הזכרנו שוב ושוב שאחד או אחר מהחוקרים הרשומים הבחין בגרעינים בתאים.
מאחר שבעבודתו של שליידן, אליה נפנה בהמשך, ניתנת לגרעין חשיבות מיוחדת, אזי, ביציאה מההצגה הכרונולוגית, נשקול כאן את ההיסטוריה של גילוי החלק החשוב ביותר של התא. הגרעין הוא שעזר לשוואן להשוות בין תאי בעלי חיים וצמחים, ולכן גילוי הגרעין מסמן את השלב החשוב ביותר בהתפתחות התיאוריה של התא.
הגרעינים נראו לראשונה באריתרוציטים של דגים על ידי Löwenhoek בשנת 1700 ותארו אותם באיור. בהמשך אותו עצם - אריתרוציטים של בעלי חוליות וחסרי חוליות רבים - שרטט יוסון (1777) גרעינים. המשמעות של החינוך הזה בכך מחזור מוקדםמקור המיקרוסקופיה, כמובן, לא יכול היה להיות מוערך לא על ידי המחברים עצמם או על ידי בני דורם. פונטנה במחקרו על ארס הצפעים, המתאר את תאי האפיתל של האפידרמיס והאריתרוציטים, מצייר את תאי הגרעין ומזכיר בקצרה "אותם בטקסט; אלא גם באותה תקופה (עבודתו של פונטן פורסמה ב-1781), כאשר ה בדיקה מיקרוסקופיתרקמות של בעלי חיים, לא ניתן היה להבין את התגלית של פונטן.
במקביל, כמה חוקרים צפו בגרעינים בביצים. קאבוליני (פיליפו קאבוליני, 1756-1810) ראה גרעינים בביצי דגים (1787); ופולי (פולי, 1791) ציינו את הגרעינים בביצים של רכיכות. התצפיות שלהם עברו ללא עקבות, מבלי למשוך תשומת לב לעצמן.
במחקר על ביצת הציפורים (1825), תיאר Purkinė את "שלפוחית העובר" (vesicula germinativa). זה היה גרעין של ביצת ציפור. לפי התיאור של פורקין, זוהי "בועה כדורית דחוסה, לבושה במעטפת הדקה ביותר. היא מכילה לימפה משלה, משולבת בפקעת הינקית הלבנה, והיא מלאה בכוח יצירתי, ולכן קראתי לה "השלפוחית העוברית". פורקין התחבר לחינוך שגילה חשיבות רבה; אחריו, החוקרים הבאים כבר לא התעלמו מה"בועה" המסתורית הזו. גילוי פורקינט, אם כן, לא עבר ללא עקבות, כמו תצפיותיהם של קבוליני ופולי, אך המשמעות של "שלפוחית הנבט" נותרה לא ברורה במשך זמן רב, שכן בהבנת חלקי הביצה, מהנקודה מבחינת המושג "תא, הדרך הנכונההוצג רק לאחר המחקר של שוון.
בצמחים, התמונה הראשונה של גרעין התא נוצרה על ידי באואר ב-1802, אך ציור זה פורסם רק ב-1830 (J. Baker, 1949). Meyen (1830) מראה גרעין באיור אחד. במחקר על המרצ'נטיה, מירבל (1831-1832) מתאר גם גרעין, נותן לו שם של כדור; גם הבוטנאי הצרפתי ברוגנר (Adolphe Brogniart, 1801-1876) ראה אותו. אבל התצפיות הראשונות הללו על גרעינים בתאי צמחים לא זכו להערכה על ידי הצופים עצמם וגם לא משכו תשומת לב.
ההכרה בגרעין כחלק הכרחי מתא הצמח היא הכשרון של הבוטנאי האנגלי רוברט בראון (רוברט בראון, 1773-1858).
החל את עבודתו הבוטנית בתיאור האוספים שנעשו במהלך טיול לאוסטרליה, בראון עובר למחקר מבנה אנטומיצמחים. הוא לא קבע בעיות מורפולוגיות גרידא בעבודתו; מחקרים אנטומיים עבורו הם כלי ללימוד טקסונומיה של צמחים, אך בעבודות אלה מגלה בראון תגליות בוטניות יוצאות דופן לגבי רבייה בצמחים. ב-1833 התפרסמה עבודתו של בראון "על האיברים ושיטת ההפריה בסחלבים" (דווחה לאגודת Linnean בלונדון כבר בנובמבר 1831). בראון כותב במאמר זה שבכל תא של האפידרמיס הוא ראה "ערולה מעוגלת אחת, בדרך כלל כהה יותר מקרום התא. עטרה זו היא פחות או יותר גרגירית, מעט קמורה, ולמרות שנראה שהיא שוכבת על פני השטח, היא למעשה מכוסה על ידי הצלחת החיצונית של התא. מיקומו בתא אינו קבוע; אולם לעתים קרובות במרכז או בקרבתו" (עמ' 710). עטרה זו, או גרעין (גרעין) התא, כפי שבראון מתייחס אחרת להיווצרות זו, נצפה על ידו לא רק בתאי האפידרמיס; הוא ראה את הגרעין בפרנכימה, בתאים הפנימיים של חלקי הצמח, "במיוחד כשהם נקיים מחומר גרגירי". בראון, עם זאת בזהירות, מציע שהגרעין הוא רגיל חלק בלתי נפרדתאים. אין לו אמירה קטגורית שהגרעין הוא אברון חובה של התא; כמו כן, בראון אינו נותן תמונות של גרעיני תאים בעבודתו. למרות זאת, במחקריו של בראון, לראשונה, הגרעין אינו מוזכר כיצור אקראי בתא, אלא מופיע כחלק מהותי כלשהו שחשוב לחיי התא.
מיין, מחברת "פיטוטומיה" - עבודה שנידונה קודם לכן - במדריך מאוחר יותר " מערכת חדשההפיזיולוגיה של הצמחים" (1837-1839) מזכירה את הגרעין כחלק קבוע מהתא, שמשמעותו נותרה מסתורית. למעשה, רק עבודתו של Negeli (S. Nageli, 1844) הוכיחה את ההפצה האוניברסלית של גרעיני תאים לא רק בצמחים פורחים, אלא גם בתאים של אצות, פטריות, טחבים ואחרים. צמחים נמוכים יותר.
בהיסטולוגיה של בעלי חיים, המונח "גרעין" הוצג על ידי ולנטין. בהודעה "אוי מבנה טובאיברי חישה" (1836) ולנטין כתב על האפיתל של הלחמית: "הוא מורכב מתאי מעוינים או מרובעים השוכנים צמוד זה לזה, שגבולותיהם נוצרים על ידי קווים חוטיים פשוטים; בכל תא, ללא יוצא מן הכלל, יש גרעין (גרעין) כהה יותר וקומפקטי יותר בעל צורה עגולה או מלבנית. לרובהיא תופסת את אמצע התא, מורכבת מחומר עדין, אך מכילה בתוכו גוף עגול לחלוטין, שלפי אותה תבנית יוצר בתוכו מעין גרעין שני" (עמ' 143). תיאור זה מראה כי ולנטין צפה בבירור בגרעינים של תאי אפיתל. בתוך הגרעין, ולנטיין ראה את הגרעין; זה היה כנראה התיאור הראשון של המבנה התוך-גרעיני הזה.
תיאור ברור ותמונה של גרעינים בתאי אפיתל ניתן על ידי Henle (1837). בייקר (1949) מציין נכון שעבודתם של ולנטין והנל מתחילה את עידן התאים הגרעיניים בהיסטולוגיה של בעלי חיים.
ב-1838 הופיע בארכיון מולר מאמר מאת הבוטנאי הצעיר שליידן תחת הכותרת "חומרים לפיטוגנזה" (Beitrage zur Phytogenesis). עבודה זו נחשבת באופן מסורתי אבן דרךבפיתוח ההוראה הסלולרית, ומחברה מוכר, יחד עם שוואן, כיוצר תורת התא. אין עוררין על משמעותה של שליידן בתולדות התיאוריה התאית, אך בספרות חינוכית, פופולרית ולעיתים גם בספרות ההיסטורית, משמעות זו מכוסה באופן שטחי ושגוי. לפעמים מיוחסת שליידן ככמעט גילתה תאי צמחים, בגלל זה. צריך להבין מהי באמת המשמעות של המדען הזה בהיסטוריה של ההוראה הסלולרית, איפה האמת באגדה שהתפתחה סביב עבודתו ועוברת באופן מסורתי מספר לימוד לספר לימוד.
מתיאס שליידן(מתיאס יעקב שלידן, 1804-1881) הוא הנציג הגדול ביותרבוטניקה גרמנית של אמצע המאה הקודמת. בתחילה סיים את לימודיו בפקולטה למשפטים ועסק בעריכת דין. ללא הצלחה בפעילות זו, שליידן ב-1831 ויתר על משפטים והחל ללמוד רפואה ו מדעי הטבע. משנת 1840 היה פרופסור לבוטניקה בינה, שם שהה עד 1862. זוהי התקופה העיקרית פעילות יצירתיתשליידן. בשנת 1842 התפרסמה עבודתו המרכזית "יסודות הבוטניקה המדעית", אשר מילאה תפקיד חשוב בכיוון של מחקר בוטני נוסף. במקום חשיבה טבעית-פילוסופית, שליידן דורש מבוא לבוטניקה שיטות מדויקותמחקרים על מבנה ותפקוד של צמחים; הוא הדגיש במיוחד את הצורך בתשומת לב להיסטוריה של הפיתוח, שבה ראה את המפתח לפתרון בעיות רבות שנויות במחלוקת. עמדות פילוסופיות של שליידן, המוצהרות על ידו בכתביו, אינן מקוריות וחושפות את חותמה של הפילוסופיה הקנטיאנית. משנת 1862 עד 1864 היה שליידן פרופסור לאנתרופולוגיה בדורפט (כיום טרטו, SSR אסטונית), ב-1864 עזב את דורפט עקב התנגשות עם חוגי הכנסייה ובמקביל הפסיק פעילות פדגוגית. להיות המחבר של לא רק סדרה עבודות מדעיות, אבל גם יצירות פופולריות רבות, שליידן היה ידוע ברבים.
"חומרים לפיטוגנזה" היא עבודתו השנייה של שלידן, שהיה אז עדיין בוטנאי מתחיל. מדובר במאמר, בגודל של כ-40 עמודים, אליו מצורפות שתי טבלאות. חוק היסוד הכללי של המוח האנושי, - כך מתחיל שליידן את עבודתו, - החוק הקובע את רצונו הבלתי ניתן לעמוד בפניו לאחדות בהכרה וביסוס, הן באופן כללי במדע והן בתחום האורגניזמים, של אנלוגיה לשניהם. מחלקות גדולות - ממלכת החיות והצמחים, - הניעו לעסוק שוב ושוב בנושא זה. כל כך הרבה מוחות היו עסוקים בזה, אבל - אי אפשר להכחיש זאת - כל הניסיונות שנעשו עד כה בעניין זה כשלו והיו אשליות. הסיבה נעוצה בעובדה שלמושג הפרט, במובן זה שהוא מיושם בטבע החי, אין יישום בעולם הצומח. הדבר הגדול ביותר שניתן לומר על הפרט במובן זה הוא בצמחים הנמוכים ביותר, כמה אצות ופטריות, המורכבים מתא אחד בלבד. אבל כל צמח מפותח יותר הוא מצרף של אינדיבידואליות סגורות אינדיבידואליות לחלוטין... שהם תאים" (עמ' 137). הבאנו את זה בכוונה ציטוט ארוך, שהוא תחילת מאמרו של שליידן, כדי להראות עד כמה זר לו הרעיון של אחדות המבנה המיקרוסקופי של בעלי חיים וצמחים, שבאה לידי ביטוי במבנה התא. בינתיים, הרעיון הזה הוא אבן היסוד של התיאוריה הסלולרית, שאחד ממחבריה נחשב בדרך כלל שליידן.
להערכה נכונה של עבודתו של שליידן, יש לזכור את עמדתה של תורת התא בבוטניקה עד 1837, כאשר השלידן השלים את עבודתו. הרעיון השקרי לחלוטין ששלידן הוכיח את ההפצה האוניברסלית של תאים בצמחים, או אפילו גילה תאים בצמחים. זהו עיוות של המציאות. התפתחות היסטוריתמדעים. בתחילת שנות השלושים של המאה הקודמת, נוצר רעיון שלם של התא כמבנה יסודי בבוטניקה. צוֹמֵחַ; שליידן בעבודתו נוקט בעמדה זו כמסקנה מבוססת ללא עוררין. אפילו חלקים לא תאיים כאלה של צמחים, כפי שנראה קודם לכן, ככלי עץ נושאי מים, נחשבים בשלב זה כתאים מתוקנים, מובחנים באופן מיוחד ומוזגים. שליידן לא היה צריך לבסס את ההפצה האוניברסלית של תאים בצמחים: ביסוס עמדה זו הייתה, כפי שראינו, הצלחה קולקטיבית של מספר יצירות של גלקסיה רבים של בוטנאים ברבע הראשון של המאה הקודמת.
ק.א.טימיריאזב (1920) כתב בצדק על הביטוי "גילוי התא": "אבל העובדה היא שאיש לא גילה את התא" (עמ' 79), ובכך הדגיש ש"גילוי" התא אינו הכשרון של התא. איזה מדען מסוים. זה גם לא נכון שליידן, כפי שכותב אשוף (1938), פיתח את הדוקטרינה של "הבנייה החובקת מתאי כל הצמחים" (עמ' 177). ובעניין זה צודק ק"א טימירייזב שכתב: "שליידן נחשב בדרך כלל ליוצר תורת התא הזה, שהתגלה כל כך עשיר בהכללות הפוריות ביותר. אבל זה בקושי הוגן... שליידן, יריב רהוט ונלהב לשגרה ולסטגנציה, יכול היה לומר על עצמו בצדק, כפי שעשה בייקון פעם, שהוא חצוצרן, מבשר, מבשר, המכריז על הופעתה של דוקטרינה זו, אבל נתונים עובדתיים המבססים זאת כבר היו קיימים קודם לכן..." (עמ' 75). אופייני לכך שאונגר (Unger, 1846) ביסודות הבוטניקה שלו, המתווה את מיקומו של התא כמבנה היסודי האוניברסלי של אורגניזמים, מצביע על Schwann ו-Kölliker בהתייחסות הספרותית, מבלי להזכיר אפילו את שליידן בהיבט זה.
עצם הרעיון של התא בשלידן אינו שונה מהרעיונות שהתגבשו קודם לכן ובאו לידי ביטוי בספר הלימוד של מייאן (1830) עוד לפני שליידן החל ללמוד בוטניקה. ברמה של רעיונות אלה, שליידן לקח את התא עבור בועה או תא, תחום על ידי קליפה, שבתוכה ניתן לאתר את התוכן. ה"תוכן" הזה של התא (פרוטופלזמה עתידית!) משך גם את תשומת לבה של מיין, שהקדישה לו מחקר רב, אך לא הבינה את משמעותו של מרכיב תאים עיקרי זה. שליידן ראה גם את הפרוטופלזמה של תאי הצמח, אבל הוא לא הבין את המשמעות של "התוכן" של התא. מבחינתו זה מסטיק (גומי) או ג'לי (גלרטה). חלק מהפרוטופלזמה שליידן מיוחסת לדופן התא. האחרון, לדעתו, מורכב משתי שכבות, ביניהן נמצא גרעין התא - "דיטובלסט", שלעולם אינו שוכן בתוך התא, אלא תמיד מוקף בדופן התא "באופן שדופן התא מתפצל לשניים. לוחות, שאחד מהם עובר החוצה והשני בתוך הציטובלסט. זה שהולך עם בְּתוֹך, בדרך כלל רך יותר וג'לטיני יותר" (עמ' 142). מהציורים של שלידן ברור איזה סוג של "רובד פנימי". דופן תא» הוא לקח את השכבה הקדמית של הפרוטופלזמה של תאי צמחים.
איזו משימה הטיל שליידן בעבודתו? "כל תא", הוא כותב, "מנהל חיים כפולים: חיים עצמאיים לחלוטין, הקשורים רק להתפתחותו שלו, ועוד אחד תלוי, שכן הוא חלק בלתי נפרד מהצמח. עם זאת, קל לראות שגם לפיזיולוגיה של הצמח וגם לפיזיולוגיה השוואתית, התהליכים החיוניים של תאים בודדים צריכים בדרך כלל להיות במקום הראשון, צריכים להוות בסיס בלתי נמנע, ובמקרה זה, השאלה מובאת קודם כל. : איך זה מוזר אורגניזם קטן, תא?" (עמ' 138). משימה זו, יצירת התא, היא הבסיס למאמר של שלידן. הרגע הגנטי במובן זה הועלה לפני כן, אך לא ניתן להכחיש כי שליידן, בהתאם לזמנו, הציג בעיה זו בצורה ברורה יותר מקודמיו.
בתשובה לשאלה שהועלתה, שליידן מפתח את התיאוריה שלו על היווצרות תאים. בתיאוריה זו, התפקיד המרכזי בפיתוח תאים חדשים מוקצה לגרעין. כפי שראינו, הוא התגלה הרבה לפני עבודתו של שליידן, אך לא זכה לשום פרשנות ודאית. לדברי שלידן, הגרעין הוא "ציטובבלסט" - הראשון של התא. ניתן לאפיין בקצרה את התיאוריה של היווצרות תאים שפותחה על ידי שלידן כדלקמן.
במסטיק, צמוד מבפנים לדפנות של תאים קיימים, נוצרים גרגרים; שליידן קורא להם ריר ומאמין שגרגרים אלו, על ידי עיבוי, יוצרים נוקלאולים, ואז נוצר גרעין, המופיע כמשקע גרגירי סביב הגרעין. על פני הגרעין, מצד אחד, נוצרת שוב קליפה מה"ליחה"; הוא תוחם את הציטובלסט, וכך נוצר חלל חומה, שבו הגרעין מוקף בעובי הקיר. החלל הזה הוא התא החדש. לכן, לפי שלידן, תאי בת מתעוררים בתוך תאי האם. מספר התאים החדשים שיכולים להתפתח בתא אימהי אחד, כמו גם גורלו של תא חינוכי אימהי זה, אינו נדון על ידי שלידן.
זוהי המהות של תורת היווצרות התאים, תמצית "המחקר המצוין של שליידן, ששפך כל כך הרבה אור על התחום הזה", - אפיון עבודתו של שליידן שניתן על ידי תיאודור שוואן. כפי שהוכח עד מהרה, התיאוריה של שליידן מבוססת על תצפיות שגויות בפירוש. את התיאוריה השגויה הזו של היווצרות תאים השתלט חברו שוואן משליידן, וזו הייתה הנקודה החלשה ביותר בתורתו של שוואן. סאקס, בתולדות הבוטניקה שלו, מאפיין את התיאוריה של שליידן במילים הקשות הבאות: "תיאוריית היווצרות התאים של שליידן נבעה ממפגש בלתי מובן של תצפיות לא ברורות ודעות קדומות, יתרה מכך, היא דומה מאוד בעיקר לתיאוריות הישנות של ספרנגל וטרוויראנוס" ( עמ' 76). שליידן עצמו הגן בעקשנות על תיאוריית הציטוגנזה שלו וציטט אותה אפילו במהדורה הרביעית. "יסודות הבוטניקה המדעית" (1861).
במאמרו, שליידן, בנוסף לתיאוריה הנחשבת של יצירת תאים, עוסק בהתפתחות של עיבויים על דפנות כלי ספירלה ומפתח טיעונים תיאורטיים על עבודתם של צמחים. עבודתו של שליידן אינה מכילה שום דבר חדש מיסודו בחלק זה, ומכיוון שחלק זה של המאמר אינו קשור ישירות לנושא שלנו, אין צורך להתעכב עליו.
איזה דירוג צריך לתת היבט היסטוריתפקידו של שליידן בפיתוח מדע הסלולר? מרטין היידנהיין (M. Heidenhain, 1899) בסוף המאה הקודמת ציין את חוסר הנכונות של הרעיון של ערך שווה ערך בהיסטוריה של התיאוריה התאית של שלידן ושוואן. מאוחר יותר, שאלה זו הועלתה שוב בכל החלטיות על בסיס ניתוח ביקורתי של הספרות של ההיסטולוג הצ'כי סטודניצ'קה (1933), אנין גדול של תולדות תורת התא. ואכן, ההשוואה המסורתית של שמותיהם של שלידן ושוואן, המובאים בדרך כלל כ"מחברים שותפים" של תורת התא, אינה מוצדקת על ידי מחקר מדוקדק של המקורות. שליידן לא היה שותף לתיאוריית התא; הוא, כפי שראינו, היה זר לחלוטין לרעיון המרכזי של תיאוריה זו - אחדות המבנה היסודי המיקרוסקופי של בעלי חיים וצמחים; הוא אינו היוצר של הדוקטרינה התאית בתחום הבוטניקה, שכן ההוראות העיקריות של דוקטרינה זו פותחו לפניו. יש להדגיש זאת, שכן בספרות, גם הזרה וגם שלנו, נוצרה "אגדה" סביב שמו של שליידן, שיש רבים כל כך בתולדות המדע עקב היכרות לא מספקת עם המקורות. סטודניצ'קה, במאמר שצוטט לעיל על שליידן, ציטט תמציות מכמה עשרות מדריכים זרים על היסטולוגיה וביולוגיה, ואפילו ממאמרים מיוחדים על ההיסטוריה של מדע הסלולר, שבהם תפקידו של שליידן מוצג באופן שגוי לחלוטין והאגדה חוזרת על עצמה כי המדע. חייב שליידן את התגלית מבנה תאיבצמחים, ששליידן ושוואן יצרו את תורת התא וכו'. לרשימת ההצהרות הלא מוצדקות, ולפעמים פשוט מגוחכות על תפקידו של שליידן, שסטדניך ציטט, אפשר לצערי להוסיף רשימה לא מבוטלת של ציטוטים מספרי לימוד חדשים יותר. אפילו יצירות מיוחדות, כולל כתבים המתיימרים להיות יצירות היסטוריות גם אצלנו וגם אצלנו ספרות זרה. התפקיד ההיסטורי של יצירתו של שלידן אינו מוטל בספק, אך תפקיד זה שונה ממה שמכוסה בדרך כלל. שליידן זוכה להכנסת הגישה הגנטית לתיאוריית הרקמות והתאים. ניסיונות של גישה דומה נעשו לפני שליידן (וולף, מירבל, ספרנגל, טרוויראנוס; בהיסטולוגיה של בעלי חיים - ולנטין), אך באותה תקופה הם לא יכלו להיות יעילים כמו עבודתו של שליידן, שהופיעה כאשר הרעיון של התא כמו המבנה הבסיסי של הצמחים כבר היה נפוץ. ללא הגישה הגנטית, שואן לא היה יכול ליצור תיאוריה סלולרית קוהרנטית, המבוססת על ידי נתונים משכנעים לאותה תקופה. רק על ידי התייחסות להיסטוריה של התפתחות רקמות ותאים, שוון הצליח להראות את ה"התכתבות" של מבנים יסודיים שונים, יכול להוכיח את ההומולוגיה שלהם. עבודתו של שליידן, כמובן, מילאה תפקיד משמעותי בהכוונת הגותו של שוון לנתיב מחקר דומה.
אבל זה לא הכל. על מנת להיות מסוגלים להראות באופן משכנע את ההומולוגיה שלהם על ידי התייחסות להיסטוריה של התפתחות מבנים יסודיים, היה צורך למצוא תכונה מנחה, ובהתחשב בחוליה מובילה, לפרום את סבך היחסים המורכבים בין מבנים יסודיים בבעלי חיים רקמות. שוואן למד את התכונה המנחה הזו משליידן. זו הליבה. תאים פנימה בדים שוניםעשויים להיות שונים מאוד זה מזה כלפי חוץ, אבל הדמיון של הגרעינים בולט, ועוזר להומולוגיה של תצורות שונות כלפי חוץ. הגרעין היה ידוע הן בתאי צמחים והן במבנים של בעלי חיים לפני שלידן. אבל רק בעבודתו רכש הגרעין את המשמעות של התכונה העיקרית של התא המתפתח. השלט הזה שימש מנוף עבור שוואן, ותפס אותו הוא הצליח ליצור תיאוריית תאים.
זו המשמעות של שליידן בהיסטוריה של מדע הסלולר. אי אפשר להציב אותו ליד שוון, הוא לא היה מחבר שותף לתיאוריית הסלולר, אבל עבודתו הייתה חוליה הכרחית בשרשרת המחקר שהכינה את החומר, שבלעדיה אולי הגאונות של שוון הייתה חסרת אונים. לעשות את ההכללות שהוא ניסח בצורה של תיאוריה תאית. Virchow (1859) ביטא זאת נכון בכך שציין ששוואן עמד "על כתפיו" של שליידן.
אם אתה מוצא שגיאה, אנא סמן קטע טקסט ולחץ Ctrl+Enter.
תרשים של גרעין תא B ביולוגיה של התא, הליבה(lat. nucleus) - אברוני תאים המצויים ברוב התאים האוקריוטים ומכילים גנים גרעיניים המרכיבים רובחומר גנטי. לגרעין שני תפקידים עיקריים: לכוון תגובות כימיות בתוך הציטופלזמה ואחסון המידע הדרוש לחלוקת התא.
הגרעין התגלה בסביבות 1833 על ידי הבוטנאי רוברט בראון.
בנוסף לגנום התא, הגרעין מכיל חלבונים מסוימים המווסתים את קריאת המידע הגנטי. קריאת גנים ברמה הגרעינית מושכת תהליכים מורכביםשעתוק, עיבוד mRNA ראשוני ויצוא של mRNA בוגר לציטופלזמה.
קוטר הליבה בדרך כלל 8-25 מיקרומטר. הוא מוקף בממברנה כפולה הנקראת המעטפת הגרעינית. נקבוביות גרעיניות עוברות דרך הממברנות הפנימיות והחיצוניות במרווחים מסוימים. המעטפת הגרעינית מווסתת ומקלה על התחבורה בין הגרעין לציטופלזמה על ידי הפרדה תגובה כימיתהמתרחש בציטופלזמה מתגובות המתרחשות בתוך הגרעין. הממברנה החיצונית רציפה עם הרטיקולום האנדופלזמי המחוספס (RER) וייתכן שיש לו ריבוזומים קשורים. המרווח בין שני הממברנות (שנקרא "החלל הפר-גרעיני") הוא רציף עם לומן RER. צד גרעיני מעטפת גרעיןמוקף ברשת של חוטים הנקראים הלמין הגרעיני.
חלק פנימיהגרעין מכיל נוקלאולי אחד או יותר המוקפים במטריצה הנקראת נוקלאופלזמה. נוקלאופלזמה היא נוזל דמוי ג'ל (דומה מבחינה זו לציטופלזמה) שבו מומסים חומרים רבים. חומרים אלו כוללים נוקלאוטיד טריפוספט, מולקולות איתות, DNA, RNA וחלבונים (אנזימים וחוטים).
גרעין ומערכת אנדוממברנה חומר גנטי קיים בגרעין בצורה של כרומטין, או קומפלקס של חלבון ו-DNA. DNA קיים כמו שורה שלמהמולקולות נפרדות הידועות ככרומוזומים. ישנם שני סוגים של כרומטין: אאוכרומטין והטרוכרומטין. Euchromatin הוא צורה פחות קומפקטית של DNA, אזורי ה-DNA בצורת Euchromatin מכילים גנים הנקראים לעתים קרובות על ידי התא.
בהטרוכרומטין, ה-DNA ארוז בצורה קומפקטית יותר. אזורי DNA בצורה של הטרוכרומטין מכילים גנים שאינם נקראים על ידי התא בשלב זה של ההתפתחות (סוג זה של הטרוכרומטין ידוע כהטרוכרומטין פקולטטיבי) או שהם אזורים המרכיבים את הטלומרים והצנטרומרים של הכרומוזומים (סוג זה של הטרוכרומטין ידוע בשם הטרוכרומטין בונה). בְּ אורגניזמים רב תאיים, תאים מתמחים ביותר לבצע פונקציות ספציפיות, ולכן יש צורך בסטים שונים של גנים ולקרוא אותם. לכן, אזורי ה-DNA שהם בצורה של הטרוכרומטין תלויים בסוג התא.
הגרעין הוא מבנה צפוף בגרעין שבו מורכבים יסודות הריבוזום. לגרעין עשוי להיות נוקלאולי אחד או יותר.
III. גרעין התא.
הגרעין התגלה ותואר ב-1831 על ידי הבוטנאי האנגלי ר' בראון.
הוא אופייני לכל התאים האוקריוטיים, למעט אריתרוציטים בוגרים של יונקים וצינורות מסננת צמחים. תאים יכולים להיות חד-גרעיניים (בדרך כלל); מרובי גרעינים (תאי כבד ושריר בבני אדם ויונקים, פטריות וכו'). הצורה והגודל של גרעין התא משתנים מאוד ותלויים בסוג האורגניזם, כמו גם בסוג, גיל ו מצב תפקודיתאים. בתאים עגולים או מצולעים, הוא בדרך כלל כדורי, בתאים מוארכים הוא בצורת מוט או סגלגל, בלוקוציטים הוא בצורת כפה או אפילו רב אונות (בתאי בלוטות הארכנואיד של כמה חרקים ועכבישים). קוטר הגרעין מ-3 עד 10 מיקרון, (5 - 20 מיקרון)
על ידי תרכובת כימיתהגרעין נבדל משאר התא תוכן גבוה DNA (15 - 30%) ו-RNA (12%). 99% מה-DNA של התא מרוכז בגרעין, שם הוא, יחד עם חלבונים (היסטונים ולא היסטונים), יוצר קומפלקסים - דאוקסיריבונוקלאופרוטאין (DNP).
פונקציות ליבה:
1) אחסון ושכפול מידע גנטי;
2) ויסות תהליכים מטבוליים המתרחשים בתא.
מחזור החיים של תא (מחזור התא) כולל שתי תקופות:
1) חלוקה, כתוצאה ממנה נוצרים שני תאי בת;
2) התקופה בין שתי חלוקות, הנקראת - interphase.
במהלך האינטרפאזה, התא מתכונן לחלוקה: התא גדל, שכפול DNA מתרחש, ו חומרים שוניםהכרחי לביקוע, אנרגיה מאוחסנת. בתהליך של חלוקה מבני תאיםהליבות עוברות שינויים משמעותיים.
בגרעין הבין-פאזי, ישנם: קרום גרעיני; מיץ גרעיני; נוקלאולי, וכרומטין.
מעטפת גרעין(cardiolemma) מיוצג על ידי שני ממברנות יסוד, שביניהם ממוקם חלל פרי גרעיני . הממברנה החיצונית של הממברנה הגרעינית, הגובלת בהיאלופלזמה, היא בעלת מבנה מקופל מורכב, במקומות מסוימים מחובר לתעלות EPS. החלל הפרי-גרעיני וצורת ER מערכת בודדתערוצי תקשורת. ריבוזומים ממוקמים על הממברנה הגרעינית החיצונית, הממברנה הפנימית חלקה. הממברנה הגרעינית מחלחלת בנקבוביות רבות, הקוטר הוא כ-30 - 100 ננומטר. מספרם משתנה בהתאם לפעילות התפקודית של הגרעין. תמיד יש יותר נקבוביות בתאים צעירים מאשר בתאים ישנים.
פונקציית פגז גרעיני:
1. ויסות חילוף החומרים בין הגרעין לציטופלזמה;
2. תפקוד מגן.
חילופי החומרים בין הגרעין לציטופלזמה מתבצעים במספר דרכים:
1. דרך נקבוביות מתן חדירות סלקטיבית;
דוגמה: שחרור של mRNA ותת-חלקיקים ריבוזמליים לציטופלזמה, או כניסה לגרעין של חלבונים, נוקלאוטידים ומולקולות ריבוזומליות המווסתות את פעילות ה-DNA.
2. על ידי שרוך החוצה את הצמחים והבליטות של המעטפת הגרעינית.
3. על ידי דיפוזיה דרך המעטפת הגרעינית.
למרות חילופי החומרים הפעילים בין הגרעין לציטופלזמה, הממברנה הגרעינית תוחמת את התוכן הגרעיני מהציטופלזמה, ומאפשרת את קיומה של סביבה תוך גרעינית מיוחדת השונה מהציטופלזמה שמסביב.
מיץ גרעיני(נוקלאופלזמה; קריופלזמה) היא מסה הומוגנית הממלאת את החלל בין מבני הגרעין (כרומטין ונוקלאולי). היא מורכבת מחלבונים (אנזימים), נוקלאוטידים, חומצות אמינו ו סוגים שונים RNA (mRNA, tRNA, rRNA) ו-DNA, כמו גם תוצרי ביניים מטבוליים. מיץ גרעיני מבצע את מערכת היחסים של מבנים גרעיניים וחילופין עם הציטופלזמה של התא.
כרומטיןמייצג deoxyribonucleoprotein(DNP), זוהה תחת מיקרוסקופ אור בצורה של גדילים דקים (סיביים) של גרגירים קטנים או גושים. הבסיס של הכרומטין הוא נוקלאופרוטאין - מולקולות DNA חוטיות ארוכות (כ-40%), המחוברות לספציפיות חלבונים - היסטונים(40%) מכיל מספר גדול שלליזין וארגינין.
בתהליך של ביקוע גרעיני, נוקלאופרוטאין מסתחררים, מתקצרים, ובכך נדחסים (פי 100-500) לכרומוזומים דחוסים בצורת מוט.
כרומוזומים(כרום יווני - צבע וסומה - גוף) נקראו כך בקשר ליכולת הכתמה אינטנסיבית. כרומוזומים מכילים גם RNA, חלבונים חומציים, שומנים ו מינרלים(יוני Ca ו-Mg), כמו גם האנזים DNA פולימראז, הדרוש לשכפול DNA. במהלך interphase, כרומוזומים נראים רק מתחת מיקרוסקופ אלקטרוניוהוא DNP הנקרא כרומטין (מצב דה-ספירלי של כרומוזומים).
כרומוזום מטאפאזהמורכב משני גדילים אורכיים של DNP - כרומטידים, המחוברים זה לזה באזור ההיצרות הראשונית - צנטרומרים . בחלק המרכזי של הצנטרומר נמצאים kinetochoresאליהם מתחברים מיקרוטובולים של חוטי ציר הביקוע במהלך מיטוזה. לחלק מהכרומוזומים יש היצרות משנית, לא קשור לחיבור של חוט הציר. אזור זה של כרומוזומים שולט בסינתזה של הגרעין (מארגן גרעיני) והוא נקרא לוויין. הצנטרומר מחלק את הכרומוזום לשתי זרועות.
מיקומו של הצנטרומר קובע את 3 סוגי הכרומוזומים העיקריים:
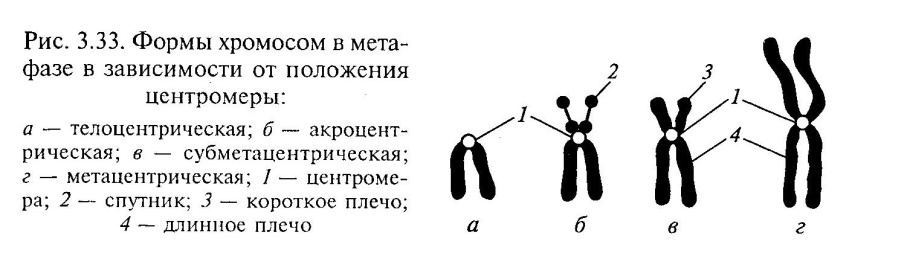 שווה-כתף
(מטאצנטרית) - עם כתפיים באורך שווה;
שווה-כתף
(מטאצנטרית) - עם כתפיים באורך שווה;
תת-מטצנטריאו כתפיים לא אחידות - כתפיים באורך לא שווה;
בצורת מוט- עם כתף אחת ארוכה והשנייה קצרה מאוד (אקרוצנטרית).
מבנה הכרומוזומים נראה בבירור בשלב המטאפאזה של המיטוזה. חקר הכרומוזומים איפשר לקבוע את העובדות הבאות:
ü בכל התאים הסומטיים של כל אורגניזם צמחי או בעל חיים, מספר הכרומוזומים זהה;
ü תאי מין מכילים תמיד חצי כרומוזומים מאשר בתאים סומטיים של סוג נתון של אורגניזם;
ü לכל האורגניזמים השייכים לאותו מין יש אותו מספר של כרומוזומים בתאים.
דוגמה: תולעת סוס עגולה - 2; זבוב תסיסנית - 8; שימפנזים - 48; אפר - 46; גבר - 46; יונה - 80; קרפיון - 104; רדיולריה (פרוטיסטית) - 1000 - 1600.
מספר הכרומוזומים אינו תלוי בגובה הארגון ולא תמיד מעיד על קשר פילוגנטי: אותו מספר ניתן למצוא במינים המרוחקים מאוד זה מזה באופן שיטתי ונבדלים מאוד באורגניזמים קרובים במקורם. לכן מספר הכרומוזומים אינו תכונה ספציפית למין. עם זאת, המאפיין של מערך הכרומוזומים בכללותו הוא ספציפי למין, כלומר. מאפיין רק סוג אחד של צמח או בעל חיים.
קריוטיפ- קבוצה של תכונות כמותיות (מספר וגודל) ואיכותיות (צורה) של קבוצת הכרומוזומים של תא סומטי. מספר הכרומוזומים בקריוטיפ הוא תמיד זוגי. זאת בשל העובדה שבתאים סומטיים יש 2 כרומוזומים באותו צורה וגודל. האחד מגיע מהאורגניזם האבהי, השני מהאימהי.
כרומוזומים באותו גודל וצורה ונושאים את אותם גנים נקראים הומולוגי.
מספר הכרומוזומים בתאי נבט בוגרים נקרא הפלואיד (יחיד) ומסומן אות לטינית נ.תאים סומטיים מכילים מספר כפול של כרומוזומים - סט דיפלואידי - מסומן 2n.תאים עם יותר משתי קבוצות של כרומוזומים נקראים פוליפלואיד (4n, 8n, 16n וכו')
נוקלאולי -אלה מעוגלים, דחוסים מאוד, אינם מוגבלים על ידי אזורי הממברנה של גרעין התא בקוטר של 1-2 מיקרומטר או יותר. הצורה, הגודל והמספר של הגרעין תלויים במצב התפקודי של הגרעין: ככל שהגרעין גדול יותר, כך פעילותו גבוהה יותר. הגרעין עשוי להכיל בין 1 ל-10 נוקלאולים, ולפעמים, למשל, בגרעיני שמרים, הם נעדרים לחלוטין. הגרעין מכיל כ-80% חלבון, 10-15% RNA, חלק מה-DNA ורכיבים כימיים אחרים.
יש גרעינים רק בגרעינים שאינם מתחלקים, במהלך החלוקה הם נעלמים, ולאחר השלמת החלוקה הם נוצרים שוב סביב חלקים מסוימים של כרומוזומים - גנים הנקראים מארגנים גרעיניים.
בגרעין, RNA מתחבר עם חלבונים, וכתוצאה מכך נוצרים ריבונוקלאופרוטאין - מבשרי הריבוזומים. האחרונים עוברים דרך הנקבוביות של הממברנה הגרעינית לתוך הציטופלזמה, שם מסתיימת היווצרותם. לפיכך, הגרעין הוא הצטברות של rRNA וריבוזומים על שלבים שוניםתצורות.
הגרעין התגלה ותואר ב-1833 על ידי האנגלי ר' בראון. הגרעין קיים בכל תאים איקריוטיים, למעט אריתרוציטים בוגרים וצינורות מסננת של צמחים. לתאים יש בדרך כלל גרעין בודד, אך לפעמים מוצאים תאים מרובי גרעינים.
הגרעין הוא כדורי או צורה אליפסה. לחלק מהתאים יש גרעינים מפולחים. גודל הגרעינים הוא בקוטר של 3 עד 10 מיקרון.
הגרעין חיוני לחיי התא. זה מווסת את פעילות התא. הגרעין אוגר מידע תורשתי הכלול ב-DNA. מידע זה, הודות לגרעין, מועבר לתאי בת במהלך חלוקת התא. הגרעין קובע את הספציפיות של חלבונים המסונתזים בתא. הגרעין מכיל חלבונים רבים הנחוצים לתפקודיו. RNA מסונתז בגרעין.
לגרעין יש קרום גרעיני המפריד בינו לבין הציטופלזמה, קריופלזמה (מיץ גרעיני), נוקלאולי אחד או יותר, כרומטין
המעטפת הגרעינית מורכבת משני ממברנות. יש לו נקבוביות שמשחקות תפקיד חשובבהובלת חומרים לתוך הציטופלזמה וממנה. נקבוביות אינן תצורות קבועות. מספרם משתנה בהתאם לפעילות התפקודית של הגרעין. מספר הנקבוביות גדל במהלך תקופת הפעילות הגרעינית הגדולה ביותר. המעטפת הגרעינית מחוברת ישירות לרשת האנדופלזמית.
עַל קרום חיצונימעטפת גרעינית, עם בחוץישנם ריבוזומים המסנתזים חלבונים ספציפיים שנוצרים רק על הריבוזומים של המעטפת הגרעינית.
מיץ גרעיני (קריופלזמה) - התוכן הפנימי של הגרעין, הוא תמיסה של חלבונים, נוקלאוטידים, יונים, צמיגיים יותר מהיאלופלזמה. בו
קיימים גם חלבונים פיברילרים. הקריופלזמה מכילה נוקלאולי וכרומטין. מיץ גרעיני מספק תפקוד רגילחומר גנטי.
נוקלאולי הם מרכיב חובה בגרעין, הם נמצאים בגרעיני אינטרפאזי והם גופים קטנים, כדוריים בצורתם. הגרעינים צפופים יותר מהגרעין. בנוקלאולים מתרחשת סינתזה של rRNA, סוגים אחרים של RNA ויצירת תת-יחידות ריבוזום.
הופעת נוקלאולי קשורה לאזורים מסוימים של כרומוזומים הנקראים מארגנים גרעיניים. מספר הנוקלאולים נקבע על פי מספר המארגנים הגרעיניים. הם מכילים גנים של rRNA.
כרומטין (חומר מוכתם) הוא חומר צפוף של הגרעין, מוכתם היטב בצבעים בסיסיים. כרומטין מורכב ממולקולות DNA בקומפלקס עם חלבונים (היסטונים ולא היסטונים), RNA.
בגרעינים שאינם מתחלקים (בין-פאזיים), הכרומטין יכול למלא באופן שווה את נפח הגרעין, בהיותו במצב מעובה. כרומטין מפוזר זה (אאוכרומטין) פעיל גנטית. מולקולות DNA המכילות מידע תורשתי מסוגלות להכפיל את עצמו במהלך השכפול, ואפשרות העברה (שעתוק) של מידע גנטי מ-DNA ל-mRNA.
לפעמים נראים גושים של כרומטין בגרעין הבין-פאזי, שהם אזורים של כרומטין מעובה (הטרוכרומטין). אלו אזורים לא פעילים. למשל, בתאים גוף נשי, כאשר קיימים שני כרומוזומי X, האחד במצב מפוזר פעיל, והשני במצב לא פעיל, מעובה.
במהלך חלוקה גרעינית, הכרומטין צובע בצורה אינטנסיבית יותר, הוא מתעבה - היווצרות חוטים ספירליים (מעוותים) יותר, הנקראים כרומוזומים.
כרומוזומים אינם פעילים מבחינה סינתטית. מבנה הכרומוזומים נחקר בצורה הטובה ביותר ברגע ההתעבות הגדול ביותר שלהם, כלומר במטאפאזה ובתחילת האנפאזה של המיטוזה.
כל כרומוזום במטאפאזה של מיטוזה מורכב משתי כרומטידות הנוצרות כתוצאה משכפול ומחוברים באמצעות צנטרומר (התכווצות ראשונית). בחלק המרכזי של הצנטרומר יש קינטוכורים, אליהם מחוברות מיקרו-צינוריות של חוטי ציר במהלך המיטוזה (איור 47). באנפאזה, הכרומטידות מופרדות זו מזו. נוצרים מהם כרומוזומי בת המכילים מידע גנטי זהה. הצנטרומר מחלק את הכרומוזום לשתי זרועות. כרומוזומים בעלי זרועות שוות נקראים זרועות שוות או מטאצנטריות, עם זרועות בעלות אורך לא שווה - זרועות לא שוות - תת-מטאצנטריות, כאשר אחת קצרה והשנייה כמעט בלתי מורגשת - בצורת מוט או אקרוצנטרי.
לחלק מהכרומוזומים יש התכווצות משנית המפרידה בין הלוויין. התכווצויות משניות נקראות מארגנים גרעיניים. בהם, ב-interphase, מתרחשת היווצרות הגרעין. המארגנים הגרעיניים מכילים DNA
אחראי לסינתזה של rRNA. זרועות הכרומוזומים מסתיימות באזורים הנקראים טלומרים שאינם יכולים להתחבר לכרומוזומים אחרים.
Kinetochores ממוקמים באזור centromere של כרומוזומים. / - kinetochore, 2 - צרור של kinetochore microtubules; 3 - כרומטיד.
מספר, גודל וצורה של כרומוזומים בקבוצה סוגים שוניםעשוי להשתנות. קבוצת התכונות של קבוצת כרומוזומים נקראת קריוטיפ.
מערך הכרומוזומים הוא ספציפי וקבוע עבור פרטים מכל מין. לבני אדם יש 46 כרומוזומים, לעכברים יש 40 כרומוזומים וכן הלאה.
בתאים סומטיים שיש להם קבוצה דיפלואידית של כרומוזומים, הכרומוזומים מזווגים. הם נקראים הומולוגיים. כרומוזום אחד בזוג מגיע ממנו אורגניזם אימהי, השני - מהאב.
שינויים במבנה הכרומוזומים או במספרם נובעים ממוטציות.
כל זוג כרומוזומים בקבוצה הוא אינדיבידואלי. כרומוזומים מזוגות שונים נקראים לא הומולוגיים.
בקריוטיפ מבחינים בכרומוזומי המין (בבני אדם זהו כרומוזום X וכרומוזום Y) ואוטוזומים (כל השאר).
לתאי מין יש סט הפלואידיכרומוזומים.
הבסיס של הכרומוזום הוא מולקולת DNA הקשורה לחלבונים (היסטונים וכו') לתוך נוקלאופרוטאין.
העמדה העיקרית של הביולוגיה המולקולרית, שנוסחה על ידי פ. קריק, קובעת כי העברת המידע הגנטי מתבצעת:
1) מ-DNA ל-DNA על ידי שכפול;
2) מ-DNA דרך mRNA (mRNA) לחלבון.
תהליך הרבייה העצמית של מקרומולקולות של חומצות גרעין (רפליקציה) מבטיח העתקה מדויקת של מידע גנטי והעברתו מדור לדור.
עקרון ההשלמה העומד בבסיס המבנה של מולקולת ה-DNA מאפשר להבין כיצד נוצרות מולקולות חדשות בתקופה הסינתטית של האינטרפאזה מעגל החייםתאים לפני חלוקה.