माइटोकॉन्ड्रिया होते हैं।
रंध्रों की संख्या और वितरण, रंध्र तंत्र के प्रकार विभिन्न पौधों में व्यापक रूप से भिन्न होते हैं। आधुनिक ब्रायोफाइट्स में रंध्र अनुपस्थित होते हैं। उनमें प्रकाश संश्लेषण गैमेटोफाइट पीढ़ी द्वारा किया जाता है, और स्पोरोफाइट स्वतंत्र अस्तित्व में सक्षम नहीं होते हैं।
आमतौर पर रंध्र पत्ती के नीचे की तरफ स्थित होते हैं। पानी की सतह पर तैरने वाले पौधों में - ऊपरी सतह पर। अनाज की पत्तियों में, रंध्र अक्सर दोनों तरफ समान रूप से स्थित होते हैं। ऐसी पत्तियाँ अपेक्षाकृत समान रूप से प्रकाशित होती हैं। सतह के 1 मिमी 2 पर, 100 से 700 रंध्र स्थित हो सकते हैं।
द्वितीयक पूर्णांक ऊतक (पेरिडर्म)। जब वार्षिक प्ररोहों का हरा रंग भूरा हो जाता है तो यह ऊतक एपिडर्मिस की जगह ले लेता है। यह बहुस्तरीय है और कैम्बियल कोशिकाओं की एक केंद्रीय परत होती है - फेलोजेन। फेलोजेन कोशिकाएं, विभाजित होकर, बाहर की तरफ फेलेम की एक परत और अंदर की तरफ फेलोडर्म बिछाती हैं। (चित्र 5)।
फेलिमा, या कॉर्क। सबसे पहले इसमें जीवित पतली दीवार वाली कोशिकाएँ होती हैं। समय के साथ, उनकी दीवारें साबरिन और वनस्पति मोम से संतृप्त हो जाती हैं और मर जाती हैं। सेल की सामग्री हवा से भरी हुई है।
फेलिमा कार्य:
नमी के नुकसान को रोकता है;
पौधे को यांत्रिक क्षति से बचाता है;
रोगजनकों से बचाता है;
थर्मल इन्सुलेशन प्रदान करता है, क्योंकि कोशिकाएं हवा से भरी होती हैं।
एपिडर्मिस में स्थित फेलोजेन की कोशिकाएं, अंतर्निहित सबपीडर्मल परत, प्राथमिक प्रांतस्था की गहरी परतों में अक्सर कम होती हैं, प्राथमिक प्रांतस्था का निर्माण आधार होती हैं।
कॉर्क परत स्थायी नहीं है। इसमें ब्रेक होते हैं, जो पास में स्थित इंटरसेलुलर स्पेस के साथ संचार करते हैं। इसी समय, सतह पर छोटे ट्यूबरकल बनते हैं - मसूर, जो वायुमंडलीय हवा के साथ अंतरकोशिकीय स्थानों के रिक्त स्थान का संचार करते हैं। (चित्र 6,7)।
शरद ऋतु में, मसूर के नीचे फेलोजेन कॉर्क कोशिकाओं की एक परत देता है, जो वाष्पोत्सर्जन को बहुत कम करता है, लेकिन इसे पूरी तरह से बाहर नहीं करता है। वसंत ऋतु में यह परत अंदर से ढह जाती है। हल्की बर्च की छाल पर, काले धब्बे के रूप में दाल स्पष्ट रूप से दिखाई देती है।
तृतीयक पूर्णांक ऊतक (क्रस्ट) भी केवल पौधों के लकड़ी के रूपों की विशेषता है।
कॉर्टेक्स की गहरी परतों में फेलोजेन को बार-बार रखा जाता है। इसके बाहर के ऊतक अंततः मर जाते हैं, एक पपड़ी बनाते हैं। इसकी कोशिकाएँ मर चुकी होती हैं और फैलने में असमर्थ होती हैं। हालांकि, जीवित कोशिकाएं गहरे विभाजन में स्थित होती हैं, जिससे तने के अनुप्रस्थ आकार में वृद्धि होती है। समय के साथ, परत की बाहरी परत टूट जाती है। इस तरह के अंतराल की घटना का समय व्यक्तिगत पौधों के लिए काफी स्थिर मूल्य है। सेब के पेड़ में, यह जीवन के सातवें वर्ष में होता है, हॉर्नबीम में - पचासवें में। कुछ प्रजातियों में यह बिल्कुल नहीं होता है। छीलने का मुख्य कार्य यांत्रिक और थर्मल क्षति से सुरक्षा है।
पैरेन्काइमा
यह विशिष्ट ऊतकों का एक समूह है जो प्रवाहकीय और यांत्रिक ऊतकों के बीच पौधे के शरीर के अंदर रिक्त स्थान को भरता है। (चित्र 8)।अधिक बार पैरेन्काइमा कोशिकाओं का एक गोल आकार होता है, कम अक्सर एक लम्बी आकृति। विकसित अंतरकोशिकीय स्थानों की उपस्थिति विशेषता है। कोशिकाओं के बीच के रिक्त स्थान एक साथ एक परिवहन प्रणाली - एपोप्लास्ट बनाते हैं। इसके अलावा, इंटरसेलुलर रिक्त स्थान पौधे की "वेंटिलेशन सिस्टम" बनाते हैं। रंध्रों, या मसूर के माध्यम से, वे वायुमंडलीय हवा से जुड़े होते हैं और पौधे के अंदर एक इष्टतम गैस संरचना प्रदान करते हैं। जलभराव वाली मिट्टी पर उगने वाले पौधों के लिए विकसित अंतरकोशिकीय स्थान विशेष रूप से आवश्यक होते हैं, जहां सामान्य गैस विनिमय मुश्किल होता है। इस पैरेन्काइमा को एरेन्काइमा कहा जाता है। (चित्र 9)।
पैरेन्काइमा के तत्व, अन्य ऊतकों के बीच अंतराल को भरते हुए, एक समर्थन का कार्य भी करते हैं। पैरेन्काइमा कोशिकाएं जीवित होती हैं, उनमें स्क्लेरेन्काइमा की तरह मोटी कोशिका भित्ति नहीं होती है। इसलिए, यांत्रिक गुण टर्गर द्वारा प्रदान किए जाते हैं। यदि पानी की मात्रा कम हो जाती है, जिससे पौधे का प्लास्मोलिसिस और विल्टिंग हो जाता है।
एसिमिलेशन पैरेन्काइमा पतली दीवार वाली कोशिकाओं द्वारा कई अंतरकोशिकीय स्थानों के साथ बनता है। इस संरचना की कोशिकाओं में बहुत सारे क्लोरोप्लास्ट होते हैं, इसलिए इसे क्लोरेन्काइमा कहा जाता है। क्लोरोप्लास्ट एक दूसरे को छायांकित किए बिना दीवार के साथ स्थित होते हैं। एसिमिलेशन पैरेन्काइमा में, प्रकाश संश्लेषण प्रतिक्रियाएं होती हैं, जो पौधे को कार्बनिक पदार्थ और ऊर्जा प्रदान करती हैं। प्रकाश संश्लेषक प्रक्रियाओं का परिणाम पृथ्वी पर सभी जीवित जीवों के अस्तित्व की संभावना है।
आत्मसात ऊतक केवल पौधे के प्रबुद्ध भागों में मौजूद होते हैं, वे एक पारदर्शी एपिडर्मिस द्वारा पर्यावरण से अलग होते हैं। यदि एपिडर्मिस को अपारदर्शी द्वितीयक पूर्णांक ऊतकों द्वारा प्रतिस्थापित किया जाता है, तो एसिमिलेशन पैरेन्काइमा गायब हो जाता है।
भंडारण पैरेन्काइमा कार्बनिक पदार्थों के लिए एक पात्र के रूप में कार्य करता है जो अस्थायी रूप से पौधे जीव द्वारा उपयोग नहीं किया जाता है। सिद्धांत रूप में, जीवित प्रोटोप्लास्ट वाली कोई भी कोशिका विभिन्न प्रकार के समावेशन के रूप में कार्बनिक पदार्थ जमा करने में सक्षम है, लेकिन कुछ कोशिकाएँ इसमें विशेषज्ञ होती हैं। (चित्र 10, 11)।ऊर्जा से भरपूर यौगिकों को केवल बढ़ते मौसम के दौरान जमा किया जाता है, सुप्त अवधि के दौरान और अगले बढ़ते मौसम की तैयारी में खपत होती है। इसलिए, आरक्षित पदार्थ केवल बारहमासी पौधों में वनस्पति अंगों में जमा होते हैं।
शेयरों का भंडार सामान्य अंग (पलायन, जड़), साथ ही विशेष वाले (राइजोम, कंद, बल्ब) हो सकते हैं। सभी बीज पौधे बीजों (बीजपत्र, एंडोस्पर्म) में ऊर्जावान रूप से मूल्यवान पदार्थों को संग्रहित करते हैं। शुष्क जलवायु में कई पौधे न केवल कार्बनिक पदार्थ, बल्कि पानी भी जमा करते हैं। (चित्र 12)।उदाहरण के लिए, मुसब्बर मांसल पत्तियों में पानी जमा करता है, शूटिंग में कैक्टि।
यांत्रिक कपड़े
पादप कोशिकाओं के यांत्रिक गुणों द्वारा प्रदान किया जाता है:
कोशिका का कठोर खोल;
स्फीति, अर्थात् कोशिकाओं की स्फीति अवस्था।
इस तथ्य के बावजूद कि लगभग सभी ऊतक कोशिकाओं में यांत्रिक गुण होते हैं, हालांकि, पौधे में ऐसे ऊतक होते हैं जिनके लिए यांत्रिक गुण बुनियादी होते हैं। ये कॉलेन्काइमा और स्क्लेरेन्काइमा हैं। वे आमतौर पर अन्य ऊतकों के साथ बातचीत में कार्य करते हैं। शरीर के अंदर पौधे एक प्रकार का ढाँचा बनाते हैं। इसलिए, उन्हें सुदृढीकरण कहा जाता है।
सभी पौधों में समान रूप से स्पष्ट यांत्रिक ऊतक नहीं होते हैं। जलीय वातावरण में रहने वाले पौधों को स्थलीय की तुलना में बहुत कम आंतरिक समर्थन की आवश्यकता होती है। इसका कारण यह है कि जलीय पौधों को कम आंतरिक समर्थन की आवश्यकता होती है। उनका शरीर काफी हद तक आसपास के पानी से समर्थित है। जमीन पर हवा इस तरह का समर्थन नहीं बनाती है, क्योंकि इसमें पानी की तुलना में कम घनत्व होता है। यही कारण है कि विशिष्ट यांत्रिक ऊतकों की उपस्थिति प्रासंगिक हो जाती है।
विकास की प्रक्रिया में आंतरिक समर्थन संरचनाओं में सुधार हुआ।
Collenchyma। यह केवल जीवित कोशिकाओं द्वारा बनता है, जो अंग की धुरी के साथ लम्बी होती हैं। प्राथमिक वृद्धि की अवधि के दौरान इस प्रकार के यांत्रिक ऊतक बहुत जल्दी बनते हैं। इसलिए, यह महत्वपूर्ण है कि कोशिकाएं जीवित रहें और पास की खींची हुई कोशिकाओं के साथ-साथ फैलने की क्षमता बनाए रखें।
कोलेन्काइमा कोशिकाओं की विशेषताएं:
खोल का असमान मोटा होना, जिसके परिणामस्वरूप इसके कुछ हिस्से पतले रहते हैं, जबकि अन्य मोटे होते हैं;
गोले लिग्निफाई नहीं करते हैं।
Collenchyma कोशिकाएं एक दूसरे के सापेक्ष अलग-अलग स्थित होती हैं। आसन्न कोशिकाओं में, एक दूसरे के सामने वाले कोनों पर गाढ़ापन बनता है ( चावल। 13). ऐसे कोलेन्काइमा को कॉर्नर कॉलेन्काइमा कहा जाता है। दूसरे मामले में, कोशिकाओं को समानांतर परतों में व्यवस्थित किया जाता है। इन परतों का सामना करने वाली कोशिका झिल्लियाँ अत्यधिक मोटी होती हैं। यह लैमेलर कोलेंकाइमा है। प्रचुर मात्रा में अंतरकोशिकीय रिक्त स्थान के साथ कोशिकाएं शिथिल रूप से स्थित हो सकती हैं - यह एक ढीला कोलेन्काइमा है। इस तरह के कोलेंकाइमा अक्सर जलभराव वाली मिट्टी पर पौधों में पाए जाते हैं।
Collenchyma युवा पौधों, जड़ी-बूटियों के रूपों, और पौधों के उन हिस्सों में भी विशेष महत्व रखता है जहाँ द्वितीयक वृद्धि नहीं होती है, जैसे कि पत्तियाँ। इस मामले में, इसे सतह के बहुत करीब रखा जाता है, कभी-कभी सिर्फ एपिडर्मिस के नीचे। यदि अंग के चेहरे हैं, तो उनके शिखर के साथ कोलेन्काइमा की मोटी परतें पाई जाती हैं।
Collenchyma कोशिकाएं केवल स्फीति की उपस्थिति में क्रियाशील होती हैं। पानी की कमी से कॉलेंकाइमा की प्रभावशीलता कम हो जाती है और पौधा अस्थायी रूप से मुरझा जाता है, उदाहरण के लिए, ककड़ी के पत्ते गर्म दिन में गिर जाते हैं। कोशिकाओं को पानी से भरने के बाद, कोलेंकाइमा के कार्य बहाल हो जाते हैं।
दृढ़ोतक। दूसरे प्रकार के यांत्रिक ऊतक। Collenchyma के विपरीत, जहाँ सभी कोशिकाएँ जीवित हैं, स्क्लेरेन्काइमा कोशिकाएँ मृत हैं। इनकी दीवारें बहुत मोटी होती हैं। वे एक यांत्रिक कार्य करते हैं। झिल्ली का एक मजबूत मोटा होना पदार्थों के परिवहन का उल्लंघन करता है, जिसके परिणामस्वरूप प्रोटोप्लास्ट मर जाता है ( चावल। 14). स्क्लेरेन्काइमा की कोशिका झिल्लियों का लिग्निफिकेशन तब होता है जब पौधे का अंग पहले ही अपनी वृद्धि पूरी कर चुका होता है। इसलिए, वे अब आसपास के ऊतकों के खिंचाव में हस्तक्षेप नहीं करते हैं।
आकार के आधार पर, दो प्रकार की स्क्लेरेन्काइमा कोशिकाएं प्रतिष्ठित होती हैं - तंतु और स्केलेरिड।
तंतुओं में बहुत मोटी दीवारों और एक छोटी गुहा के साथ एक मजबूत लम्बी आकृति होती है। वे लकड़ी के तंतुओं से कुछ छोटे होते हैं। अक्सर एपिडर्मिस के नीचे अनुदैर्ध्य परतें और किस्में बनती हैं। फ्लोएम या जाइलम में ये अकेले या समूहों में पाए जाते हैं। फ्लोएम में उन्हें बास्ट फाइबर कहा जाता है ( चावल। 15), और जाइलम में - लाइब्रिफॉर्म फाइबर ( चावल। 16).
स्केलेरिड्स, या स्टोनी कोशिकाएं, शक्तिशाली झिल्लियों के साथ गोल या शाखित कोशिकाओं द्वारा दर्शाई जाती हैं। एक पौधे के शरीर में, वे अकेले (सहायक कोशिकाओं) में पाए जा सकते हैं ( अंजीर.17) या समूह (चित्र 18)। यह ध्यान दिया जाना चाहिए कि यांत्रिक गुण दृढ़ता से स्केलेरिड्स के स्थान पर निर्भर करते हैं। स्क्लेराइड्स का हिस्सा निरंतर परतें बनाता है, उदाहरण के लिए, नट के खोल में या फलों के पत्थरों (पत्थर के फल) में।
उत्सर्जक ऊतक
किसी भी जीवित कोशिका में प्रारंभ में पदार्थों को हटाने का कार्य होता है, जबकि परिवहन सांद्रता प्रवणता के साथ और ढाल के विरुद्ध दोनों जा सकता है। पदार्थ जो कोशिका में संश्लेषित किए गए हैं उन्हें हटाया जा सकता है और बाद में अन्य कोशिकाओं (फाइटोहोर्मोन, एंजाइम) की गतिविधि को प्रभावित करेगा। इस प्रक्रिया को स्राव कहते हैं। मामले में जब पदार्थ जो कोशिका के अपशिष्ट उत्पाद हैं, उत्सर्जित होते हैं, इस प्रक्रिया को उत्सर्जन कहा जाता है। इस तथ्य के बावजूद कि पौधों और जानवरों की कोशिकाओं में पदार्थों के उत्सर्जन में मौलिक समानता है, पौधों में कई विशेषताएं हैं जो महत्वपूर्ण गतिविधि में मौलिक अंतर से उत्पन्न होती हैं। पौधों में चयापचय प्रतिक्रियाओं का स्तर जानवरों की तुलना में बहुत कम है। इसलिए, आनुपातिक रूप से कम मात्रा में कचरा उत्सर्जित होता है। एक अन्य विशेषता यह है कि, लगभग सभी आवश्यक कार्बनिक यौगिकों को स्वतंत्र रूप से संश्लेषित करते हुए, पौधे कभी भी उनका अत्यधिक भंडार नहीं बनाते हैं। जारी किए गए पदार्थ अन्य प्रतिक्रियाओं के लिए प्रारंभिक सामग्री के रूप में काम कर सकते हैं (उदाहरण के लिए, CO2 और H2O)।
यदि जानवरों में विषाक्त पदार्थों के उत्सर्जन की प्रक्रिया शरीर से उनके निष्कासन से जुड़ी होती है, तो पौधे ऐसा नहीं कर सकते हैं, जीवित प्रोटोप्लास्ट में अनावश्यक पदार्थों को अलग करना (वैक्यूल में विभिन्न पदार्थों को हटाना), मृत कोशिकाओं में (एक बारहमासी के अधिकांश ऊतक) पौधे में ऐसी कोशिकाएँ होती हैं), अंतरकोशिकीय स्थानों में।
उत्सर्जित पदार्थों को दो बड़े समूहों में विभाजित किया जा सकता है। पहला समूह जैविक पदार्थ है जो सीधे कोशिका द्वारा संश्लेषित होता है (एंजाइम, पॉलीसेकेराइड, लिग्निन, टेरपेन, बाद वाले आवश्यक तेलों और रेजिन के घटक तत्व हैं)। दूसरा समूह - पदार्थ जो शुरू में जाइलम या फ्लोएम परिवहन (पानी, खनिज लवण, अमीनो एसिड, मोनोसैकराइड, आदि) की मदद से बाहर से कोशिका में प्रवेश करते हैं। उत्सर्जित रहस्य शायद ही कभी सजातीय होता है और आमतौर पर एक मिश्रण होता है जिसमें एक पदार्थ में उच्चतम एकाग्रता होती है।
जंतुओं के विपरीत पौधों में पूर्ण उत्सर्जन तंत्र नहीं होता है। पूरे संयंत्र में बिखरी हुई केवल विशेष संरचनाएं हैं - इडियोब्लास्ट्स। उन्हें बनाने वाली कोशिकाएँ पास में पड़ी पैरेन्काइमा कोशिकाओं से छोटी होती हैं। उनके पास एक इलेक्ट्रॉन-सघन साइटोप्लाज्म है, जिसमें एंडोप्लाज्मिक रेटिकुलम और गोल्गी कॉम्प्लेक्स के विकसित तत्व हैं। अक्सर व्यक्त नहीं किया जाता है, एक केंद्रीय रिक्तिका होती है। अपने और अन्य जीवित कोशिकाओं के बीच, ये कोशिकाएँ असंख्य प्लास्मोडेस्माटा द्वारा जुड़ी होती हैं।
स्थान के आधार पर, उत्सर्जक ऊतक बाहरी और आंतरिक हो सकते हैं।
बाहरी उत्सर्जक ऊतक।ग्रंथियों के बाल एपिडर्मिस के डेरिवेटिव हैं। Morphologically, वे परिवर्तनशील हैं - उनके पास एक बहुकोशिकीय सिर हो सकता है, लम्बी हो सकती है, एक पैर पर ढाल के रूप में, आदि। (अंजीर। 19)। इनमें चुभने वाले बिछुआ बाल शामिल हैं।
हाइडेथोड ऐसी संरचनाएं हैं जो कम वाष्पोत्सर्जन और उच्च आर्द्रता - गट्टेशन की स्थितियों में अतिरिक्त पानी छोड़ती हैं। ये संरचनाएं रंगहीन जीवित कोशिकाओं के समूहों द्वारा पतली दीवारों के साथ बनाई जाती हैं - एपिथेम। यह ऊतक प्रवाहकीय बंडल (चित्र 20) के निकट है। पानी को विशेष जल रंध्रों के माध्यम से छोड़ा जाता है, जो सामान्य रंध्रों से गतिहीनता और लगातार खुले अंतराल से भिन्न होते हैं। गुटेशन द्रव की संरचना लगभग शुद्ध पानी से लेकर पदार्थों के बहुत जटिल मिश्रण तक व्यापक रूप से भिन्न होती है।
अमृत। कई पौधों को 7 से 87% मोनो- और ओलिगोसेकेराइड युक्त तरल की रिहाई की विशेषता है। यह प्रक्रिया विशेष संरचनाओं - अमृत द्वारा की जाती है। स्थान के आधार पर, फूल, या पुष्प, अमृत प्रतिष्ठित हैं (चित्र 21), साथ ही तनों और पत्तियों पर स्थित पौधे - अतिरिक्त पुष्प, या अतिरिक्त अमृत। मकरंद अपरिवर्तित फ्लोएम रस हो सकता है, जो अंतरकोशिकीय स्थानों के माध्यम से सतह पर पहुंचाया जाता है और रंध्रों के माध्यम से उत्सर्जित होता है। ग्रंथियों के पैरेन्काइमा द्वारा अधिक जटिल अमृत बनते हैं, जो ग्रंथियों के बालों के साथ एपिडर्मिस से ढके होते हैं। अमृत या तो एपिडर्मल कोशिकाओं या ग्रंथियों के बालों द्वारा उत्सर्जित होता है। इस मामले में, स्रावित अमृत फ्लोएम सैप से भिन्न होता है। चूंकि इसमें ग्लूकोज, फ्रुक्टोज, सुक्रोज और फ्लोएम जूस - ग्लूकोज का प्रभुत्व है। आयन कम सांद्रता में मौजूद हैं। आयन कम सांद्रता में मौजूद हैं। परागणकों को आकर्षित करने के लिए अमृत में स्टेरॉयड हार्मोन हो सकते हैं, जो कीड़ों के लिए आवश्यक हैं।
कीटभक्षी पौधों (सूंड्यू, नेपेंथिस, आदि) में पाचन ग्रंथियां मौजूद होती हैं। छोटे अकशेरूकीय के स्पर्श के जवाब में, वे हाइड्रोलाइटिक एंजाइम, फॉर्मिक, ब्यूटिरिक और प्रोपियोनिक एसिड युक्त एक रहस्य का स्राव करते हैं। नतीजतन, बायोपॉलिमर्स मोनोमर्स में टूट जाते हैं, जिनका उपयोग पौधे अपनी जरूरतों के लिए करते हैं।
लवणीय मिट्टी में उगने वाले पौधों में लवण ग्रंथियाँ विकसित होती हैं। ये ग्रंथियां पत्तियों पर स्थित होती हैं और घुले हुए रूप में, अतिरिक्त आयन सतह पर लाए जाते हैं। नमक पहले छल्ली पर जमा होता है, फिर इसे बारिश से धोया जाता है (चित्र 22)। इसके अलावा, कई पौधों की पत्तियों पर नमक के बाल होते हैं। प्रत्येक बाल में दो कोशिकाएँ होती हैं: एक सिर बनाती है, दूसरी - तना। सिर की कोशिका की रसधानी में लवण जमा हो जाते हैं, और जब उनकी सघनता अधिक हो जाती है, तो सिर गिर जाता है। इसके स्थान पर एक नई कोशिका विकसित होती है, जो लवण भी जमा करती है। पत्ती के विकास के दौरान, सिर गिर सकता है और कई बार बन सकता है। नमक बालों का लाभ यह है कि नमक छोड़ने से वे थोड़ा पानी खो देते हैं। खारी मिट्टी पर उगने वाले पौधों के लिए यह घटना बहुत महत्वपूर्ण है, जहां ताजे पानी की कमी है।
आंतरिक उत्सर्जक ऊतक।इडियोब्लास्ट्स के रूप में पौधे के पूरे शरीर में बिखरी हुई आंतरिक उत्सर्जन संरचनाएं, शरीर के बाहर पदार्थों को नहीं हटाती हैं, उन्हें स्वयं में जमा करती हैं। इडियोब्लास्ट पादप कोशिकाएं हैं जो एक ही ऊतक में अन्य कोशिकाओं से आकार, संरचना या सामग्री में भिन्न होती हैं। उत्पत्ति के आधार पर, स्रावित पदार्थों के स्किज़ोजेनिक (ग्रीक स्किसो - स्प्लिट) और लाइसोजेनिक (ग्रीक लिसीज़ - विघटन) रिसेप्टेकल्स को प्रतिष्ठित किया जाता है (चित्र 23)।
स्किज़ोजेनिक रिसेप्टेकल्स। ये कमोबेश व्यापक अंतरकोशिकीय स्थान हैं जो उत्सर्जित पदार्थों से भरे हुए हैं। उदाहरण के लिए, कोनिफर्स, अरालिया, छाता और अन्य के राल मार्ग। यह माना जाता है कि राल में जीवाणुनाशक गुण होते हैं, शाकाहारी जीवों को पीछे हटाते हैं, जिससे पौधे उनके लिए अखाद्य हो जाते हैं।
लाइसजेनिक रिसेप्टेकल्स जीवित कोशिकाओं के स्थान पर बनते हैं, जो मर जाते हैं और उनमें पदार्थों के जमा होने के बाद नष्ट हो जाते हैं। छिलके में इस तरह के गठन देखे जा सकते हैं
प्लांट सेल की संरचना
पादप कोशिका कमोबेश कठोर होती है कोशिका भित्तिऔर प्रोटोप्लास्ट।कोशिका भित्ति कोशिका भित्ति और साइटोप्लाज्मिक झिल्ली है। प्रोटोप्लास्ट शब्द शब्द से आया है जीवद्रव्य,जो लंबे समय से सभी जीवित चीजों को संदर्भित करने के लिए उपयोग किया जाता है। प्रोटोप्लास्ट एक व्यक्तिगत कोशिका का प्रोटोप्लाज्म है।
प्रोटोप्लास्ट बना होता है कोशिका द्रव्यऔर गुठली।साइटोप्लाज्म में ऑर्गेनेल (राइबोसोम, माइक्रोट्यूबुल्स, प्लास्टिड्स, माइटोकॉन्ड्रिया) और मेम्ब्रेन सिस्टम (एंडोप्लाज्मिक रेटिकुलम, डिक्टीयोसोम) होते हैं। साइटोप्लाज्म में साइटोप्लाज्मिक मैट्रिक्स भी शामिल है ( वास्तविक पदार्थ) जिसमें ऑर्गेनेल और मेम्ब्रेन सिस्टम डूबे हुए हैं। साइटोप्लाज्म को कोशिका भित्ति से अलग किया जाता है प्लाज्मा झिल्ली, जो एक प्राथमिक झिल्ली है। अधिकांश पशु कोशिकाओं के विपरीत, पादप कोशिकाओं में एक या अधिक होते हैं रिक्तिकाएं. ये द्रव से भरे पुटिका होते हैं जो एक प्रारंभिक झिल्ली से घिरे होते हैं ( टोनोप्लास्ट)।
एक जीवित पादप कोशिका में, जमीनी पदार्थ निरंतर गति में रहता है। साइटोप्लाज्मिक करंट या नामक एक आंदोलन में चक्रवात,ऑर्गेनेल शामिल हैं। साइक्लोसिस कोशिका में पदार्थों की आवाजाही और कोशिका और पर्यावरण के बीच उनके आदान-प्रदान की सुविधा प्रदान करता है।
प्लाज्मा झिल्ली।यह एक बाइलेयर फॉस्फोलिपिड संरचना है। पादप कोशिकाओं की विशेषता प्लाज्मा झिल्ली के अंतर्वलन से होती है।
प्लाज्मा झिल्ली निम्नलिखित कार्य करती है:
कोशिका और पर्यावरण के बीच पदार्थों के आदान-प्रदान में भाग लेता है;
कोशिका भित्ति के सेल्युलोज सूक्ष्मतंतुओं के संश्लेषण और संयोजन का समन्वय करता है;
हार्मोनल और बाहरी संकेतों को प्रसारित करता है जो सेल के विकास और भेदभाव को नियंत्रित करते हैं।
मुख्य।यह यूकेरियोटिक कोशिका के साइटोप्लाज्म में सबसे प्रमुख संरचना है। कर्नेल दो महत्वपूर्ण कार्य करता है:
कोशिका की महत्वपूर्ण गतिविधि को नियंत्रित करता है, यह निर्धारित करता है कि कौन से प्रोटीन और किस समय संश्लेषित किए जाने चाहिए;
आनुवंशिक जानकारी को संग्रहीत करता है और इसे कोशिका विभाजन के दौरान संतति कोशिकाओं तक पहुंचाता है।
यूकेरियोटिक कोशिका का केंद्रक दो प्रारंभिक झिल्लियों से घिरा होता है जो बनती हैं परमाणु लिफाफा।यह 30 से 100 एनएम के व्यास के साथ कई छिद्रों से व्याप्त है, जो केवल एक इलेक्ट्रॉन माइक्रोस्कोप में दिखाई देता है। छिद्रों की एक जटिल संरचना होती है। कुछ स्थानों पर परमाणु झिल्ली की बाहरी झिल्ली को एंडोप्लाज्मिक रेटिकुलम के साथ जोड़ा जाता है। परमाणु लिफाफे को एंडोप्लाज्मिक रेटिकुलम (ईआर) के एक विशेष, स्थानीय रूप से विभेदित भाग के रूप में देखा जा सकता है।
विशेष रंगों से रंगे हुए कोर में, पतले धागों और गांठों को प्रतिष्ठित किया जा सकता है क्रोमेटिनऔर न्यूक्लियोप्लाज्म(नाभिक का मुख्य पदार्थ)। क्रोमेटिन हिस्टोन नामक विशेष प्रोटीन से बंधे डीएनए से बना होता है। कोशिका विभाजन की प्रक्रिया में, क्रोमैटिन अधिक से अधिक सघन हो जाता है और इसमें इकट्ठा हो जाता है गुणसूत्र।डीएनए आनुवंशिक जानकारी को एनकोड करता है।
दैहिक कोशिकाओं में जीव गुणसूत्रों की संख्या में भिन्न होते हैं। उदाहरण के लिए, गोभी में - 20 गुणसूत्र होते हैं; सूरजमुखी - 34; गेहूँ - 42; लोग - 46, और फर्न की प्रजातियों में से एक ओफियोग्लोसम – 1250. सेक्स कोशिकाओं (युग्मक) में शरीर की दैहिक कोशिकाओं की विशेषता वाले गुणसूत्रों की संख्या का केवल आधा हिस्सा होता है। युग्मकों में गुणसूत्रों की संख्या कहलाती है अगुणित(एकल), दैहिक कोशिकाओं में - द्विगुणित(दोहरा)। गुणसूत्रों के दो से अधिक सेट वाली कोशिकाओं को कहा जाता है बहुगुणित.
एक प्रकाश सूक्ष्मदर्शी से गोलाकार संरचनाओं को देखा जा सकता है - उपकेन्द्रक. प्रत्येक नाभिक में एक या एक से अधिक नाभिक होते हैं, जो अविभाजित नाभिक में दिखाई देते हैं। राइबोसोमल आरएनए को न्यूक्लियोलस में संश्लेषित किया जाता है। आमतौर पर, द्विगुणित जीवों के नाभिक में दो नाभिक होते हैं, गुणसूत्रों के प्रत्येक अगुणित सेट के लिए एक। न्यूक्लियोली की अपनी झिल्ली नहीं होती है। बायोकेमिकल रूप से, न्यूक्लियोली को आरएनए की उच्च सांद्रता की विशेषता होती है, जो यहां फॉस्फोप्रोटीन के साथ जुड़ा हुआ है। नाभिक का आकार कोशिका की कार्यात्मक अवस्था पर निर्भर करता है। यह देखा गया है कि एक तेजी से बढ़ने वाली कोशिका में, जिसमें प्रोटीन संश्लेषण की गहन प्रक्रियाएँ हो रही होती हैं, नाभिक का आकार बढ़ जाता है। न्यूक्लियोली में, एमआरएनए और राइबोसोम उत्पन्न होते हैं, जो केवल न्यूक्लियस में सिंथेटिक कार्य करते हैं।
न्यूक्लियोप्लाज्म (कार्योप्लाज्म) एक सजातीय तरल द्वारा दर्शाया जाता है जिसमें एंजाइम सहित विभिन्न प्रोटीन घुल जाते हैं।
प्लास्टिड्स।रिक्तिकाएं, सेल्युलोज कोशिका भित्ति और प्लास्टिडपादप कोशिकाओं के विशिष्ट घटक हैं। प्रत्येक प्लास्टिड का अपना खोल होता है, जिसमें दो प्राथमिक झिल्लियाँ होती हैं। प्लास्टिड के अंदर, एक झिल्ली प्रणाली और सजातीय पदार्थ की एक अलग डिग्री प्रतिष्ठित होती है - स्ट्रोमा।परिपक्व प्लास्टिड्स को उनमें मौजूद पिगमेंट के आधार पर वर्गीकृत किया जाता है।
क्लोरोप्लास्ट,जिसमें प्रकाश संश्लेषण होता है, इसमें क्लोरोफिल और कैरोटीनॉयड होते हैं। आमतौर पर 4 - 5 माइक्रोन के व्यास वाली डिस्क का रूप होता है। मेसोफिल (पत्ती के मध्य) की एक कोशिका में 40-50 क्लोरोप्लास्ट हो सकते हैं; मिमी 2 शीट्स में - लगभग 500,000। साइटोप्लाज्म में, क्लोरोप्लास्ट आमतौर पर कोशिका झिल्ली के समानांतर स्थित होते हैं।
क्लोरोप्लास्ट की आंतरिक संरचना जटिल है। स्ट्रोमा को बुलबुले के रूप में झिल्लियों की एक विकसित प्रणाली द्वारा अनुमति दी जाती है - थायलाकोइड्स।प्रत्येक थायलाकोइड में दो झिल्ली होते हैं। थायलाकोइड्स एकल प्रणाली बनाते हैं। एक नियम के रूप में, उन्हें ढेर में एकत्र किया जाता है - अनाजसिक्कों के स्तम्भों के समान। अलग-अलग ग्रैने के थायलाकोइड्स स्ट्रोमा थायलाकोइड्स, या इंटरग्रेनल थायलाकोइड्स द्वारा परस्पर जुड़े होते हैं। क्लोरोफिल और कैरोटीनॉयड थायलाकोइड झिल्ली में एम्बेडेड होते हैं। हरे पौधों और शैवाल में क्लोरोप्लास्ट में अक्सर स्टार्च के दाने और छोटे लिपिड (वसा) की बूंदें होती हैं। स्टार्च के दाने प्रकाश संश्लेषण उत्पादों के अस्थायी भंडार होते हैं। वे केवल 24 घंटों के लिए अंधेरे में रहने वाले पौधे के क्लोरोप्लास्ट से गायब हो सकते हैं, और पौधे को प्रकाश में स्थानांतरित करने के 3-4 घंटे बाद फिर से प्रकट हो सकते हैं।
क्लोरोप्लास्ट अर्ध-स्वायत्त अंगक होते हैं और बैक्टीरिया के समान होते हैं। उदाहरण के लिए, बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम में काफी उच्च समानता होती है। वे यूकेरियोटिक राइबोसोम से छोटे होते हैं। बैक्टीरिया और क्लोरोप्लास्ट के राइबोसोम पर प्रोटीन संश्लेषण क्लोरैम्फेनिकॉल द्वारा बाधित होता है, जिसका यूकेरियोटिक कोशिकाओं पर कोई प्रभाव नहीं पड़ता है। इसके अलावा, बैक्टीरिया और क्लोरोप्लास्ट दोनों में एक समान प्रकार का न्यूक्लियॉइड होता है, जो समान तरीके से व्यवस्थित होता है। इस तथ्य के बावजूद कि क्लोरोप्लास्ट का निर्माण और उनमें निहित पिगमेंट का संश्लेषण काफी हद तक कोशिका के क्रोमोसोमल डीएनए द्वारा नियंत्रित होता है, फिर भी, अपने स्वयं के डीएनए की अनुपस्थिति में, क्लोरोप्लास्ट नहीं बनते हैं।
क्लोरोप्लास्ट को मुख्य सेलुलर अंग माना जा सकता है, क्योंकि वे सौर ऊर्जा के परिवर्तन की श्रृंखला में पहले हैं, जिसके परिणामस्वरूप मानवता को भोजन और ईंधन दोनों प्राप्त होते हैं। क्लोरोप्लास्ट में न केवल प्रकाश संश्लेषण होता है। वे अमीनो एसिड और फैटी एसिड के संश्लेषण में भी शामिल हैं, स्टार्च के अस्थायी भंडार के भंडार के रूप में काम करते हैं।
क्रोमोप्लास्ट -रंजित प्लास्टिड्स। विविध रूप में, उनके पास क्लोरोफिल नहीं है, लेकिन कैरोटीनॉयड को संश्लेषित और संचित करते हैं, जो फूलों, पुरानी पत्तियों, फलों और जड़ों को पीला, नारंगी, लाल रंग देते हैं। क्रोमोप्लास्ट क्लोरोप्लास्ट से विकसित हो सकते हैं, जो इस मामले में क्लोरोफिल और आंतरिक झिल्ली संरचनाओं को खो देते हैं, कैरोटीनॉयड जमा करते हैं। यह कई फलों के पकने के दौरान होता है। क्रोमोप्लास्ट कीड़ों और अन्य जानवरों को आकर्षित करते हैं जिनके साथ वे विकसित हुए हैं।
ल्युकोप्लास्ट्स -अवर्णित प्लास्टिड्स। उनमें से कुछ स्टार्च को संश्लेषित करते हैं ( एमाइलोप्लास्ट), अन्य लिपिड और प्रोटीन सहित विभिन्न पदार्थ बनाने में सक्षम हैं। प्रकाश में, ल्यूकोप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं।
प्रोप्लास्टिड्स -छोटे रंगहीन या हल्के हरे रंग के अविभाजित प्लास्टिड्स जो जड़ों और टहनियों की मेरिस्टेमेटिक (विभाजित) कोशिकाओं में पाए जाते हैं। वे अन्य, अधिक विभेदित प्लास्टिड्स - क्लोरोप्लास्ट्स, क्रोमोप्लास्ट्स और एमिनोप्लास्ट्स के पूर्ववर्ती हैं। यदि प्रकाश की कमी के कारण प्रोटोप्लास्टिड्स के विकास में देरी हो रही है, तो एक या अधिक प्रोलामेलर निकाय, जो ट्यूबलर झिल्ली के अर्ध-क्रिस्टलीय संचय हैं। प्रोलामेलर बॉडी वाले प्लास्टिड्स कहलाते हैं etioplasts.प्रकाश में, एटिओप्लास्ट क्लोरोप्लास्ट में बदल जाते हैं, जबकि प्रोलामेलर निकायों की झिल्लियां थायलाकोइड्स बनाती हैं। इटियोप्लास्ट पौधों की पत्तियों में बनते हैं जो अंधेरे में होते हैं। बीज भ्रूण के प्रोटोप्लास्ट पहले इटियोप्लास्ट में बदलते हैं, जिससे क्लोरोप्लास्ट प्रकाश में विकसित होते हैं। प्लास्टिड्स को एक प्रकार से दूसरे प्रकार में अपेक्षाकृत आसान संक्रमणों की विशेषता है। प्लास्टिड्स, बैक्टीरिया की तरह, दो में विभाजित करके प्रजनन करते हैं। मेरिस्टेमेटिक कोशिकाओं में, प्रोटोप्लास्टिड्स के विभाजन का समय लगभग कोशिका विभाजन के समय के साथ मेल खाता है। हालांकि, परिपक्व कोशिकाओं में, अधिकांश प्लास्टिड परिपक्व प्लास्टिड्स के विभाजन के परिणामस्वरूप बनते हैं।
माइटोकॉन्ड्रिया।क्लोरोप्लास्ट की तरह, माइटोकॉन्ड्रिया दो प्राथमिक झिल्लियों से घिरा होता है। भीतरी झिल्ली कई तहें और उभार बनाती है - संकट,जो माइटोकॉन्ड्रिया की आंतरिक सतह को महत्वपूर्ण रूप से बढ़ाते हैं। वे प्लास्टिड्स से काफी छोटे हैं, व्यास में लगभग 0.5 माइक्रोन और लंबाई और आकार में परिवर्तनशील हैं।
माइटोकॉन्ड्रिया में, श्वसन की प्रक्रिया होती है, जिसके परिणामस्वरूप कार्बनिक अणु ऊर्जा की रिहाई के साथ टूट जाते हैं और एटीपी अणुओं में स्थानांतरित हो जाते हैं, जो सभी यूकेरियोटिक कोशिकाओं का मुख्य ऊर्जा भंडार है। अधिकांश पादप कोशिकाओं में सैकड़ों या हजारों माइटोकॉन्ड्रिया होते हैं। एक सेल में उनकी संख्या सेल की एटीपी की आवश्यकता से निर्धारित होती है। माइटोकॉन्ड्रिया निरंतर गति में हैं, कोशिका के एक भाग से दूसरे भाग में जाते हैं, एक दूसरे के साथ विलय और विभाजित होते हैं। माइटोकॉन्ड्रिया आमतौर पर वहां इकट्ठा होते हैं जहां ऊर्जा की जरूरत होती है। यदि प्लाज़्मा झिल्ली सक्रिय रूप से कोशिका से कोशिका में पदार्थों को स्थानांतरित करती है, तो माइटोकॉन्ड्रिया झिल्ली की सतह के साथ स्थित होते हैं। मोटिव एककोशिकीय शैवाल में, माइटोकॉन्ड्रिया फ्लैगेल्ला के आधार पर जमा होता है, जो उनके आंदोलन के लिए आवश्यक ऊर्जा की आपूर्ति करता है।
माइटोकॉन्ड्रिया, प्लास्टिड्स की तरह, अर्ध-स्वायत्त अंग होते हैं जिनमें अपने स्वयं के प्रोटीन के संश्लेषण के लिए आवश्यक घटक होते हैं। आंतरिक झिल्ली एक द्रव मैट्रिक्स को घेरती है जिसमें प्रोटीन, आरएनए, डीएनए, बैक्टीरिया जैसे राइबोसोम और विभिन्न विलेय होते हैं। डीएनए एक या एक से अधिक न्यूक्लियॉइड्स में स्थित गोलाकार अणुओं के रूप में मौजूद होता है।
माइटोकॉन्ड्रिया और यूकेरियोटिक कोशिकाओं के क्लोरोप्लास्ट के साथ बैक्टीरिया की समानता के आधार पर, यह माना जा सकता है कि माइटोकॉन्ड्रिया और क्लोरोप्लास्ट बैक्टीरिया से उत्पन्न हुए हैं जो बड़े हेटरोट्रॉफ़िक कोशिकाओं में "शरण" पाए गए - यूकेरियोट्स के अग्रदूत।
माइक्रोबॉडीज।प्लास्टिड्स और माइटोकॉन्ड्रिया के विपरीत, जो दो झिल्लियों द्वारा सीमांकित होते हैं, microbodyएक झिल्ली से घिरे गोलाकार अंगक होते हैं। माइक्रोबॉडी में एक दानेदार (दानेदार) सामग्री होती है, कभी-कभी उनमें क्रिस्टलीय प्रोटीन का समावेश भी होता है। माइक्रोबॉडी एंडोप्लाज्मिक रेटिकुलम के एक या दो क्षेत्रों से जुड़े होते हैं।
कुछ माइक्रोबॉडीज कहलाते हैं समीपस्थ,ग्लाइकोलिक एसिड के चयापचय में एक महत्वपूर्ण भूमिका निभाते हैं, जो सीधे प्रकाश श्वसन से संबंधित है। हरी पत्तियों में, वे माइटोकॉन्ड्रिया और क्लोरोप्लास्ट से जुड़े होते हैं। अन्य माइक्रोबॉडीज कहलाते हैं ग्लाइऑक्सिसोम,वसा को कार्बोहाइड्रेट में बदलने के लिए आवश्यक एंजाइम होते हैं। यह कई बीजों में अंकुरण के दौरान होता है।
रिक्तिकाएं -ये झिल्ली द्वारा परिसीमित कोशिका के क्षेत्र हैं, जो तरल से भरे हुए हैं - सेल एसएपी।वे घिरे हुए हैं tonoplast(वैक्यूलर मेम्ब्रेन)।
एक युवा पादप कोशिका में कई छोटे रिक्तिकाएँ होती हैं, जो कोशिका आयु के रूप में एक बड़ी रसधानी में विलीन हो जाती हैं। एक परिपक्व कोशिका में, इसकी मात्रा का 90% तक रिक्तिका द्वारा कब्जा किया जा सकता है। इस मामले में, साइटोप्लाज्म कोशिका झिल्ली को एक पतली परिधीय परत के रूप में दबाया जाता है। कोशिका आकार में वृद्धि मुख्य रूप से रसधानी की वृद्धि के कारण होती है। नतीजतन, टर्गर दबाव उत्पन्न होता है और ऊतक लोच बनाए रखा जाता है। यह रिक्तिका और टोनोप्लास्ट के मुख्य कार्यों में से एक है।
रस का मुख्य घटक पानी है, बाकी पौधे के प्रकार और उसकी शारीरिक अवस्था के आधार पर भिन्न होता है। रिक्तिका में लवण, शर्करा, कम अक्सर प्रोटीन होते हैं। टोनोप्लास्ट रिक्तिका में कुछ आयनों के परिवहन और संचय में सक्रिय भूमिका निभाता है। सेल सैप में आयनों की सांद्रता पर्यावरण में इसकी सांद्रता से काफी अधिक हो सकती है। कुछ पदार्थों की उच्च सामग्री के साथ, रिक्तिका में क्रिस्टल बनते हैं। सबसे आम कैल्शियम ऑक्सालेट क्रिस्टल होते हैं, जिनका एक अलग आकार होता है।
रिक्तिकाएं चयापचय उत्पादों (चयापचय) के संचय के स्थान हैं। ये प्रोटीन, अम्ल और यहां तक कि मनुष्य के लिए विषाक्त पदार्थ (एल्कलॉइड) भी हो सकते हैं। पिगमेंट अक्सर जमा होते हैं। नीला, बैंगनी, बैंगनी, गहरा लाल, किरमिजी पौधे की कोशिकाओं को एंथोसायनिन समूह से वर्णक देते हैं। अन्य पिगमेंट के विपरीत, वे पानी में अच्छी तरह से घुल जाते हैं और सेल सैप में पाए जाते हैं। वे कई सब्जियों (मूली, शलजम, गोभी), फल (अंगूर, आलूबुखारा, चेरी), फूल (कॉर्नफ्लॉवर, जेरेनियम, डेल्फीनियम, गुलाब, peonies) के लाल और नीले रंग का निर्धारण करते हैं। कभी-कभी ये वर्णक पत्तियों में क्लोरोफिल को ढक लेते हैं, उदाहरण के लिए, सजावटी लाल मेपल में। एंथोसायनिन रंग शरद ऋतु चमकदार लाल छोड़ देता है। वे ठंडी धूप के मौसम में बनते हैं, जब पत्तियों में क्लोरोफिल का संश्लेषण बंद हो जाता है। पत्तियों में, जब एंथोसायनिन नहीं बनते हैं, क्लोरोफिल के विनाश के बाद, क्लोरोप्लास्ट के पीले-नारंगी कैरोटीनॉयड ध्यान देने योग्य हो जाते हैं। ठंडे, स्पष्ट शरद ऋतु में पत्तियाँ सबसे चमकीले रंग की होती हैं।
कोशिका में उनके घटकों के संचलन में, रिक्तिकाएं मैक्रोमोलेक्युलस के विनाश में शामिल होती हैं। राइबोसोम, माइटोकॉन्ड्रिया, प्लास्टिड, रिक्तिका में गिरकर नष्ट हो जाते हैं। इस पाचन क्रिया के द्वारा उनकी तुलना की जा सकती है लाइसोसोम- पशु कोशिकाओं के अंग।
एंडोप्लाज्मिक रेटिकुलम (रेटिकुलम) से रिक्तिकाएं बनती हैं
राइबोसोम।छोटे कण (17 - 23nm), जिसमें प्रोटीन और RNA की लगभग समान मात्रा होती है। राइबोसोम में, अमीनो एसिड मिलकर प्रोटीन बनाते हैं। सक्रिय चयापचय वाले कोशिकाओं में उनमें से अधिक हैं। राइबोसोम कोशिका के साइटोप्लाज्म में स्वतंत्र रूप से स्थित होते हैं या एंडोप्लाज्मिक रेटिकुलम (80S) से जुड़े होते हैं। वे नाभिक (80S), माइटोकॉन्ड्रिया (70S), प्लास्टिड्स (70S) में भी पाए जाते हैं।
राइबोसोम एक जटिल बना सकते हैं जिस पर समान पॉलीपेप्टाइड्स का एक साथ संश्लेषण होता है, जिसके बारे में जानकारी एक अणु और आरएनए से ली जाती है। ऐसा परिसर कहा जाता है पॉलीरिबोसोम (पॉलीसोम)।बड़ी मात्रा में प्रोटीन को संश्लेषित करने वाली कोशिकाओं में पॉलीसोम्स की एक व्यापक प्रणाली होती है, जो अक्सर परमाणु लिफाफे की बाहरी सतह से जुड़ी होती हैं।
अन्तः प्रदव्ययी जलिका।यह अनिश्चितकालीन सीमा की एक जटिल त्रि-आयामी झिल्ली प्रणाली है। ईआर के संदर्भ में उनके बीच एक संकीर्ण पारदर्शी स्थान के साथ दो प्राथमिक झिल्लियों की तरह दिखता है। ईआर का आकार और लंबाई सेल प्रकार, इसकी चयापचय गतिविधि और भेदभाव के चरण पर निर्भर करती है। कोशिकाओं में जो प्रोटीन को स्रावित या संग्रहीत करते हैं, ईआर फ्लैट थैली या के रूप में होता है कुंड,इसकी बाहरी सतह पर कई राइबोसोम जुड़े होते हैं। इस रेटिकुलम को कहा जाता है रफ अन्तर्द्रव्यी जालिका।एक चिकनी ईआर आमतौर पर आकार में ट्यूबलर होती है। खुरदरी और चिकनी अंतर्द्रव्यी जालिका एक ही कोशिका में मौजूद हो सकती है। एक नियम के रूप में, उनके बीच कई संबंध हैं।
एंडोप्लाज्मिक रेटिकुलम सेल की संचार प्रणाली के रूप में कार्य करता है। यह कर्नेल के बाहरी आवरण से जुड़ा होता है। वास्तव में, ये दो संरचनाएं एकल झिल्ली प्रणाली बनाती हैं। जब कोशिका विभाजन के दौरान परमाणु लिफाफा फट जाता है, तो इसके टुकड़े ER अंशों के समान हो जाते हैं। एंडोप्लाज्मिक रेटिकुलम पदार्थों के परिवहन के लिए एक प्रणाली है: प्रोटीन, लिपिड, कार्बोहाइड्रेट, कोशिका के विभिन्न भागों में। पड़ोसी कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम साइटोप्लाज्मिक स्ट्रैंड्स के माध्यम से जुड़े हुए हैं - प्लाज्मोडेसमाटा -जो कोशिका भित्ति से होकर गुजरते हैं।
कोशिका झिल्लियों के संश्लेषण के लिए एंडोप्लाज्मिक रेटिकुलम मुख्य स्थल है। कुछ पादप कोशिकाओं में, रसधानियों की झिल्लियाँ और सूक्ष्म शरीर, कुंड dictyosomes.
गॉल्जीकाय।यह शब्द सभी को संदर्भित करने के लिए प्रयोग किया जाता है तानाशाही, या गोलगी बॉडी, एक पिंजरे में। डिक्टियोसोम फ्लैट, डिस्क के आकार के पुटिकाओं या सिस्टर्न के समूह हैं, जो किनारों के साथ नलिकाओं की एक जटिल प्रणाली में शाखा करते हैं। उच्च पौधों में डिक्टायोसोम में एक साथ एकत्रित 4 - 8 कुंड होते हैं।
आमतौर पर, टैंकों के एक पैकेट में, एक विकासशील और परिपक्व पक्ष प्रतिष्ठित होता है। बनने वाले गढ्ढों की झिल्लियां संरचना में ईआर की झिल्लियों से मिलती-जुलती हैं, और परिपक्व हौदों की झिल्लियां प्लाज्मा झिल्ली से मिलती-जुलती हैं।
डिक्टायोसोम स्राव में शामिल होते हैं और अधिकांश उच्च पौधों में कोशिका झिल्लियों के निर्माण में शामिल होते हैं। डिक्टायोसोम द्वारा संश्लेषित कोशिका भित्ति पॉलीसेकेराइड पुटिकाओं में जमा हो जाते हैं, जो तब परिपक्व होने वाले सिस्टर्न से अलग हो जाते हैं। ये स्रावी पुटिकाएं सीएमपी के साथ माइग्रेट और फ्यूज हो जाती हैं; जबकि उनमें निहित पॉलीसेकेराइड कोशिका झिल्ली में निर्मित होते हैं। कुछ पदार्थ जो तानाशाहों में जमा होते हैं, अन्य संरचनाओं में बनते हैं, उदाहरण के लिए, ईआर में, और फिर तानाशाहों में ले जाया जाता है, जहां स्राव से पहले उन्हें संशोधित (संशोधित) किया जाता है। उदाहरण के लिए, ग्लाइकोप्रोटीन कोशिका झिल्ली की एक महत्वपूर्ण निर्माण सामग्री है। प्रोटीन भाग को किसी न किसी ईआर के पॉलीसोम द्वारा संश्लेषित किया जाता है, कार्बोहाइड्रेट भाग को डिक्टायोसोम में संश्लेषित किया जाता है, जहां दोनों भाग ग्लाइकोप्रोटीन बनाने के लिए संयोजित होते हैं।
झिल्लियां गतिशील, मोबाइल संरचनाएं हैं जो लगातार अपना आकार और क्षेत्र बदलती रहती हैं। अवधारणा झिल्ली गतिशीलता पर आधारित है एंडोप्लाज्मिक सिस्टम।इस अवधारणा के अनुसार, माइटोकॉन्ड्रिया और प्लास्टिड्स की झिल्लियों को छोड़कर, साइटोप्लाज्म की आंतरिक झिल्लियां एक ही पूरी होती हैं और एंडोप्लाज्मिक रेटिकुलम से उत्पन्न होती हैं। मध्यवर्ती पुटिका चरण के माध्यम से एंडोप्लाज्मिक रेटिकुलम से डिक्टायोसोम के नए कुंड बनते हैं, और स्रावी पुटिकाएं जो डिक्टीयोसोम से अलग होती हैं, अंततः प्लाज्मा झिल्ली के निर्माण में योगदान करती हैं। इस प्रकार, एंडोप्लाज्मिक रेटिकुलम और डिक्टियोसोम एक कार्यात्मक इकाई बनाते हैं जिसमें डिक्टायोसोम प्लाज्मा जैसी झिल्ली में एंडोप्लाज्मिक रेटिकुलम जैसी झिल्लियों को परिवर्तित करने की प्रक्रिया में मध्यवर्ती संरचनाओं की भूमिका निभाते हैं। उन ऊतकों में जिनकी कोशिकाएं बढ़ती हैं और कमजोर रूप से विभाजित होती हैं, झिल्ली के घटकों को लगातार अद्यतन किया जाता है।
सूक्ष्मनलिकाएंलगभग सभी यूकेरियोटिक कोशिकाओं में पाया जाता है। वे लगभग 24 एनएम के व्यास के साथ बेलनाकार संरचनाएं हैं। इनकी लंबाई अलग-अलग होती है। प्रत्येक नलिका एक प्रोटीन की उपइकाइयों से बनी होती है जिसे कहा जाता है ट्यूबुलिन।उपइकाइयां केंद्रीय गुहा के चारों ओर 13 अनुदैर्ध्य तंतु बनाती हैं। सूक्ष्मनलिकाएं गतिशील संरचनाएं हैं, वे नियमित रूप से नष्ट हो जाती हैं और कोशिका चक्र के कुछ चरणों में बनती हैं। इनका संयोजन विशेष स्थानों पर होता है, जिन्हें सूक्ष्मनलिकाय संगठन केंद्र कहते हैं। पादप कोशिकाओं में, उनके पास कमजोर व्यक्त अनाकार संरचना होती है।
सूक्ष्मनलिकाएं के कार्य: कोशिका झिल्ली के निर्माण में भाग लेते हैं; बनाने वाली झिल्ली के लिए तानाशाहों के सीधे बुलबुले, जैसे धुरी के धागे जो एक विभाजित कोशिका में बनते हैं; सेल प्लेट (बेटी कोशिकाओं के बीच प्रारंभिक सीमा) के निर्माण में भूमिका निभाते हैं। इसके अलावा, सूक्ष्मनलिकाएं फ्लैगेल्ला और सिलिया का एक महत्वपूर्ण घटक हैं, जिसके संचलन में वे महत्वपूर्ण भूमिका निभाते हैं।
सूक्ष्म तंतु,सूक्ष्मनलिकाएं की तरह, वे लगभग सभी यूकेरियोटिक कोशिकाओं में पाए जाते हैं। वे 5-7 एनएम मोटे लंबे तंतु होते हैं, जिनमें सिकुड़ा हुआ प्रोटीन एक्टिन होता है। उच्च पौधों की कई कोशिकाओं में माइक्रोफ़िल्मेंट्स के बंडल पाए जाते हैं। जाहिर है, वे साइटोप्लाज्म की धाराओं में एक महत्वपूर्ण भूमिका निभाते हैं। माइक्रोफ़िल्मेंट्स, सूक्ष्मनलिकाएं के साथ मिलकर एक लचीला नेटवर्क बनाते हैं जिसे कहा जाता है साइटोस्केलेटन।
आधार पदार्थलंबे समय तक एक सजातीय (सजातीय) प्रोटीन युक्त समाधान माना जाता था जिसमें कम संख्या में संरचनाएं या यहां तक कि संरचना रहित होती थी। हालांकि, वर्तमान में, एक उच्च-वोल्टेज इलेक्ट्रॉन माइक्रोस्कोप का उपयोग करके, यह स्थापित किया गया है कि जमीनी पदार्थ एक त्रि-आयामी जाली है जो पतली (3–6 एनएम व्यास) किस्में से निर्मित होती है जो पूरे सेल को भरती है। साइटोप्लाज्म के अन्य घटक, जिनमें सूक्ष्मनलिकाएं और माइक्रोफ़िल्मेंट्स शामिल हैं, इससे निलंबित हैं माइक्रोट्राबेकुलर ग्रिड।
माइक्रोट्राबेकुलर संरचना प्रोटीन स्ट्रैंड्स की एक जाली है, जिसके बीच का स्थान पानी से भरा होता है। पानी के साथ, जाली में जेल की स्थिरता होती है; जेल जिलेटिनस निकायों की तरह दिखता है।
ऑर्गेनेल माइक्रोट्राबेकुलर जाली से जुड़े होते हैं। जाली कोशिका के अलग-अलग हिस्सों के बीच संचार प्रदान करती है और इंट्रासेल्युलर परिवहन को निर्देशित करती है।
लिपिड बूँदें- एक गोलाकार आकार की संरचनाएं, एक प्रकाश सूक्ष्मदर्शी के तहत एक पादप कोशिका के साइटोप्लाज्म को ग्रैन्युलैरिटी देती हैं। इलेक्ट्रॉन माइक्रोग्राफ पर, वे अनाकार दिखते हैं। प्लास्टिड्स में बहुत समान लेकिन छोटी बूंदें पाई जाती हैं।
लिपिड बूँदें, गलती से उन्हें ऑर्गेनेल समझकर, उन्हें स्फेरोसोम्स कहते हैं और मानते हैं कि वे एक या दो परत वाली झिल्ली से घिरे हुए हैं। हालाँकि, हाल के आंकड़े बताते हैं कि लिपिड बूंदों में झिल्ली नहीं होती है, लेकिन प्रोटीन के साथ लेपित हो सकते हैं।
एर्गास्टिक पदार्थ -ये प्रोटोप्लास्ट के "निष्क्रिय उत्पाद" हैं: आरक्षित पदार्थ या अपशिष्ट उत्पाद। वे कोशिका चक्र की विभिन्न अवधियों में प्रकट और गायब हो सकते हैं। स्टार्च अनाज, क्रिस्टल, एंथोसायनिन पिगमेंट और लिपिड बूंदों को छोड़कर। इनमें रेजिन, गोंद, टैनिन और प्रोटीन शामिल हैं। एर्गैस्टिक पदार्थ कोशिका झिल्ली का हिस्सा होते हैं, साइटोप्लाज्म और ऑर्गेनेल का मुख्य पदार्थ, जिसमें रिक्तिकाएं भी शामिल हैं।
फ्लैगेल्ला और सिलिया -वे पतली, बालों जैसी संरचनाएं हैं जो कई यूकेरियोटिक कोशिकाओं की सतह से फैली हुई हैं। उनका एक स्थिर व्यास है, लेकिन लंबाई 2 से 150 माइक्रोन तक भिन्न होती है। परंपरागत रूप से, उनमें से लंबे और कुछ को फ्लैगेल्ला कहा जाता है, और छोटे और अधिक कई को सिलिया कहा जाता है। इन दो प्रकार की संरचनाओं के बीच कोई स्पष्ट अंतर नहीं है, इसलिए इस शब्द का प्रयोग दोनों को संदर्भित करने के लिए किया जाता है। कशाभिका।
कुछ शैवालों तथा कवकों में कशाभिका (Flagella) चलन अंग होते हैं, जिनकी सहायता से ये जल में गति करते हैं। पौधों में (उदाहरण के लिए, काई, लिवरवॉर्ट्स, फ़र्न, कुछ जिम्नोस्पर्म), केवल जर्म कोशिकाओं (युग्मक) में फ्लैगेल्ला होता है।
प्रत्येक फ्लैगेलम का एक विशिष्ट संगठन होता है। सूक्ष्मनलिकाएं के 9 जोड़े की एक बाहरी रिंग फ्लैगेलम के केंद्र में स्थित दो अतिरिक्त सूक्ष्मनलिकाएं को घेरे हुए है। एंजाइम युक्त "हैंडल" प्रत्येक बाहरी जोड़े के एक सूक्ष्मनलिका से फैलता है। यह यूकेरियोटिक जीवों में सभी कशाभों में पाया जाने वाला बुनियादी 9+2 संगठन पैटर्न है। यह माना जाता है कि फ्लैगेल्ला की गति सूक्ष्मनलिकाएं के फिसलने पर आधारित होती है, जबकि सूक्ष्मनलिकाएं के बाहरी जोड़े बिना संकुचन के एक दूसरे के साथ चलते हैं। एक दूसरे के सापेक्ष सूक्ष्मनलिकाएं के जोड़े के फिसलने से फ्लैगेलम का स्थानीय झुकाव होता है।
फ्लैगेल्ला साइटोप्लाज्मिक बेलनाकार संरचनाओं से "बढ़ता है" कहा जाता है बेसल निकाय,फ्लैगेलम का गठन और बेसल हिस्सा। बेसल निकायों में एक फ्लैगेलम जैसी आंतरिक संरचना होती है, सिवाय इसके कि बाहरी नलिकाओं को जोड़े के बजाय ट्रिपल में व्यवस्थित किया जाता है, और केंद्रीय नलिकाएं अनुपस्थित होती हैं।
कोशिका भित्ति।कोशिका भित्ति प्रोटोप्लास्ट के आकार को परिसीमित करती है और रिक्तिका द्वारा पानी के अवशोषण के कारण इसके टूटने को रोकती है।
कोशिका भित्ति में विशिष्ट कार्य होते हैं जो न केवल कोशिका और उस ऊतक के लिए महत्वपूर्ण होते हैं जिसमें कोशिका स्थित होती है, बल्कि पूरे पौधे के लिए भी महत्वपूर्ण होती है। कोशिका भित्ति पदार्थों के अवशोषण, परिवहन और उत्सर्जन में एक आवश्यक भूमिका निभाती है, और इसके अलावा, लाइसोसोमल या पाचन गतिविधि उनमें केंद्रित हो सकती है।
सेल की दीवार के घटक।कोशिका भित्ति का सबसे विशिष्ट घटक है सेलूलोज़,जो काफी हद तक इसकी वास्तुकला को निर्धारित करता है। सेल्युलोज अणु अंत से अंत तक जुड़े ग्लूकोज अणुओं को दोहराते हुए बने होते हैं। लंबे पतले सेल्युलोज अणु संयुक्त होते हैं सूक्ष्मतंतुमोटाई 10 - 25 एनएम। माइक्रोफ़ाइब्रिल्स आपस में जुड़ते हैं और पतले धागे बनाते हैं, जो बदले में एक दूसरे के चारों ओर लपेटे जा सकते हैं, जैसे कि रस्सी में किस्में। ऐसी प्रत्येक "रस्सी", या मैक्रोफिब्रिल की मोटाई लगभग 0.5 माइक्रोन होती है, जो 4 माइक्रोन की लंबाई तक पहुंचती है। मैक्रोफाइब्रिल समान आकार के स्टील के तार जितने मजबूत होते हैं।
सेल की दीवार का सेल्यूलोज फ्रेम इसके साथ जुड़े सेल्यूलोज मैट्रिक्स अणुओं से भरा होता है। इसमें पॉलीसेकेराइड कहा जाता है हेमिकेलुलोज,और पेक्टिन,या पेक्टिन,रासायनिक रूप से हेमिकेलुलोज के बहुत करीब।
कोशिका भित्ति का एक अन्य घटक है लिग्निन -पादप कोशिकाओं में सेल्युलोज के बाद सर्वाधिक मात्रा में पाया जाने वाला बहुलक है। लिग्निन दीवार की कठोरता को बढ़ाता है और आमतौर पर उन कोशिकाओं में पाया जाता है जो एक सहायक या यांत्रिक कार्य करती हैं।
क्यूटिन, सबरिन, वैक्स- आमतौर पर पौधों के सुरक्षात्मक ऊतकों के गोले में जमा होते हैं। क्यूटिन, उदाहरण के लिए, एपिडर्मिस की कोशिका झिल्लियों में पाया जाता है, और सुबेरिन द्वितीयक सुरक्षात्मक ऊतक, कॉर्क में पाया जाता है। दोनों पदार्थ वैक्स के संयोजन में पाए जाते हैं और पौधे से अत्यधिक पानी के नुकसान को रोकते हैं।
कोशिका भित्ति की परतें।पादप कोशिकाओं की दीवार की मोटाई व्यापक रूप से भिन्न होती है, जो पादप संरचना में कोशिकाओं की भूमिका और स्वयं कोशिका की आयु पर निर्भर करती है। एक इलेक्ट्रॉन सूक्ष्मदर्शी के नीचे, पौधे की कोशिका भित्ति में दो परतें दिखाई देती हैं: बीच की थाली(जिसे अंतरकोशिकीय पदार्थ भी कहा जाता है), और प्राथमिक कोशिका भित्ति।कई कोशिकाएं एक और परत बिछाती हैं - द्वितीयक कोशिका भित्ति।मीडियन प्लेट आसन्न कोशिकाओं की प्राथमिक दीवारों के बीच स्थित होती है। द्वितीयक दीवार, यदि कोई हो, प्राथमिक सेल दीवार की आंतरिक सतह पर सेल प्रोटोप्लास्ट द्वारा जमा की जाती है।
बीच की थाली।मीडियन लैमिना में मुख्य रूप से पेक्टिन होता है। जहां एक कोशिका भित्ति उत्पन्न होनी चाहिए, दो नवगठित कोशिकाओं के बीच, एंडोप्लाज़मिक रेटिकुलम के नलिकाओं का एक घना जाल और गोल्गी तंत्र (डिक्टायोसोम) का कुंड पहले नोट किया जाता है। फिर इस जगह में पेक्टिन (पॉलीसेकेराइड से) से भरे बुलबुले दिखाई देते हैं। इन बुलबुलों को गॉल्जी उपकरण की टंकियों से अलग किया जाता है। प्रारंभिक कोशिका भित्ति में विभिन्न पॉलीसेकेराइड होते हैं, जिनमें से मुख्य हैं पेक्टिन और हेमिकेलुलोज। बाद में, इसमें सघन पदार्थ - सेल्युलोज और लिग्निन होते हैं।
प्राथमिक कोशिका भित्ति।यह सेल्युलोज आवरण की एक परत है जो कोशिका वृद्धि से पहले या उसके दौरान जमा होती है। सेलूलोज़, हेमिसेल्यूलोज़ और पेक्टिन के अलावा, प्राथमिक झिल्ली में ग्लाइकोप्रोटीन होता है। प्राथमिक गोले लिग्निफाई कर सकते हैं। पेक्टिन घटक नमनीयता प्रदान करता है, जो प्राथमिक खोल को जड़, तना या पत्ती के रूप में विस्तारित करने की अनुमति देता है।
सक्रिय रूप से विभाजित होने वाली कोशिकाओं (प्रकाश संश्लेषण, श्वसन और स्राव की प्रक्रियाओं में शामिल अधिकांश परिपक्व कोशिकाओं) में प्राथमिक झिल्ली होती है। एक प्राथमिक झिल्ली और एक जीवित प्रोटोप्लास्ट वाली ऐसी कोशिकाएं अपने विशिष्ट आकार को खोने, विभाजित करने और एक नए प्रकार की कोशिका में अंतर करने में सक्षम होती हैं। वे पौधों में घाव भरने और ऊतक पुनर्जनन में शामिल हैं।
प्राथमिक कोशिका झिल्लियां अपनी पूरी लंबाई में मोटाई में एक समान नहीं होती हैं, लेकिन पतले खंड कहलाते हैं प्राथमिक छिद्र क्षेत्र।साइटोप्लाज्म, या प्लास्मोडेस्माटा की किस्में, जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ती हैं, आमतौर पर प्राथमिक छिद्र क्षेत्रों से गुजरती हैं।
द्वितीयक कोशिका भित्ति।इस तथ्य के बावजूद कि कई पादप कोशिकाओं में केवल एक प्राथमिक दीवार होती है, कुछ में, प्रोटोप्लास्ट कोशिका के केंद्र की ओर एक द्वितीयक दीवार जमा करता है। यह आमतौर पर कोशिका वृद्धि की समाप्ति के बाद होता है और प्राथमिक झिल्ली का क्षेत्र अब नहीं बढ़ता है। इस कारण से, द्वितीयक खोल प्राथमिक से भिन्न होता है। माध्यमिक झिल्लियों की विशेष रूप से विशेष कोशिकाओं द्वारा आवश्यकता होती है जो पौधे को मजबूत करती हैं और पानी का संचालन करती हैं। द्वितीयक झिल्ली के निक्षेपण के बाद, इन कोशिकाओं के प्रोटोप्लास्ट, एक नियम के रूप में, मर जाते हैं। माध्यमिक गोले में प्राथमिक की तुलना में अधिक सेल्यूलोज होता है, और उनमें पेक्टिन पदार्थ और ग्लाइकोप्रोटीन अनुपस्थित होते हैं। द्वितीयक झिल्ली को फैलाना मुश्किल होता है, इसके मैट्रिक्स में हेमिकेलुलोज होता है।
द्वितीयक खोल में, तीन परतों को प्रतिष्ठित किया जा सकता है - बाहरी, मध्य और आंतरिक (S 1, S 2, S 3)। द्वितीयक गोले की स्तरित संरचना उनकी ताकत में काफी वृद्धि करती है। द्वितीयक झिल्ली में सेल्युलोज माइक्रोफाइब्रिल्स प्राथमिक की तुलना में सघन रूप से जमा होते हैं। लिग्निन लकड़ी की द्वितीयक खाल का एक सामान्य घटक है।
संपर्क कोशिकाओं की झिल्लियों में छिद्र एक दूसरे के विपरीत स्थित होते हैं। दो विरोधी छिद्र और एक छिद्र झिल्ली बनती है कुछ बार।द्वितीयक झिल्लियों वाली कोशिकाओं में, दो मुख्य प्रकार के छिद्र होते हैं: सरल और सीमाबद्ध।सीमाबद्ध छिद्रों में, द्वितीयक झिल्ली लटकी रहती है छिद्र गुहा।सिंपल पोर्स में ऐसा नहीं होता है।
कोशिका भित्ति का विकास।जैसे-जैसे कोशिका बढ़ती है, कोशिका झिल्ली की मोटाई और क्षेत्रफल बढ़ता जाता है। शेल स्ट्रेचिंग एक जटिल प्रक्रिया है। यह प्रोटोप्लास्ट द्वारा नियंत्रित और एक हार्मोन द्वारा नियंत्रित होता है auxin.
सभी दिशाओं में समान रूप से बढ़ने वाली कोशिकाओं में, मायोफिब्रिल्स का जमाव यादृच्छिक होता है। ये मायोफिब्रिल एक अनियमित नेटवर्क बनाते हैं। ऐसी कोशिकाएं स्टेम कोर, स्टोरेज टिश्यू और इन विट्रो में सेल कल्टीवेशन के दौरान पाई जाती हैं। लम्बी कोशिकाओं में, पार्श्व आवरण के पेशीतंतुओं को बढ़ाव के अक्ष पर समकोण पर जमा किया जाता है।
मैट्रिक्स पदार्थ - पेक्टिन, हेमिकेलुलोज और ग्लाइकोप्रोटीन को डिक्टीयोसोम के पुटिकाओं में झिल्ली में स्थानांतरित किया जाता है। इसी समय, पेक्टिन बढ़ती कोशिकाओं की अधिक विशेषता है, जबकि हेमिकेलुलोज गैर-बढ़ती कोशिकाओं में प्रबल होते हैं।
सेल्युलोज माइक्रोफिब्रिल को कोशिका की सतह पर प्लाज्मा झिल्ली से जुड़े एक एंजाइम कॉम्प्लेक्स की मदद से संश्लेषित किया जाता है। सूक्ष्मतंतुओं का उन्मुखीकरण प्लाज्मा झिल्ली की आंतरिक सतह के पास स्थित सूक्ष्मनलिकाएं द्वारा नियंत्रित होता है।
प्लाज्मोडेस्मा।ये साइटोप्लाज्म के पतले धागे हैं जो पड़ोसी कोशिकाओं के प्रोटोप्लास्ट को जोड़ते हैं। प्लास्मोडेस्माटा या तो कहीं भी कोशिका भित्ति से होकर गुजरता है, या प्राथमिक छिद्र क्षेत्रों में या छिद्रों के जोड़े के बीच झिल्लियों में केंद्रित होता है। एक इलेक्ट्रॉन माइक्रोस्कोप के तहत, प्लास्मोडेमाटा प्लाज्मा झिल्ली के साथ पंक्तिबद्ध संकीर्ण चैनल के रूप में दिखाई देते हैं। एक छोटी बेलनाकार ट्यूब नहर की धुरी के साथ एक कोशिका से दूसरी कोशिका तक फैली होती है - डेस्मोट्यूब,जो दोनों आसन्न कोशिकाओं के एंडोप्लाज्मिक रेटिकुलम के साथ संचार करता है। कोशिका विभाजन के दौरान कई प्लास्मोडेस्माटा बनते हैं, जब ट्यूबलर एंडोप्लाज्मिक रेटिकुलम विकासशील सेल प्लेट द्वारा लिया जाता है। प्लास्मोडेस्माटा अविभाजित कोशिकाओं की झिल्लियों में भी बन सकता है। ये संरचनाएं कोशिका से कोशिका में कुछ पदार्थों का कुशल स्थानांतरण प्रदान करती हैं।
कोशिका विभाजन।बहुकोशिकीय जीवों में, कोशिका विभाजन, उनके आकार में वृद्धि के साथ, पूरे जीव के विकास का एक तरीका है। विभाजन के दौरान बनने वाली नई कोशिकाएं मूल कोशिका और एक दूसरे दोनों की संरचना और कार्य में समान होती हैं। यूकेरियोट्स में विभाजन प्रक्रिया को दो आंशिक रूप से अतिव्यापी चरणों में विभाजित किया जा सकता है: पिंजरे का बँटवाराऔर साइटोकाइनेसिस।
पिंजरे का बँटवारा- यह एक नाभिक से दो पुत्री नाभिकों का निर्माण है, रूपात्मक और आनुवंशिक रूप से एक दूसरे के समतुल्य। साइटोकिनेसिस -यह बेटी कोशिकाओं के निर्माण के साथ कोशिका के साइटोप्लाज्मिक भाग का विभाजन है।
कोशिका चक्र।एक जीवित कोशिका क्रमिक घटनाओं की एक श्रृंखला से गुजरती है जो कोशिका चक्र बनाती है। चक्र की अवधि स्वयं कोशिका के प्रकार और बाहरी कारकों जैसे तापमान या पोषक तत्वों की उपलब्धता के आधार पर भिन्न होती है। आमतौर पर चक्र में बांटा गया है interphaseऔर चार चरण समसूत्रण।
अंतरावस्था।क्रमिक माइटोटिक विभाजनों के बीच की अवधि।
इंटरपेज़ को तीन अवधियों में विभाजित किया गया है, जिसे G 1, S, G 2 के रूप में दर्शाया गया है।
जी 1 अवधि के दौरान, जो माइटोसिस के बाद शुरू होती है। इस अवधि के दौरान, विभिन्न जीवों सहित साइटोप्लाज्म की मात्रा बढ़ जाती है। इसके अलावा, आधुनिक परिकल्पना के अनुसार, जी 1 अवधि के दौरान, पदार्थ संश्लेषित होते हैं जो एस अवधि और शेष चक्र को उत्तेजित या बाधित करते हैं, इस प्रकार विभाजन प्रक्रिया का निर्धारण करते हैं।
एस अवधि जी 1 अवधि का अनुसरण करती है, जिस समय आनुवंशिक सामग्री (डीएनए) को दोहराया जाता है।
G2 अवधि के दौरान, जो S का अनुसरण करता है, संरचनाएं बनती हैं जो सीधे माइटोसिस में शामिल होती हैं, उदाहरण के लिए, स्पिंडल घटक।
कुछ कोशिकाएं असीमित संख्या में सेल चक्रों से गुजरती हैं। ये एककोशिकीय जीव और सक्रिय विकास क्षेत्रों (मेरिस्टेम) की कुछ कोशिकाएँ हैं। परिपक्वता के बाद कुछ विशिष्ट कोशिकाएं प्रजनन करने की अपनी क्षमता खो देती हैं। कोशिकाओं का तीसरा समूह, उदाहरण के लिए, घाव के ऊतक (कैलस) बनाने वाले, केवल विशेष परिस्थितियों में विभाजित करने की क्षमता को बरकरार रखते हैं।
सूत्रीविभाजन,या परमाणु विखंडन। यह एक सतत प्रक्रिया है जिसे चार चरणों में विभाजित किया गया है: प्रोफ़ेज़, मेटाफ़ेज़, एनाफ़ेज़, टेलोफ़ेज़।माइटोसिस के परिणामस्वरूप, आनुवंशिक सामग्री जो इंटरपेज़ में दोगुनी हो गई है, दो बेटी नाभिकों के बीच समान रूप से विभाजित हो जाती है।
विभाजन के लिए एक कोशिका के संक्रमण के शुरुआती संकेतों में से एक प्लाज्मा झिल्ली के ठीक नीचे सूक्ष्मनलिकाएं के एक संकीर्ण, कुंडलाकार बैंड की उपस्थिति है। यह अपेक्षाकृत सघन बेल्ट भविष्य के माइटोटिक स्पिंडल के विषुवतीय तल में नाभिक को घेरता है। चूंकि यह प्रोफ़ेज़ से पहले प्रकट होता है, इसलिए इसे कहा जाता है प्रीप्रोफ़ेज़ बेल्ट।यह माइटोटिक स्पिंडल के बाद गायब हो जाता है, सेल प्लेट के लेट टेलोफ़ेज़ में प्रकट होने से बहुत पहले, जो केंद्र से परिधि तक बढ़ता है और प्रीप्रोफ़ेज़ बेल्ट द्वारा कब्जा किए गए क्षेत्र में मातृ कोशिका झिल्ली के साथ विलीन हो जाता है।
प्रोफ़ेज़।प्रोफ़ेज़ की शुरुआत में, गुणसूत्र नाभिक के अंदर बिखरे हुए लंबे धागों के समान होते हैं। फिर, जैसे-जैसे तंतु छोटे और मोटे होते हैं, यह देखा जा सकता है कि प्रत्येक गुणसूत्र में एक नहीं, बल्कि दो गुंथे हुए तंतु होते हैं, जिन्हें कहा जाता है क्रोमैटिड।देर से प्रोफ़ेज़ में, प्रत्येक गुणसूत्र के दो छोटे जोड़े वाले क्रोमैटिड समानांतर में समानांतर होते हैं, जो एक संकीर्ण खंड से जुड़े होते हैं जिन्हें कहा जाता है सेंट्रोमियर।यह प्रत्येक गुणसूत्र पर एक विशिष्ट स्थिति रखता है और गुणसूत्र को अलग-अलग लंबाई की दो भुजाओं में विभाजित करता है।
सूक्ष्मनलिकाएं धुरी की धुरी के साथ नाभिक की सतह के समानांतर स्थित होती हैं। यह माइटोटिक स्पिंडल असेंबली की सबसे पहली अभिव्यक्ति है।
प्रोफ़ेज़ के अंत तक, नाभिक धीरे-धीरे अपनी स्पष्ट रूपरेखा खो देता है और अंत में गायब हो जाता है। इसके तुरंत बाद, परमाणु लिफाफा भी बिखर जाता है।
रूपक।सर्वप्रथम रूपक धुरी,जो एक त्रि-आयामी संरचना का प्रतिनिधित्व करता है, मध्य में सबसे चौड़ा और ध्रुवों की ओर पतला होता है, जो पहले नाभिक द्वारा कब्जा कर लिया गया स्थान लेता है। स्पिंडल फाइबर सूक्ष्मनलिकाएं के बंडल हैं। मेटाफ़ेज़ के दौरान, क्रोमोसोम, जिनमें से प्रत्येक में दो क्रोमैटिड होते हैं, को व्यवस्थित किया जाता है ताकि उनके सेंट्रोमर्स स्पिंडल के इक्वेटोरियल प्लेन में स्थित हों। इसके सेंट्रोमियर के साथ, प्रत्येक गुणसूत्र स्पिंडल थ्रेड्स से जुड़ा होता है। हालाँकि, कुछ किस्में गुणसूत्रों से जुड़े बिना एक ध्रुव से दूसरे ध्रुव तक जाती हैं।
जब सभी गुणसूत्र विषुवतीय तल में स्थित होते हैं, तो मेटाफ़ेज़ पूरा हो जाता है। गुणसूत्र विभाजित होने के लिए तैयार होते हैं।
पश्चावस्था।प्रत्येक गुणसूत्र के क्रोमैटिड अलग हो जाते हैं। अभी इसे बच्चागुणसूत्र। सबसे पहले, सेंट्रोमियर विभाजित होता है, और दो बेटी गुणसूत्र विपरीत ध्रुवों पर खींचे जाते हैं। इस मामले में, सेंट्रोमर्स आगे बढ़ते हैं, और गुणसूत्रों की भुजाएँ पीछे की ओर खिंचती हैं। क्रोमोसोम से जुड़े स्पिंडल फिलामेंट्स छोटे हो जाते हैं, क्रोमैटिड्स को अलग करने और विपरीत दिशाओं में बेटी क्रोमोसोम के आंदोलन में योगदान करते हैं।
टेलोफ़ेज़।टेलोफ़ेज़ में, गुणसूत्रों के दो समान समूहों का पृथक्करण पूरा हो जाता है, जबकि उनमें से प्रत्येक के चारों ओर एक परमाणु झिल्ली बन जाती है। रफ रेटिकुलम इसमें सक्रिय भाग लेता है। धुरी तंत्र गायब हो जाता है। टेलोफ़ेज़ के दौरान, गुणसूत्र अपनी रूपरेखा की स्पष्टता खो देते हैं, फैल जाते हैं, पतले धागों में बदल जाते हैं। न्यूक्लियोली को बहाल किया जाता है। जब गुणसूत्र अदृश्य हो जाते हैं, तो माइटोसिस पूरा हो जाता है। दो संतति केंद्रक अंतरावस्था में प्रवेश करते हैं। वे आनुवंशिक रूप से एक दूसरे के और मूल नाभिक के बराबर हैं। यह बहुत महत्वपूर्ण है, क्योंकि अनुवांशिक कार्यक्रम, और इसके साथ सभी संकेतों को बेटी जीवों में स्थानांतरित किया जाना चाहिए।
माइटोसिस की अवधि विभिन्न जीवों में भिन्न होती है और ऊतक के प्रकार पर निर्भर करती है। हालाँकि, प्रोफ़ेज़ सबसे लंबा है और एनाफ़ेज़ सबसे छोटा है। रूट टिप की कोशिकाओं में, प्रोफ़ेज़ की अवधि 1-2 घंटे होती है; रूपक - 5 - 15 मिनट; पश्चावस्था - 2 - 10 मिनट; टेलोफ़ेज़ - 10 - 30 मिनट। इंटरपेज़ की अवधि 12 से 30 घंटे तक है।
कई यूकेरियोटिक कोशिकाओं में माइटोटिक स्पिंडल के गठन के लिए जिम्मेदार सूक्ष्मनलिका आयोजन केंद्र जुड़े हुए हैं केन्द्रक.
साइटोकिनेसिस।यह साइटोप्लाज्म के विभाजन की प्रक्रिया है। अधिकांश जीवों में, कोशिकाएँ कोशिका झिल्ली को पीछे खींचकर और एक विखंडन खांचे का निर्माण करके विभाजित होती हैं, जो धीरे-धीरे गहरा होता है, माइटोटिक स्पिंडल के शेष तंतुओं को संकुचित करता है। सभी पौधों (ब्रायोफाइट्स और संवहनी) में और कुछ शैवाल में, कोशिकाएँ गठन के कारण विभाजित होती हैं। कोशिका की थाली।
आरंभिक टीलोफ़ेज़ में, दो संतति केन्द्रकों के बीच तंतुओं की एक नली के आकार की प्रणाली बनती है, जिसे कहते हैं phragmoplast.फैगमोप्लास्ट फाइबर, माइटोटिक स्पिंडल के तंतुओं की तरह, सूक्ष्मनलिकाएं होते हैं। फ्रैगमोप्लास्ट के विषुवतीय तल में छोटी-छोटी बूंदें दिखाई देती हैं। वे एक सेल प्लेट बनाने के लिए फ्यूज हो जाते हैं जो तब तक बढ़ता है जब तक कि वह विभाजित सेल की दीवार तक नहीं पहुंच जाता। यह दो बेटी कोशिकाओं के अलगाव को पूरा करता है। विलय की बूंदें गोल्गी तंत्र से निकलने वाले बुलबुले हैं। मूल रूप से, उनमें पेक्टिन पदार्थ होते हैं, जिनसे मध्य प्लेट बनती है। प्लेट के दोनों किनारों पर प्लाज्मा झिल्ली के निर्माण में पुटिका झिल्ली शामिल होती है। इसी समय, ट्यूबलर एंडोप्लाज्मिक रेटिकुलम के टुकड़ों से प्लास्मोडेस्मेटा बनता है।
मीडियन प्लेट के बनने के बाद, प्रत्येक प्रोटोप्लास्ट उस पर एक प्राथमिक झिल्ली जमा करता है। इसके अलावा, प्रत्येक बेटी कोशिका पूरे प्रोटोप्लास्ट के चारों ओर झिल्ली की एक नई परत जमा करती है, जो कोशिका प्लेट से उत्पन्न हुई झिल्ली को जारी रखती है। बेटी कोशिकाओं के बढ़ने पर मूल कोशिका का मूल खोल नष्ट हो जाता है।
विभिन्न प्रकार के यूकेरियोटिक माइटोसिस
पौधों और जानवरों में ऊपर वर्णित कोशिका विभाजन अप्रत्यक्ष कोशिका विभाजन का एकमात्र रूप नहीं है। माइटोसिस का सबसे सरल प्रकार है प्लूरोमिटोसिस।यह बाइनरी डिवीजन जैसा दिखता है प्रोकैरियोटिक कोशिकाएं, जिसमें प्रतिकृति के बाद न्यूक्लियॉइड प्लाज्मा झिल्ली से जुड़े रहते हैं। झिल्ली डीएनए बंधन बिंदुओं के बीच बढ़ने लगती है और इस तरह क्रोमोसोम को कोशिका के विभिन्न भागों में फैला देती है। उसके बाद, एक कोशिका संकुचन के निर्माण के दौरान, प्रत्येक डीएनए अणु एक नई अलग कोशिका में होगा।
विभाजन के लिए विशेषता यूकेरियोटिक कोशिकाएंसूक्ष्मनलिकाएं से निर्मित एक धुरी का निर्माण है। पर बंद प्लूरोमिटोसिस(इसे बंद कहा जाता है क्योंकि गुणसूत्रों का विचलन परमाणु लिफाफे को बाधित किए बिना होता है) सेंट्रीओल्स नहीं, बल्कि परमाणु झिल्ली के अंदरूनी हिस्से में स्थित अन्य संरचनाएं सूक्ष्मनलिका आयोजन केंद्र (MCMT) के रूप में भाग लेती हैं। ये अनिश्चित आकृति विज्ञान के तथाकथित ध्रुवीय निकाय हैं, जिनसे सूक्ष्मनलिकाएं फैलती हैं। इनमें से दो शरीर हैं। वे परमाणु लिफाफे से संपर्क खोए बिना एक दूसरे से अलग हो जाते हैं। इसके परिणामस्वरूप, गुणसूत्रों से जुड़े दो अर्ध-स्पिंडल बनते हैं। इस मामले में माइटोटिक स्पिंडल गठन और गुणसूत्र अलगाव की पूरी प्रक्रिया परमाणु लिफाफे के अंतर्गत होती है। इस प्रकार का माइटोसिस प्रोटोजोआ के बीच होता है और व्यापक रूप से कवक (काइट्रिडिया, ज़ाइगोमाइसेट्स, यीस्ट, ओमीसाइकेट्स, एस्कोमाइसेट्स, मायक्सोमाइसेट्स, आदि) में वितरित किया जाता है। रूपों से मिलें अर्ध-बंद प्लूरोमिटोसिसजब गठित धुरी के ध्रुवों पर परमाणु लिफाफा नष्ट हो जाता है।
माइटोसिस का अगला रूप है ऑर्थोमाइटोसिस।इस मामले में, COMTs साइटोप्लाज्म में स्थित होते हैं, शुरुआत से ही, अर्ध-स्पिंडल नहीं बनते हैं, लेकिन एक द्विध्रुवीय स्पिंडल होता है। तीन रूप हैं ऑर्थोमाइटोसिस (सामान्य माइटोसिस) अर्द्ध बंदऔर बंद किया हुआ. अर्ध-बंद ऑर्थोमाइटोसिस में, साइटोप्लाज्म में स्थित TsOMT की मदद से एक बिसिमेट्रिक स्पिंडल बनता है, ध्रुवीय क्षेत्रों के अपवाद के साथ, परमाणु लिफाफा पूरे माइटोसिस में संरक्षित होता है। दानेदार सामग्री, या सेंट्रीओल्स के द्रव्यमान को COMT के रूप में पहचाना जा सकता है। माइटोसिस का यह रूप हरे, भूरे और लाल शैवाल के जूस्पोर्स में पाया जाता है, कुछ निचले कवकों में और ग्रेगरीन में। बंद ऑर्थोमाइटोसिस के साथ, परमाणु झिल्ली पूरी तरह से संरक्षित होती है, जिसमें एक वास्तविक धुरी बनती है। सूक्ष्मनलिकाएं कार्योप्लाज्म में बनती हैं, कम बार वे आंतरिक TsOMT से बढ़ती हैं, जो परमाणु झिल्ली के साथ संबद्ध (प्लुरोमिटोसिस के विपरीत) नहीं होती है। इस प्रकार का माइटोसिस सिलियेट माइक्रोन्यूक्लि के विभाजन के लिए विशिष्ट है, लेकिन प्रोटोजोआ में भी हो सकता है। खुले ऑर्थोमाइटोसिस में, परमाणु लिफाफा पूरी तरह से विघटित हो जाता है। इस प्रकार का कोशिका विभाजन पशु जीवों, कुछ प्रोटोजोआ और उच्च पौधों की कोशिकाओं की विशेषता है। माइटोसिस का यह रूप, बदले में, सूक्ष्म और अनास्ट्रल प्रकारों द्वारा दर्शाया जाता है।
संक्षेप में समीक्षा की गई सामग्री से, यह देखा जा सकता है कि सामान्य रूप से माइटोसिस की मुख्य विशेषता विखंडन धुरी संरचनाओं की उपस्थिति है, जो सीएमटी के संबंध में बनती हैं, जो उनकी संरचना में विविध हैं।
माइटोटिक आकृति की आकृति विज्ञान
माइटोसिस के मेटाफ़ेज़ चरण में माइटोटिक उपकरण विशेष रूप से अच्छी तरह से व्यक्त किया गया है। मेटाफ़ेज़ में, कोशिका के भूमध्यरेखीय तल में, गुणसूत्र स्थित होते हैं, जिनसे तथाकथित तकला धागे,माइटोटिक आकृति के दो अलग-अलग ध्रुवों पर अभिसरण। इस प्रकार, माइटोटिक स्पिंडल गुणसूत्रों, ध्रुवों और तंतुओं का एक संग्रह है। धुरी के तंतु एकल सूक्ष्मनलिकाएं या उनके बंडल होते हैं। सूक्ष्मनलिकाएं धुरी के ध्रुवों से शुरू होती हैं, और उनमें से कुछ जाती हैं सेंट्रोमर्स को कहाँ स्थित हैं गुणसूत्रबिंदुओं क्रोमोसोम (कीनेटोकोर सूक्ष्मनलिकाएं), कुछ विपरीत ध्रुव की ओर आगे बढ़ते हैं, लेकिन उस तक नहीं पहुंच पाते हैं। उन्हें "इंटरपोलर माइक्रोट्यूबुल्स" कहा जाता है। रेडियल सूक्ष्मनलिकाएं का एक समूह ध्रुवों से निकलता है, जिससे उनके चारों ओर एक "उज्ज्वल चमक" जैसी संरचना बनती है - ये सूक्ष्म सूक्ष्मनलिकाएं हैं।
आकारिकी द्वारा, माइटोटिक आकृतियों को सूक्ष्म और अनास्ट्रल प्रकारों में विभाजित किया जाता है।
सूक्ष्म धुरी प्रकार, या अभिसारी, इस तथ्य की विशेषता है कि इसके ध्रुवों को एक छोटे से क्षेत्र द्वारा दर्शाया जाता है जिसमें सूक्ष्मनलिकाएं अभिसरण (अभिसरण) करती हैं। आमतौर पर, सेंट्रीओल्स वाले सेंट्रोसोम सूक्ष्म स्पिंडल के ध्रुवों पर स्थित होते हैं। हालांकि, सेंट्रीओलर एस्ट्रल माइटोस के मामले ज्ञात हैं (कुछ अकशेरूकीय के अर्धसूत्रीविभाजन के दौरान)। इसके अलावा, ध्रुवों से निकलने वाली रेडियल सूक्ष्मनलिकाएं नोट की जाती हैं, जो धुरी का हिस्सा नहीं हैं, लेकिन तारकीय क्षेत्र - सिटास्टर बनाती हैं। इस प्रकार का माइटोटिक डिवीजन एक डंबल जैसा दिखता है। माइटोटिक आकृति के एनास्ट्रियल प्रकार में ध्रुवों पर सिटास्टर नहीं होते हैं। यहां धुरी के ध्रुवीय क्षेत्र चौड़े हैं, उन्हें ध्रुवीय टोपी कहा जाता है, उनमें सेंट्रीओल्स शामिल हैं। इस मामले में, धुरी के तंतु एक बिंदु से नहीं हटते हैं, लेकिन ध्रुवीय टोपी के पूरे क्षेत्र से एक विस्तृत मोर्चे (विचलन) में विचलन करते हैं। इस प्रकार की धुरी उच्च पौधों की कोशिकाओं को विभाजित करने की विशेषता है, लेकिन उच्च जानवरों में भी पाई जा सकती है। स्तनधारियों के प्रारंभिक भ्रूणजनन में, सेंट्रीओलर (अपसारी) माइटोस डिम्बाणुजनकोशिका परिपक्वता के विभाजन के दौरान और जाइगोट के विभाजन I और II में देखे जाते हैं। लेकिन पहले से ही तीसरे कोशिका विभाजन में और बाद की सभी कोशिकाओं में, वे सूक्ष्म स्पिंडल की भागीदारी के साथ विभाजित होते हैं, जिनके ध्रुवों में सेंट्रीओल्स हमेशा पाए जाते हैं। सामान्य तौर पर, माइटोसिस के सभी रूपों के लिए, उनके किनेटोफ़ोर्स, ध्रुवीय निकायों (सेंट्रोसोम्स) और स्पिंडल फाइबर के साथ गुणसूत्र सामान्य संरचनाएं बने रहते हैं।
सेंट्रोमियर और काइनेटोकोर
क्रोमोसोम की लंबाई के साथ सेंट्रोमर्स का अलग-अलग स्थानीयकरण हो सकता है। holocentricसेंट्रोमियर तब होते हैं जब सूक्ष्मनलिकाएं पूरे गुणसूत्र (कुछ कीड़े, नेमाटोड, कुछ पौधे) की लंबाई के साथ जुड़ी होती हैं। मोनोसेंट्रिकसेंट्रोमियर - जब सूक्ष्मनलिकाएं एक क्षेत्र में गुणसूत्रों से जुड़ी होती हैं। मोनोसेंट्रिक सेंट्रोमर्स हो सकते हैं तुच्छ(उदाहरण के लिए, कुछ नवोदित यीस्ट में), जब केवल एक सूक्ष्मनलिका कीनेटोकोर तक पहुँचती है, और जोनल, जहां सूक्ष्मनलिकाएं का एक बंडल जटिल कीनेटोकोर तक पहुंचता है। सेंट्रोमियर ज़ोन की विविधता के बावजूद, वे सभी एक जटिल संरचना से जुड़े हैं। kinetocore, जिसकी संरचना और कार्य में सभी यूकेरियोट्स में मौलिक समानता है। काइनेटोकोर विशेष प्रोटीन संरचनाएं हैं, जो ज्यादातर क्रोमोसोम के सेंट्रोमियर जोन में स्थित होती हैं। ये कई प्रोटीनों से युक्त जटिल परिसर हैं। रूपात्मक रूप से, वे बहुत समान हैं, समान संरचना है, डायटम से लेकर मनुष्यों तक। वे तीन-परत संरचनाएं हैं: गुणसूत्र के शरीर से सटे आंतरिक घने परत, मध्य ढीली परत और बाहरी घने परत। कई तंतु बाहरी परत से फैलते हैं, जो कीनेटोचोर के तथाकथित रेशेदार मुकुट का निर्माण करते हैं। सामान्य रूप में, कीनेटोकोर्स क्रोमोसोम के प्राथमिक कसना के क्षेत्र में स्थित प्लेट या डिस्क की तरह दिखते हैं, सेंट्रोमियर में। आमतौर पर प्रति क्रोमोसोम या क्रोमैटिड में एक काइनेटोकोर होता है। एनाफेज से पहले, प्रत्येक बहन क्रोमैटिड पर काइनेटोकोर्स स्थित होते हैं, प्रत्येक सूक्ष्मनलिकाएं के अपने स्वयं के बंडल के साथ संचार करता है। पौधों में, काइनेटोकोर प्लेटों की तरह नहीं, बल्कि गोलार्द्धों की तरह दिखता है। कीनेटोकोर्स की कार्यात्मक भूमिका बहन क्रोमैटिड्स को एक दूसरे से बांधना है, माइटोटिक माइक्रोट्यूबुल्स को ठीक करना, क्रोमोसोम पृथक्करण को विनियमित करना और वास्तव में माइटोसिस के दौरान माइक्रोट्यूबुल्स की भागीदारी के साथ क्रोमोसोम को स्थानांतरित करना है। सामान्य तौर पर, प्रोटीन संरचनाएं, किनेटोकोर्स, एस-अवधि में दोगुनी, गुणसूत्र दोहरीकरण के समानांतर। लेकिन उनके प्रोटीन कोशिका चक्र की सभी अवधियों में गुणसूत्रों पर मौजूद होते हैं।
संयंत्र ऊतक
शैक्षिक ऊतक (मेरिस्टेम)
पौधों के शरीर में शैक्षिक ऊतक विभिन्न स्थानों पर स्थित होते हैं, इसलिए उन्हें निम्नलिखित समूहों में विभाजित किया जाता है (चित्र 0;1)।
एपिकल (एपिकल) मेरिस्टेम्सअक्षीय अंगों के शीर्ष, या शीर्ष पर स्थित - तना, जड़। इन विभज्योतकों की सहायता से पौधों के कायिक अंग लंबाई में वृद्धि करते हैं।
पार्श्व विभज्योतकअक्षीय अंगों की विशेषता। वहां वे क्लच के रूप में केंद्रित रूप से स्थित हैं।
इंटरकैलरी,या इंटरक्लेरी, मेरिस्टेम्सएपिकल मेरिस्टेम से उत्पन्न। ये कोशिकाओं के समूह हैं जो अभी तक गुणा करने में सक्षम नहीं हैं, लेकिन भेदभाव के मार्ग पर चल पड़े हैं। उनमें कोई प्रारंभिक कोशिकाएँ नहीं हैं, लेकिन कई विशिष्ट कोशिकाएँ हैं।
घाव विभज्योतकशरीर के क्षतिग्रस्त हिस्से की बहाली प्रदान करें। पुनर्जनन की शुरुआत निर्विभेदीकरण से होती है, यानी विशिष्ट कोशिकाओं से विभज्योतक कोशिकाओं तक का उल्टा विकास। वे बदल जाते हैं साथी,जो बनता है कॉर्कघाव की सतह को ढंकना। विविभेदित कोशिकाएं, विभाजित होकर ढीले पैरेन्काइमल ऊतक बना सकती हैं - घट्टा।इससे कुछ शर्तों के तहत पौधे के अंगों का निर्माण होता है।
पूर्णांक ऊतक
वे एक सीमा बाधा के रूप में कार्य करते हैं, अंतर्निहित ऊतकों को पर्यावरण से अलग करते हैं। पौधे के प्राथमिक अध्यावरण में केवल जीवित कोशिकाएँ होती हैं। द्वितीयक और तृतीयक अध्यावरण - अधिकतर मृत कोशिका भित्तियों से प्राप्त होता है।
पूर्णांक ऊतकों के मुख्य कार्य:
पौधे को सूखने से बचाना;
हानिकारक सूक्ष्मजीवों के प्रवेश से सुरक्षा;
सनबर्न सुरक्षा;
यांत्रिक क्षति से सुरक्षा;
संयंत्र और पर्यावरण के बीच चयापचय का विनियमन;
जलन की धारणा।
प्राथमिक पूर्णांक ऊतक - एपिडर्मिस, एपिडर्मिस . जीवित कोशिकाओं से मिलकर बनता है। शीर्ष विभज्योतक से निर्मित। युवा बढ़ते तनों और पत्तियों को ढकता है।
एपिडर्मिस का गठन पौधों में जलीय आवास से बाहर निकलने के संबंध में किया गया था ताकि इसे सूखने से रोका जा सके। स्टोमेटा के अलावा, एपिडर्मिस की सभी कोशिकाएं कसकर आपस में जुड़ी होती हैं। मुख्य कोशिकाओं की बाहरी दीवारें बाकी की तुलना में मोटी होती हैं। पूरी सतह क्यूटिन और वेजिटेबल वैक्स की एक परत से ढकी होती है। यह परत कहलाती है छल्ली(त्वचा)। यह पौधों की बढ़ती जड़ों और पानी के नीचे के हिस्सों पर अनुपस्थित है। सूखने पर छल्ली की पारगम्यता काफी कमजोर हो जाती है।
के अलावा बुनियादी कोशिकाएं, विशेष रूप से एपिडर्मिस में अन्य हैं बाल, या ट्राइकोम्स. वे एककोशिकीय और बहुकोशिकीय हैं (अंक 2)। कार्यात्मक रूप से, वे एपिडर्मिस की सतह को बढ़ाते हैं, उदाहरण के लिए, जड़ विकास क्षेत्र में, यांत्रिक सुरक्षा के रूप में कार्य करते हैं, एक समर्थन से चिपकते हैं, और पानी के नुकसान को कम करते हैं। बहुत से पौधे हैं ग्रंथियों के बालजैसे बिछुआ।
एपिडर्मिस में केवल उच्च पौधों में ही होते हैं रंध्रजो पानी और गैसों के आदान-प्रदान को नियंत्रित करता है। यदि छल्ली नहीं है, तो रंध्रों की कोई आवश्यकता नहीं है। स्टोमेटा कोशिकाओं का एक समूह है जो बनता है रंध्र तंत्र,जिसमें दो होते हैं रक्षक कोषऔर आसपास के एपिडर्मल कोशिकाएं पार्श्व कोशिकाएं. वे मुख्य एपिडर्मल कोशिकाओं से भिन्न होते हैं (चित्र 3 ). गार्ड कोशिकाएं अपने आसपास की कोशिकाओं से आकार और बड़ी संख्या में क्लोरोप्लास्ट और असमान रूप से मोटी दीवारों की उपस्थिति से भिन्न होती हैं। जो एक दूसरे का सामना करते हैं वे बाकी की तुलना में मोटे होते हैं (अंजीर.4) . गार्ड कोशिकाओं के बीच बनता है रंध्र खोलना, जिससे होता है सबस्टोमेटल स्पेसबुलाया सबस्टोमेटल कैविटी।गार्ड कोशिकाएं अत्यधिक प्रकाश संश्लेषक होती हैं। उनमें बड़ी मात्रा में आरक्षित स्टार्च और कई माइटोकॉन्ड्रिया होते हैं।
रंध्रों की संख्या और वितरण, रंध्र तंत्र के प्रकार विभिन्न पौधों में व्यापक रूप से भिन्न होते हैं। आधुनिक ब्रायोफाइट्स में रंध्र अनुपस्थित होते हैं। उनमें प्रकाश संश्लेषण गैमेटोफाइट पीढ़ी द्वारा किया जाता है, और स्पोरोफाइट स्वतंत्र अस्तित्व में सक्षम नहीं होते हैं।
आमतौर पर रंध्र पत्ती के नीचे की तरफ स्थित होते हैं। पानी की सतह पर तैरने वाले पौधों में - ऊपरी सतह पर। अनाज की पत्तियों में, रंध्र अक्सर दोनों तरफ समान रूप से स्थित होते हैं। ऐसी पत्तियाँ अपेक्षाकृत समान रूप से प्रकाशित होती हैं। सतह के 1 मिमी 2 पर, 100 से 700 रंध्र स्थित हो सकते हैं।
द्वितीयक पूर्णांक ऊतक (पेरिडर्म)। जब वार्षिक प्ररोहों का हरा रंग भूरा हो जाता है तो यह ऊतक एपिडर्मिस की जगह ले लेता है। यह बहुपरतीय होती है और कैम्बियल कोशिकाओं की एक केंद्रीय परत होती है - phlogen.फेलोजन कोशिकाएं, विभाजित होकर, बाहर एक परत बिछाती हैं दोस्तों,और अंदर - फेलोडर्मा(चित्र 5)।
फेलिमा, या कॉर्क. सबसे पहले इसमें जीवित पतली दीवार वाली कोशिकाएँ होती हैं। समय के साथ, उनकी दीवारें साबरिन और वनस्पति मोम से संतृप्त हो जाती हैं और मर जाती हैं। सेल की सामग्री हवा से भरी हुई है।
फेलिमा कार्य:
नमी के नुकसान को रोकता है;
पौधे को यांत्रिक क्षति से बचाता है;
रोगजनकों से बचाता है;
थर्मल इन्सुलेशन प्रदान करता है, क्योंकि कोशिकाएं हवा से भरी होती हैं।
एपिडर्मिस में स्थित फेलोजेन की कोशिकाएं, अंतर्निहित सबपीडर्मल परत, प्राथमिक प्रांतस्था की गहरी परतों में अक्सर कम होती हैं, प्राथमिक प्रांतस्था का निर्माण आधार होती हैं।
कॉर्क परत स्थायी नहीं है। इसमें ब्रेक होते हैं, जो पास में स्थित इंटरसेलुलर स्पेस के साथ संचार करते हैं। उसी समय, सतह पर छोटे ट्यूबरकल बनते हैं - मसूर की दाल, जो वायुमंडलीय वायु के साथ अंतरकोशिकीय स्थानों का संचार करते हैं (चित्र 6,7)।
शरद ऋतु में, मसूर के नीचे फेलोजेन कॉर्क कोशिकाओं की एक परत देता है, जो वाष्पोत्सर्जन को बहुत कम करता है, लेकिन इसे पूरी तरह से बाहर नहीं करता है। वसंत ऋतु में यह परत अंदर से ढह जाती है। हल्की बर्च की छाल पर, काले धब्बे के रूप में दाल स्पष्ट रूप से दिखाई देती है।
तृतीयक पूर्णांक ऊतक (क्रस्ट),केवल लकड़ी के पौधों के रूपों की भी विशेषता है।
कॉर्टेक्स की गहरी परतों में फेलोजेन को बार-बार रखा जाता है। इसके बाहर के ऊतक अंततः मर जाते हैं, एक पपड़ी बनाते हैं। इसकी कोशिकाएँ मर चुकी होती हैं और फैलने में असमर्थ होती हैं। हालांकि, जीवित कोशिकाएं गहरे विभाजन में स्थित होती हैं, जिससे तने के अनुप्रस्थ आकार में वृद्धि होती है। समय के साथ, परत की बाहरी परत टूट जाती है। इस तरह के अंतराल की घटना का समय व्यक्तिगत पौधों के लिए काफी स्थिर मूल्य है। सेब के पेड़ में, यह जीवन के सातवें वर्ष में होता है, हॉर्नबीम में - पचासवें में। कुछ प्रजातियों में यह बिल्कुल नहीं होता है। छीलने का मुख्य कार्य यांत्रिक और थर्मल क्षति से सुरक्षा है।
पैरेन्काइमा
यह विशिष्ट ऊतकों का एक समूह है जो प्रवाहकीय और यांत्रिक ऊतकों के बीच पौधे के शरीर के अंदर रिक्त स्थान को भरता है। (चित्र 8)। अधिक बार पैरेन्काइमा कोशिकाओं का एक गोल आकार होता है, कम अक्सर एक लम्बी आकृति। विकसित अंतरकोशिकीय स्थानों की उपस्थिति विशेषता है। कोशिकाओं के बीच के रिक्त स्थान मिलकर एक परिवहन तंत्र का निर्माण करते हैं - एपोप्लास्टइसके अलावा, इंटरसेलुलर रिक्त स्थान पौधे की "वेंटिलेशन सिस्टम" बनाते हैं। रंध्रों, या मसूर के माध्यम से, वे वायुमंडलीय हवा से जुड़े होते हैं और पौधे के अंदर एक इष्टतम गैस संरचना प्रदान करते हैं। जलभराव वाली मिट्टी पर उगने वाले पौधों के लिए विकसित अंतरकोशिकीय स्थान विशेष रूप से आवश्यक होते हैं, जहां सामान्य गैस विनिमय मुश्किल होता है। इस पैरेन्काइमा को कहा जाता है aerenchyma(चित्र 9)।
पैरेन्काइमा के तत्व, अन्य ऊतकों के बीच अंतराल को भरते हुए, एक समर्थन का कार्य भी करते हैं। पैरेन्काइमा कोशिकाएं जीवित होती हैं, उनमें स्क्लेरेन्काइमा की तरह मोटी कोशिका भित्ति नहीं होती है। इसलिए, यांत्रिक गुण टर्गर द्वारा प्रदान किए जाते हैं। यदि पानी की मात्रा कम हो जाती है, जिससे पौधे का प्लास्मोलिसिस और विल्टिंग हो जाता है।
एसिमिलेशन पैरेन्काइमाकई अंतरकोशिकीय स्थानों के साथ पतली दीवार वाली कोशिकाओं द्वारा निर्मित। इस संरचना की कोशिकाओं में कई क्लोरोप्लास्ट होते हैं, इसलिए इसे कहा जाता है क्लोरेन्काइमा. क्लोरोप्लास्ट एक दूसरे को छायांकित किए बिना दीवार के साथ स्थित होते हैं। एसिमिलेशन पैरेन्काइमा में, प्रकाश संश्लेषण प्रतिक्रियाएं होती हैं, जो पौधे को कार्बनिक पदार्थ और ऊर्जा प्रदान करती हैं। प्रकाश संश्लेषक प्रक्रियाओं का परिणाम पृथ्वी पर सभी जीवित जीवों के अस्तित्व की संभावना है।
आत्मसात ऊतक केवल पौधे के प्रबुद्ध भागों में मौजूद होते हैं, वे एक पारदर्शी एपिडर्मिस द्वारा पर्यावरण से अलग होते हैं। यदि एपिडर्मिस को अपारदर्शी द्वितीयक पूर्णांक ऊतकों द्वारा प्रतिस्थापित किया जाता है, तो एसिमिलेशन पैरेन्काइमा गायब हो जाता है।
भंडारण पैरेन्काइमाकार्बनिक पदार्थों के भंडार के रूप में कार्य करता है जो अस्थायी रूप से पौधे जीव द्वारा उपयोग नहीं किया जाता है। सिद्धांत रूप में, जीवित प्रोटोप्लास्ट वाली कोई भी कोशिका विभिन्न प्रकार के समावेशन के रूप में कार्बनिक पदार्थ जमा करने में सक्षम है, लेकिन कुछ कोशिकाएँ इसमें विशेषज्ञ होती हैं। (चित्र 10, 11)। ऊर्जा से भरपूर यौगिकों को केवल बढ़ते मौसम के दौरान जमा किया जाता है, सुप्त अवधि के दौरान और अगले बढ़ते मौसम की तैयारी में खपत होती है। इसलिए, आरक्षित पदार्थ केवल बारहमासी पौधों में वनस्पति अंगों में जमा होते हैं।
शेयरों का भंडार सामान्य अंग (पलायन, जड़), साथ ही विशेष वाले (राइजोम, कंद, बल्ब) हो सकते हैं। सभी बीज पौधे बीजों (बीजपत्र, एंडोस्पर्म) में ऊर्जावान रूप से मूल्यवान पदार्थों को संग्रहित करते हैं। शुष्क जलवायु में कई पौधे न केवल कार्बनिक पदार्थ, बल्कि पानी भी जमा करते हैं। (चित्र 12)। उदाहरण के लिए, मुसब्बर मांसल पत्तियों में पानी जमा करता है, शूटिंग में कैक्टि।
यांत्रिक कपड़े
पादप कोशिकाओं के यांत्रिक गुणों द्वारा प्रदान किया जाता है:
कोशिका का कठोर खोल;
स्फीति, अर्थात् कोशिकाओं की स्फीति अवस्था।
इस तथ्य के बावजूद कि लगभग सभी ऊतक कोशिकाओं में यांत्रिक गुण होते हैं, हालांकि, पौधे में ऐसे ऊतक होते हैं जिनके लिए यांत्रिक गुण बुनियादी होते हैं। यह कोलेनकाइमाऔर स्क्लेरेनकाइमा. वे आमतौर पर अन्य ऊतकों के साथ बातचीत में कार्य करते हैं। शरीर के अंदर पौधे एक प्रकार का ढाँचा बनाते हैं। इसलिए उन्हें कहा जाता है मजबूत.
सभी पौधों में समान रूप से स्पष्ट यांत्रिक ऊतक नहीं होते हैं। जलीय वातावरण में रहने वाले पौधों को स्थलीय की तुलना में बहुत कम आंतरिक समर्थन की आवश्यकता होती है। इसका कारण यह है कि जलीय पौधों को कम आंतरिक समर्थन की आवश्यकता होती है। उनका शरीर काफी हद तक आसपास के पानी से समर्थित है। जमीन पर हवा इस तरह का समर्थन नहीं बनाती है, क्योंकि इसमें पानी की तुलना में कम घनत्व होता है। यही कारण है कि विशिष्ट यांत्रिक ऊतकों की उपस्थिति प्रासंगिक हो जाती है।
विकास की प्रक्रिया में आंतरिक समर्थन संरचनाओं में सुधार हुआ।
Collenchyma।यह केवल जीवित कोशिकाओं द्वारा बनता है, जो अंग की धुरी के साथ लम्बी होती हैं। प्राथमिक वृद्धि की अवधि के दौरान इस प्रकार के यांत्रिक ऊतक बहुत जल्दी बनते हैं। इसलिए, यह महत्वपूर्ण है कि कोशिकाएं जीवित रहें और पास की खींची हुई कोशिकाओं के साथ-साथ फैलने की क्षमता बनाए रखें।
कोलेन्काइमा कोशिकाओं की विशेषताएं:
खोल का असमान मोटा होना, जिसके परिणामस्वरूप इसके कुछ हिस्से पतले रहते हैं, जबकि अन्य मोटे होते हैं;
गोले लिग्निफाई नहीं करते हैं।
Collenchyma कोशिकाएं एक दूसरे के सापेक्ष अलग-अलग स्थित होती हैं। आसन्न कोशिकाओं में, एक दूसरे के सामने वाले कोनों पर गाढ़ापन बनता है ( चावल। 13 ). इस कॉलेन्काइमा को कहा जाता है कोना।दूसरे मामले में, कोशिकाओं को समानांतर परतों में व्यवस्थित किया जाता है। इन परतों का सामना करने वाली कोशिका झिल्लियाँ अत्यधिक मोटी होती हैं। यह लैमेलर कोलेन्काइमा।प्रचुर मात्रा में अंतरकोशिकीय स्थानों के साथ कोशिकाओं को शिथिल रूप से व्यवस्थित किया जा सकता है ढीला Collenchyma है।इस तरह के कोलेंकाइमा अक्सर जलभराव वाली मिट्टी पर पौधों में पाए जाते हैं।
Collenchyma युवा पौधों, जड़ी-बूटियों के रूपों, और पौधों के उन हिस्सों में भी विशेष महत्व रखता है जहाँ द्वितीयक वृद्धि नहीं होती है, जैसे कि पत्तियाँ। इस मामले में, इसे सतह के बहुत करीब रखा जाता है, कभी-कभी सिर्फ एपिडर्मिस के नीचे। यदि अंग के चेहरे हैं, तो उनके शिखर के साथ कोलेन्काइमा की मोटी परतें पाई जाती हैं।
Collenchyma कोशिकाएं केवल स्फीति की उपस्थिति में क्रियाशील होती हैं। पानी की कमी से कॉलेंकाइमा की प्रभावशीलता कम हो जाती है और पौधा अस्थायी रूप से मुरझा जाता है, उदाहरण के लिए, ककड़ी के पत्ते गर्म दिन में गिर जाते हैं। कोशिकाओं को पानी से भरने के बाद, कोलेंकाइमा के कार्य बहाल हो जाते हैं।
दृढ़ोतक।दूसरे प्रकार के यांत्रिक ऊतक। Collenchyma के विपरीत, जहाँ सभी कोशिकाएँ जीवित हैं, स्क्लेरेन्काइमा कोशिकाएँ मृत हैं। इनकी दीवारें बहुत मोटी होती हैं। वे एक यांत्रिक कार्य करते हैं। झिल्ली का एक मजबूत मोटा होना पदार्थों के परिवहन का उल्लंघन करता है, जिसके परिणामस्वरूप प्रोटोप्लास्ट मर जाता है ( चावल। 14 ). स्क्लेरेन्काइमा की कोशिका झिल्लियों का लिग्निफिकेशन तब होता है जब पौधे का अंग पहले ही अपनी वृद्धि पूरी कर चुका होता है। इसलिए, वे अब आसपास के ऊतकों के खिंचाव में हस्तक्षेप नहीं करते हैं।
आकार के आधार पर, दो प्रकार की स्क्लेरेन्काइमा कोशिकाएं प्रतिष्ठित होती हैं - तंतु और स्केलेरिड।
फाइबरबहुत मोटी दीवारों और एक छोटी गुहा के साथ एक मजबूत लम्बी आकृति है। वे लकड़ी के तंतुओं से कुछ छोटे होते हैं। अक्सर एपिडर्मिस के नीचे अनुदैर्ध्य परतें और किस्में बनती हैं। फ्लोएम या जाइलम में ये अकेले या समूहों में पाए जाते हैं। फ्लोएम में इन्हें कहते हैं बस्ट फाइबर(चावल। 15 ), और जाइलम में लाइब्रिफॉर्म फाइबर(चावल। 16 ).
स्केलेरिड्स,या पथरीली कोशिकाएँ, शक्तिशाली गोले के साथ गोल या शाखित कोशिकाओं द्वारा दर्शाए जाते हैं। एक पौधे के शरीर में, वे अकेले (सहायक कोशिकाओं) में पाए जा सकते हैं ( अंजीर.17 ) या समूह (चित्र 18)। यह ध्यान दिया जाना चाहिए कि यांत्रिक गुण दृढ़ता से स्केलेरिड्स के स्थान पर निर्भर करते हैं। स्क्लेराइड्स का हिस्सा निरंतर परतें बनाता है, उदाहरण के लिए, नट के खोल में या फलों के पत्थरों (पत्थर के फल) में।
उत्सर्जक ऊतक
किसी भी जीवित कोशिका में प्रारंभ में पदार्थों को हटाने का कार्य होता है, जबकि परिवहन सांद्रता प्रवणता के साथ और ढाल के विरुद्ध दोनों जा सकता है। पदार्थ जो कोशिका में संश्लेषित किए गए हैं उन्हें हटाया जा सकता है और बाद में अन्य कोशिकाओं (फाइटोहोर्मोन, एंजाइम) की गतिविधि को प्रभावित करेगा। ऐसी प्रक्रिया कहलाती है स्राव. मामले में जब पदार्थ जो कोशिका के अपशिष्ट उत्पाद हैं, उत्सर्जित होते हैं, इस प्रक्रिया को कहा जाता है मलत्याग. इस तथ्य के बावजूद कि पौधों और जानवरों की कोशिकाओं में पदार्थों के उत्सर्जन में मौलिक समानता है, पौधों में कई विशेषताएं हैं जो महत्वपूर्ण गतिविधि में मौलिक अंतर से उत्पन्न होती हैं। पौधों में चयापचय प्रतिक्रियाओं का स्तर जानवरों की तुलना में बहुत कम है। इसलिए, आनुपातिक रूप से कम मात्रा में कचरा उत्सर्जित होता है। एक अन्य विशेषता यह है कि, लगभग सभी आवश्यक कार्बनिक यौगिकों को स्वतंत्र रूप से संश्लेषित करते हुए, पौधे कभी भी उनका अत्यधिक भंडार नहीं बनाते हैं। जारी किए गए पदार्थ अन्य प्रतिक्रियाओं के लिए प्रारंभिक सामग्री के रूप में काम कर सकते हैं (उदाहरण के लिए, सीओ 2 और एच 2 ओ)।
यदि जानवरों में विषाक्त पदार्थों के उत्सर्जन की प्रक्रिया शरीर से उनके निष्कासन से जुड़ी होती है, तो पौधे ऐसा नहीं कर सकते हैं, जीवित प्रोटोप्लास्ट में अनावश्यक पदार्थों को अलग करना (वैक्यूल में विभिन्न पदार्थों को हटाना), मृत कोशिकाओं में (एक बारहमासी के अधिकांश ऊतक) पौधे में ऐसी कोशिकाएँ होती हैं), अंतरकोशिकीय स्थानों में।
उत्सर्जित पदार्थों को दो बड़े समूहों में विभाजित किया जा सकता है। पहला समूह जैविक पदार्थ है जो सीधे कोशिका द्वारा संश्लेषित होता है (एंजाइम, पॉलीसेकेराइड, लिग्निन, टेरपेन, बाद वाले आवश्यक तेलों और रेजिन के घटक तत्व हैं)। दूसरा समूह - पदार्थ जो शुरू में जाइलम या फ्लोएम परिवहन (पानी, खनिज लवण, अमीनो एसिड, मोनोसैकराइड, आदि) की मदद से बाहर से कोशिका में प्रवेश करते हैं। उत्सर्जित रहस्य शायद ही कभी सजातीय होता है और आमतौर पर एक मिश्रण होता है जिसमें एक पदार्थ में उच्चतम एकाग्रता होती है।
जंतुओं के विपरीत पौधों में पूर्ण उत्सर्जन तंत्र नहीं होता है। पूरे संयंत्र में बिखरी हुई केवल विशेष संरचनाएं हैं - idioblasts.उन्हें बनाने वाली कोशिकाएँ पास में पड़ी पैरेन्काइमा कोशिकाओं से छोटी होती हैं। उनके पास एक इलेक्ट्रॉन-सघन साइटोप्लाज्म है, जिसमें एंडोप्लाज्मिक रेटिकुलम और गोल्गी कॉम्प्लेक्स के विकसित तत्व हैं। अक्सर व्यक्त नहीं किया जाता है, एक केंद्रीय रिक्तिका होती है। अपने और अन्य जीवित कोशिकाओं के बीच, ये कोशिकाएँ असंख्य प्लास्मोडेस्माटा द्वारा जुड़ी होती हैं।
स्थान के आधार पर उत्सर्जक ऊतक हो सकते हैं बाहरी और आंतरिक।
बाहरी उत्सर्जक ऊतक।ग्रंथियों के बालडेरिवेटिव हैं एपिडर्मिस. Morphologically, वे परिवर्तनशील हैं - उनके पास एक बहुकोशिकीय सिर हो सकता है, लम्बी हो सकती है, एक पैर पर ढाल के रूप में, आदि। (अंजीर। 19)। इनमें चुभने वाले बिछुआ बाल शामिल हैं।
हाइडेथोड्स- ये ऐसी संरचनाएं हैं जो कम वाष्पोत्सर्जन और उच्च आर्द्रता की स्थितियों में अतिरिक्त पानी छोड़ती हैं - कण्ठ. ये संरचनाएं पतली दीवारों वाली रंगहीन जीवित कोशिकाओं के समूहों द्वारा बनाई जाती हैं - उपमा।यह ऊतक प्रवाहकीय बंडल (चित्र 20) के निकट है। पानी को विशेष जल रंध्रों के माध्यम से छोड़ा जाता है, जो सामान्य रंध्रों से गतिहीनता और लगातार खुले अंतराल से भिन्न होते हैं। गुटेशन द्रव की संरचना लगभग शुद्ध पानी से लेकर पदार्थों के बहुत जटिल मिश्रण तक व्यापक रूप से भिन्न होती है।
अमृत।कई पौधों को 7 से 87% मोनो- और ओलिगोसेकेराइड युक्त तरल की रिहाई की विशेषता है। यह प्रक्रिया विशेष संरचनाओं - अमृत द्वारा की जाती है। स्थान के आधार पर, वहाँ हैं फूल,या फूलों, अमृत (चित्र 21), साथ ही तनों, पत्तियों पर स्थित पौधे - एक्स्ट्राफ्लोरा, या एक्स्ट्राफ्लोरल अमृत।अमृत प्रतिनिधित्व कर सकता है यह एक अपरिवर्तित फ्लोएम रस है, जो अंतरकोशिकीय स्थानों के माध्यम से सतह पर पहुंचाया जाता है और रंध्रों के माध्यम से उत्सर्जित होता है। ग्रंथियों के पैरेन्काइमा द्वारा अधिक जटिल अमृत बनते हैं, जो ग्रंथियों के बालों के साथ एपिडर्मिस से ढके होते हैं। अमृत या तो एपिडर्मल कोशिकाओं या ग्रंथियों के बालों द्वारा उत्सर्जित होता है। इस मामले में, स्रावित अमृत फ्लोएम सैप से भिन्न होता है। चूंकि इसमें ग्लूकोज, फ्रुक्टोज, सुक्रोज और फ्लोएम जूस - ग्लूकोज का प्रभुत्व है। आयन कम सांद्रता में मौजूद हैं। आयन कम सांद्रता में मौजूद हैं। परागणकों को आकर्षित करने के लिए अमृत में स्टेरॉयड हार्मोन हो सकते हैं, जो कीड़ों के लिए आवश्यक हैं।
पाचन ग्रंथियांकीटभक्षी पौधों (ओस, नेपेंटिस, आदि) में मौजूद। छोटे अकशेरूकीय के स्पर्श के जवाब में, वे हाइड्रोलाइटिक एंजाइम, फॉर्मिक, ब्यूटिरिक और प्रोपियोनिक एसिड युक्त एक रहस्य का स्राव करते हैं। नतीजतन, बायोपॉलिमर्स मोनोमर्स में टूट जाते हैं, जिनका उपयोग पौधे अपनी जरूरतों के लिए करते हैं।
नमक ग्रंथियांखारी मिट्टी पर उगने वाले पौधों में विकसित होते हैं। ये ग्रंथियां पत्तियों पर स्थित होती हैं और घुले हुए रूप में, अतिरिक्त आयन सतह पर लाए जाते हैं। नमक पहले छल्ली पर जमा होता है, फिर इसे बारिश से धोया जाता है (चित्र 22)। इसके अलावा, कई पौधों की पत्तियों पर होता है नमक बाल।प्रत्येक बाल में दो कोशिकाएँ होती हैं: एक सिर बनाती है, दूसरी - तना। सिर की कोशिका की रसधानी में लवण जमा हो जाते हैं, और जब उनकी सघनता अधिक हो जाती है, तो सिर गिर जाता है। इसके स्थान पर एक नई कोशिका विकसित होती है, जो लवण भी जमा करती है। पत्ती के विकास के दौरान, सिर गिर सकता है और कई बार बन सकता है। नमक बालों का लाभ यह है कि नमक छोड़ने से वे थोड़ा पानी खो देते हैं। खारी मिट्टी पर उगने वाले पौधों के लिए यह घटना बहुत महत्वपूर्ण है, जहां ताजे पानी की कमी है।
आंतरिक उत्सर्जक ऊतक।इडियोब्लास्ट्स के रूप में पौधे के पूरे शरीर में बिखरी हुई आंतरिक उत्सर्जन संरचनाएं, शरीर के बाहर पदार्थों को नहीं हटाती हैं, उन्हें स्वयं में जमा करती हैं। इडियोब्लास्ट पादप कोशिकाएं हैं जो एक ही ऊतक में अन्य कोशिकाओं से आकार, संरचना या सामग्री में भिन्न होती हैं।उत्पत्ति के आधार पर, स्रावित पदार्थों के स्किज़ोजेनिक (ग्रीक स्किसो - स्प्लिट) और लाइसोजेनिक (ग्रीक लिसीज़ - विघटन) रिसेप्टेकल्स को प्रतिष्ठित किया जाता है (चित्र 23)।
स्किज़ोजेनिक रिसेप्टेकल्स।ये कमोबेश व्यापक अंतरकोशिकीय स्थान हैं जो उत्सर्जित पदार्थों से भरे हुए हैं। उदाहरण के लिए, कोनिफर्स, अरालिया, छाता और अन्य के राल मार्ग। यह माना जाता है कि राल में जीवाणुनाशक गुण होते हैं, शाकाहारी जीवों को पीछे हटाते हैं, जिससे पौधे उनके लिए अखाद्य हो जाते हैं।
लाइसिजेनिक रिसेप्टेकल्सजीवित कोशिकाओं के स्थान पर बनते हैं जो मर जाती हैं और उनमें पदार्थों के जमा होने के बाद नष्ट हो जाती हैं। खट्टे फलों के छिलके में इस तरह की संरचनाएँ देखी जा सकती हैं।
मुख्य प्रकार के अलावा, मध्यवर्ती रूप भी हैं। दूध का- दो प्रकार हैं: संयुक्तऔर गैर खंडित किया. पूर्व का निर्माण जंजीरों में व्यवस्थित कई जीवित कोशिकाओं से होता है। संपर्क के बिंदु पर, गोले नष्ट हो जाते हैं, प्रोटोप्लास्ट विलीन हो जाते हैं, और परिणामस्वरूप एक एकल नेटवर्क बनता है। ऐसे दूधिया कम्पोजिट में पाए जाते हैं। गैर खंडित कियालैक्टिफेर्स एक विशाल बहुकेन्द्रीय कोशिका द्वारा निर्मित होते हैं। यह विकास के प्रारंभिक चरण में उत्पन्न होता है, लंबा होता है और पौधे के बढ़ने पर शाखाएं, एक जटिल नेटवर्क बनाते हैं, उदाहरण के लिए, यूफोरबिया पौधों (चित्र 25) में।
लैक्टिफेरस की कोशिका भित्ति अत्यधिक जलयुक्त होती है। वे लिग्निफाई नहीं करते हैं और इसलिए प्लास्टिक हैं। लैक्टिस के मध्य भाग में लेटेक्स के साथ एक रिक्तिका होती है, और प्रोटोप्लास्ट एक निकट-दीवार की स्थिति में होता है। प्रोटोप्लास्ट और रसधानी के बीच की सीमा हमेशा निर्धारित नहीं की जा सकती है। लैक्टिफेर मेरिस्टेम में प्रवेश करते हैं और पौधे के अंग के साथ बढ़ते हैं।
प्रवाहकीय ऊतक
इस प्रकार का है कठिनऊतक, अलग-अलग विभेदित कोशिकाओं के होते हैं। वास्तविक प्रवाहकीय तत्वों के अलावा, ऊतक में यांत्रिक, उत्सर्जन और भंडारण तत्व होते हैं (चित्र 26)। प्रवाहकीय ऊतक सभी पौधों के अंगों को एक प्रणाली में एकजुट करते हैं। प्रवाहकीय ऊतक दो प्रकार के होते हैं: जाइलमऔर फ्लाएम(ग्रीक ज़ाइलॉन - पेड़; फ्लोयोस - छाल, बस्ट)। उनके पास संरचनात्मक और कार्यात्मक दोनों अंतर हैं।
जाइलम के संवाहक तत्व मृत कोशिकाओं द्वारा बनते हैं। वे जड़ से पत्तियों तक पानी और उसमें घुले पदार्थों का लंबी दूरी तक परिवहन करते हैं। फ्लोएम के प्रवाहकीय तत्व प्रोटोप्लास्ट को जीवित रखते हैं। वे प्रकाश संश्लेषक पत्तियों से जड़ तक लंबी दूरी का परिवहन करते हैं।
आमतौर पर जाइलम और फ्लोएम एक निश्चित क्रम में पौधे के शरीर में स्थित होते हैं, जिससे परतें बनती हैं या प्रवाहकीय बंडल. संरचना के आधार पर, कई प्रकार के संवाहक बंडलों को प्रतिष्ठित किया जाता है, जो पौधों के कुछ समूहों की विशेषता है। संपार्श्विक खुले बंडल मेंजाइलम और फ्लोएम के बीच कैम्बियम होता है, जो द्वितीयक वृद्धि प्रदान करता है (चित्र 27-ए, 28)। द्विसंपार्श्विक खुले बंडल मेंफ्लोएम जाइलम के सापेक्ष दोनों तरफ स्थित होता है (चित्र 27-बी, 29)। बंद बंडलकैम्बियम नहीं होता है, और इसलिए द्वितीयक गाढ़ा करने में सक्षम नहीं होते हैं (चित्र 27-बी, 27-डी, 30.31)। दो और प्रकार हैं गाढ़ा बंडल,जहां या तो फ्लोएम जाइलम को घेरता है (चित्र 27-ई, 32), या जाइलम फ्लोएम को घेरता है (चित्र 27-ई)।
जाइलम (लकड़ी)। उच्च पौधों में जाइलम का विकास जल विनिमय के प्रावधान से जुड़ा हुआ है। चूंकि एपिडर्मिस के माध्यम से पानी लगातार उत्सर्जित होता है, नमी की समान मात्रा को पौधे द्वारा अवशोषित किया जाना चाहिए और उन अंगों में जोड़ा जाना चाहिए जो वाष्पोत्सर्जन करते हैं। यह ध्यान में रखा जाना चाहिए कि जल-संचालन कोशिकाओं में एक जीवित प्रोटोप्लास्ट की उपस्थिति परिवहन को बहुत धीमा कर देगी, और मृत कोशिकाएं यहां अधिक कार्यात्मक हैं। हालाँकि, एक मृत कोशिका नहीं होती है सूजन के साथ इसलिए, खोल में यांत्रिक गुण होने चाहिए। टिप्पणी: मरोड़ - पौधों की कोशिकाओं, ऊतकों और अंगों की स्थिति जिसमें वे अपनी लोचदार झिल्लियों पर कोशिकाओं की सामग्री के दबाव के कारण लोचदार हो जाते हैं।दरअसल, जाइलम के संवाहक तत्वों में मोटे लिग्निफाइड गोले के साथ अंग की धुरी के साथ फैली हुई मृत कोशिकाएं होती हैं।
प्रारंभ में, जाइलम प्राथमिक मेरिस्टेम - प्रोकैम्बियम से बनता है, जो अक्षीय अंगों के शीर्ष पर स्थित होता है। पहले भेद करता है प्रोटोजाइलम,तब मेटाजाइलम।तीन प्रकार के जाइलम गठन ज्ञात हैं। पर एक्ज़कप्रकार, प्रोटोजाइलम तत्व पहले प्रोकैम्बियल बंडल की परिधि पर दिखाई देते हैं, फिर मेटाजाइलम तत्व केंद्र में दिखाई देते हैं। यदि प्रक्रिया विपरीत दिशा में जाती है (अर्थात केंद्र से परिधि तक), तो यह endarchicप्रकार। पर मेसार्च प्रकारप्रोकैम्बियल बंडल के केंद्र में जाइलम रखा जाता है, जिसके बाद इसे केंद्र की ओर और परिधि की ओर जमा किया जाता है।
जड़ की विशेषता एक एक्सार्क प्रकार के जाइलम गठन से होती है, जबकि तनों में एक एंडार्क प्रकार होता है। कम संगठित पौधों में, जाइलम निर्माण के तरीके बहुत विविध होते हैं और व्यवस्थित विशेषताओं के रूप में काम कर सकते हैं।
कुछ पौधों में (उदाहरण के लिए, मोनोकॉट्स), सभी प्रकोष्ठीय कोशिकाएं प्रवाहकीय ऊतकों में अंतर करती हैं जो द्वितीयक गाढ़ा करने में सक्षम नहीं होते हैं। अन्य रूपों में (उदाहरण के लिए, वुडी), पार्श्व विभज्योतक (केंबियम) जाइलम और फ्लोएम के बीच रहते हैं। ये कोशिकाएँ जाइलम और फ्लोएम को विभाजित करने, नवीनीकृत करने में सक्षम हैं। ऐसी प्रक्रिया कहलाती है द्वितीयक वृद्धि।अपेक्षाकृत स्थिर जलवायु परिस्थितियों में उगने वाले कई पौधे लगातार बढ़ते हैं। मौसमी जलवायु परिवर्तन के अनुकूल रूपों में - समय-समय पर। नतीजतन, अच्छी तरह से परिभाषित वार्षिक वृद्धि के छल्ले बनते हैं।
प्रकोष्ठीय कोशिकाओं के विभेदन के मुख्य चरण। इसकी कोशिकाओं में पतली झिल्लियां होती हैं जो अंग के विकास के दौरान उन्हें फैलने से नहीं रोकती हैं। प्रोटोप्लास्ट तब द्वितीयक झिल्ली बिछाना शुरू कर देता है। लेकिन इस प्रक्रिया में स्पष्ट विशेषताएं हैं। द्वितीयक झिल्ली एक सतत परत में जमा नहीं होती है, जो कोशिका को फैलने नहीं देती, बल्कि छल्ले या सर्पिल के रूप में होती है। सेल बढ़ाव मुश्किल नहीं है। युवा कोशिकाओं में, हेलिक्स के छल्ले या कुंडली एक दूसरे के करीब स्थित होते हैं। परिपक्व कोशिकाओं में, वे कोशिका खिंचाव (चित्र 33) के परिणामस्वरूप विचलन करते हैं। खोल के चक्राकार और सर्पिल मोटा होना विकास को नहीं रोकता है, हालांकि, यंत्रवत् वे गोले से नीच होते हैं, जहां द्वितीयक मोटा होना एक सतत परत बनाता है। इसलिए, जाइलम में वृद्धि की समाप्ति के बाद, एक सतत लिग्नीफाइड खोल वाले तत्व बनते हैं ( मेटाजाइलम). यह ध्यान दिया जाना चाहिए कि यहां द्वितीयक मोटा होना कुंडलाकार या सर्पिल नहीं है, बल्कि पंचर, सीढ़ी जैसा, जालीदार (चित्र 34) है। इसकी कोशिकाएं खिंचने में सक्षम नहीं होती हैं और कुछ ही घंटों में मर जाती हैं। आस-पास की कोशिकाओं में यह प्रक्रिया समन्वित तरीके से होती है। साइटोप्लाज्म में बड़ी संख्या में लाइसोसोम दिखाई देते हैं। फिर वे विघटित हो जाते हैं, और उनमें मौजूद एंजाइम प्रोटोप्लास्ट को नष्ट कर देते हैं। जब अनुप्रस्थ दीवारें नष्ट हो जाती हैं, तो एक के ऊपर एक श्रृंखला में स्थित कोशिकाएं एक खोखले बर्तन (चित्र 35) का निर्माण करती हैं। अधिकांश एंजियोस्पर्म और कुछ फ़र्न में वाहिकाएँ होती हैं।
एक प्रवाहकीय कोशिका जो इसकी दीवार में छिद्रों के माध्यम से नहीं बनती है, कहलाती है ट्रेकिड।वाहिकाओं के माध्यम से ट्रेकिड्स के माध्यम से पानी की गति धीमी होती है। तथ्य यह है कि ट्रेकिड्स में प्राथमिक झिल्ली कहीं भी बाधित नहीं होती है। ट्रेकिड्स एक दूसरे के माध्यम से संवाद करते हैं तब से।यह स्पष्ट किया जाना चाहिए कि पौधों में, छिद्र प्राथमिक झिल्ली तक द्वितीयक झिल्ली में केवल एक अवसाद है, और ट्रेकिड्स के बीच छिद्रों के माध्यम से नहीं होते हैं।
सबसे आम बॉर्डर वाले छिद्र हैं (चित्र 35-1)। उनका चैनल, कोशिका की गुहा का सामना करते हुए, एक विस्तार बनाता है - ताकना कक्ष।प्राथमिक खोल पर अधिकांश शंकुधारी पौधों के छिद्रों में एक मोटापन होता है - टोरस,जो एक प्रकार का वाल्व है और जल परिवहन की तीव्रता को नियंत्रित करने में सक्षम है। स्थानांतरित करके, टोरस छिद्र के माध्यम से पानी के प्रवाह को अवरुद्ध करता है, लेकिन उसके बाद यह एक बार की कार्रवाई करते हुए अपनी पिछली स्थिति में वापस नहीं आ सकता है।
छिद्र अधिक या कम गोल होते हैं, लम्बी धुरी के लंबवत होते हैं (इन छिद्रों का एक समूह सीढ़ी जैसा दिखता है, इसलिए इस छिद्र को सीढ़ी छिद्र कहा जाता है)। छिद्रों के माध्यम से परिवहन अनुदैर्ध्य और अनुप्रस्थ दोनों दिशाओं में किया जाता है। छिद्र न केवल ट्रेकिड्स में मौजूद होते हैं, बल्कि एक पोत बनाने वाली व्यक्तिगत संवहनी कोशिकाओं में भी मौजूद होते हैं।
विकासवादी सिद्धांत के दृष्टिकोण से, ट्रेकिड्स पहली और मुख्य संरचना है जो उच्च पौधों के शरीर में पानी का संचालन करती है। ऐसा माना जाता है कि जहाजों की उत्पत्ति उनके बीच की अनुप्रस्थ दीवारों (चित्र 36) के लसीका के कारण ट्रेकिड्स से हुई थी। अधिकांश फ़र्न और जिम्नोस्पर्म में नहीं होता है। वे ट्रेकिड्स के माध्यम से पानी ले जाते हैं।
विकासवादी विकास की प्रक्रिया में, पौधों के विभिन्न समूहों में जहाजों का बार-बार उदय हुआ, लेकिन उन्होंने एंजियोस्पर्म में सबसे महत्वपूर्ण कार्यात्मक महत्व प्राप्त किया, जिसमें वे ट्रेकिड्स के साथ हैं। यह माना जाता है कि परिवहन के अधिक उन्नत तंत्र के कब्जे ने उन्हें न केवल जीवित रहने में मदद की, बल्कि महत्वपूर्ण रूपों को प्राप्त करने में भी मदद की।
जाइलम एक जटिल ऊतक है, इसमें जल-संवाहक तत्वों के अलावा अन्य तत्व भी होते हैं। यांत्रिक कार्य लाइब्रिफॉर्म फाइबर द्वारा किए जाते हैं ( अव्यक्त।मुक्ति - लब, रूप - रूप)। अतिरिक्त यांत्रिक संरचनाओं की उपस्थिति महत्वपूर्ण है, क्योंकि पानी ले जाने वाले तत्वों की दीवारें मोटी होने के बावजूद अभी भी बहुत पतली हैं। वे स्वतंत्र रूप से एक बारहमासी पौधे के बड़े द्रव्यमान को धारण करने में सक्षम नहीं हैं। ट्रेकिड्स से तंतु विकसित होते हैं। वे छोटे आकार, लिग्नीफाइड (लिग्नीफाइड) गोले और संकीर्ण गुहाओं की विशेषता हैं। दीवार पर किनारों से रहित छिद्र पाए जा सकते हैं। ये रेशे पानी का संचालन नहीं कर सकते, इनका मुख्य कार्य आधार है।
जाइलम में जीवित कोशिकाएं भी होती हैं। उनका द्रव्यमान लकड़ी की कुल मात्रा का 25% तक पहुंच सकता है। चूँकि इन कोशिकाओं का आकार गोल होता है, इसलिए इन्हें काष्ठ पैरेन्काइमा कहा जाता है। पैरेन्काइमा पौधे के शरीर में दो तरह से स्थित होता है। पहले मामले में, कोशिकाओं को लंबवत तारों के रूप में व्यवस्थित किया जाता है - यह है किनारा पैरेन्काइमा. एक अन्य मामले में, पैरेन्काइमा क्षैतिज किरणें बनाता है। उन्हें बुलाया जाता है कोर किरणें, क्योंकि वे कोर और छाल को जोड़ते हैं। कोर पदार्थों के भंडारण सहित कई कार्य करता है।
फ्लोएम (बस्ट)। यह एक जटिल ऊतक है, क्योंकि यह विषम कोशिकाओं द्वारा बनता है। मुख्य संवाहक कोशिका कहलाती है चलनी तत्व(चित्र 37)। जाइलम के संवाहक तत्व मृत कोशिकाओं द्वारा बनते हैं, जबकि फ्लोएम में वे कार्य करने की अवधि के दौरान जीवित रहते हैं, हालांकि बहुत बदल जाते हैं, प्रोटोप्लास्ट। फ्लोएम प्रकाश संश्लेषक अंगों से प्लास्टिक पदार्थों का बहिर्वाह करता है। पौधों की सभी जीवित कोशिकाओं में कार्बनिक पदार्थों के संचालन की क्षमता होती है। और इसलिए, यदि जाइलम केवल उच्च पौधों में ही पाया जा सकता है, तो कोशिकाओं के बीच कार्बनिक पदार्थों का परिवहन निचले पौधों में भी होता है।
जाइलम और फ्लोएम एपिकल मेरिस्टेम से विकसित होते हैं। पहले चरण में, प्रकोम्बियल कॉर्ड में, ए प्रोटोफ्लोएम।जैसे-जैसे आसपास के ऊतक बढ़ते हैं, यह फैलता है, और जब विकास पूरा हो जाता है, प्रोटोफ्लोएम के बजाय, यह बनता है रूपक।
उच्च पौधों के विभिन्न समूहों में दो प्रकार के छलनी तत्व पाए जा सकते हैं। फ़र्न और जिम्नोस्पर्म में, इसका प्रतिनिधित्व किया जाता है चलनी कोशिकाओं।कोशिकाओं में छलनी के क्षेत्र बगल की दीवारों के साथ बिखरे हुए हैं। प्रोटोप्लास्ट थोड़ा नष्ट हुए नाभिक को बरकरार रखता है।
एंजियोस्पर्म में चलनी तत्व कहलाते हैं चलनी ट्यूब।वे चलनी प्लेटों के माध्यम से एक दूसरे के साथ संवाद करते हैं। परिपक्व कोशिकाओं में केन्द्रक का अभाव होता है। हालांकि, छलनी ट्यूब के बगल में स्थित है उपग्रह सेल, जो सामान्य मातृ कोशिका (चित्र 38) के माइटोटिक विभाजन के परिणामस्वरूप छलनी ट्यूब के साथ मिलकर बनता है। साथी कोशिका में बड़ी संख्या में सक्रिय माइटोकॉन्ड्रिया के साथ एक सघन साइटोप्लाज्म होता है, साथ ही एक पूरी तरह से काम करने वाला नाभिक, बड़ी संख्या में प्लास्मोडेस्माटा (अन्य कोशिकाओं की तुलना में दस गुना अधिक)। साथी कोशिकाएं गैर-परमाणु चलनी ट्यूब कोशिकाओं की कार्यात्मक गतिविधि को प्रभावित करती हैं।
परिपक्व छलनी कोशिकाओं की संरचना में कुछ ख़ासियतें होती हैं। कोई रिक्तिका नहीं है, इसलिए साइटोप्लाज्म अत्यधिक द्रवीभूत होता है। केंद्रक अनुपस्थित हो सकता है (एंजियोस्पर्म में) या झुर्रीदार, कार्यात्मक रूप से निष्क्रिय अवस्था में हो सकता है। राइबोसोम और गोल्गी कॉम्प्लेक्स भी अनुपस्थित हैं, लेकिन एंडोप्लाज्मिक रेटिकुलम अच्छी तरह से विकसित है, जो न केवल साइटोप्लाज्म में प्रवेश करता है, बल्कि छलनी क्षेत्रों के छिद्रों के माध्यम से पड़ोसी कोशिकाओं में भी जाता है। अच्छी तरह से विकसित माइटोकॉन्ड्रिया और प्लास्टिड बहुतायत में पाए जाते हैं।
कोशिकाओं के बीच, पदार्थों का परिवहन कोशिका झिल्लियों पर स्थित छिद्रों से होता है। इस तरह के उद्घाटन को छिद्र कहा जाता है, लेकिन ट्रेकिड छिद्रों के विपरीत, वे होते हैं। यह माना जाता है कि वे अत्यधिक विस्तारित प्लास्मोडेस्माटा हैं, जिनकी दीवारों पर कॉलोस पॉलीसेकेराइड जमा होता है। छिद्रों को समूहों में व्यवस्थित किया जाता है, बनाते हैं छलनी के मैदान. आदिम रूपों में, छलनी के क्षेत्र बेतरतीब ढंग से खोल की पूरी सतह पर बिखरे हुए हैं; अधिक उन्नत एंजियोस्पर्म में, वे एक दूसरे से सटे पड़ोसी कोशिकाओं के सिरों पर स्थित होते हैं, जो बनाते हैं चलनी प्लेट(चित्र 39)। यदि उस पर एक छलनी का क्षेत्र है, तो इसे सरल कहा जाता है, यदि कई - जटिल हैं।
चलनी तत्वों के माध्यम से समाधानों की गति की गति 150 सेमी ∕ घंटे तक है। यह मुक्त प्रसार की दर का एक हजार गुना है। संभवतः, सक्रिय परिवहन होता है, और छलनी तत्वों और उपग्रह कोशिकाओं के कई माइटोकॉन्ड्रिया इसके लिए आवश्यक एटीपी की आपूर्ति करते हैं।
फ्लोएम के छलनी तत्वों का जीवन पार्श्व विभज्योतकों की उपस्थिति पर निर्भर करता है। यदि वे मौजूद हैं, तो छलनी तत्व पौधे के जीवन भर काम करते हैं।
छलनी तत्वों और उपग्रह कोशिकाओं के अलावा, फ्लोएम में बस्ट फाइबर, स्केलेरिड्स और पैरेन्काइमा होते हैं।
नियंत्रण और पुनरावृत्ति के लिए प्रश्न
उच्च पौधों के लक्षण? उच्च पौधों की किस पीढ़ी को गैमेटोफाइट कहा जाता है? स्पोरोफाइट?
वे कौन से कारण हैं जो स्थलीय पौधों में विभेदित ऊतकों के विकास में योगदान करते हैं।
पौधों के प्रकारों की सूची बनाइए।
आप विभज्योतकों के किन समूहों को जानते हैं? वे पौधे के शरीर में कहाँ स्थित हैं?
पूर्णांक ऊतक, उनके कार्य, महत्व।
विभिन्न प्रकार के पूर्णांक ऊतकों का संगठन।
रंध्र तंत्र की संरचना। आपको ज्ञात रंध्र उपकरण के प्रकारों की सूची बनाएं। रंध्र कैसे कार्य करते हैं?
पैरेन्काइमल ऊतकों की विशेषताएं, पैरेन्काइमा के प्रकार।
पैरेन्काइमा और स्क्लेरेन्काइमा के लक्षण।
पौधों की उत्सर्जी संरचनाओं के प्रकारों की सूची बनाइए। उनका क्या महत्व है?
उत्सर्जित पदार्थों के लिए पात्र बनाने की विधियाँ।
दूधवाले क्या होते हैं? वे किस प्रकार में विभाजित हैं?
प्रवाहकीय ऊतक कैसे व्यवस्थित होते हैं?
प्रवाहकीय बंडल क्या हैं? उनके प्रकार।
जाइलम में पदार्थों की गति की दिशा? फ्लोएम? क्या परिवहन किए गए पदार्थों की संरचना जाइलम और फ्लोएम में भिन्न होती है?
जाइलम, फ्लोएम की संरचना और कार्य।
विभिन्न व्यवस्थित समूहों से पौधों में फ्लोएम संवाहक तत्वों का संगठन।
विशिष्ट और जीवाणु, एक बाहरी कोशिका भित्ति होती है। उच्च कोशिकाओं में ऑर्गेनेल की संरचना और कार्य कोशिकाओं. जी. मेंडल नियंत्रण कार्य के नियम >>
प्रकार कपड़े. हमारे निकटतम रिश्तेदार, अन्य स्तनधारी, संरचनाऔर... , एक से मिलकर कोशिकाओं. पौधेइसमें विभिन्न ... सूक्ष्मनलिकाएं, सेंट्रीओल्स भी शामिल हैं कोशिका केंद्र. में सब्ज़ी कोशिकाओंक्लोरोप्लास्ट भी स्रावित होते हैं, जिनमें...
लाल रंग। में ऊतकों पौधेएंथ्रासीन डेरिवेटिव पाए जा सकते हैं ... एन्थ्रेसीन डेरिवेटिव औषधीय के रूप में सब्ज़ीकच्चे माल, और ... 2-3 से घिरे हैं, कम अक्सर 4 कोशिकाओंएपिडर्मिस। एककोशिकीय बाल, ... यौगिक, आधारित इमारतोंजिनमें से एन्थ्रेसीन की संरचना...