מיטוכונדריה.בציטופלזמה של תאי בעלי חיים וצמחים, יש מה שנקרא אברוני אנרגיה - מיטוכונדריה (מהיוונית "מיטוס" - חוט, "כונדריון" - דגן). צורת המיטוכונדריה שונה; הם יכולים להיות סגלגלים, בצורת מוט, חוטי עם קוטר ממוצע של 1 מיקרומטר ואורך של 7 מיקרון. מספר המיטוכונדריות תלוי בפעילות התפקודית של התא ויכול להגיע לעשרות אלפים בשרירים המעופפים של חרקים.
מבנה פנימיהמיטוכונדריה (איור 11, 12; 15) נחקרה באמצעות מיקרוסקופ אלקטרונים. מיקרוגרפי אלקטרונים מראים שהמיטוכונדריה מוגבלת חיצונית על ידי קרום חיצוני, שבעצם יש לו מבנה זהה לממברנת הפלזמה. מתחת לקרום החיצוני נמצא הממברנה הפנימית, היוצר קפלים רבים - cristae. בתוך המיטוכונדריה נמצאים RNA, DNA וריבוזומים הנבדלים מאלה ציטופלזמיים. בממברנות שלו מובנים אנזימים ספציפיים, בעזרתם האנרגיה מומרת למיטוכונדריה. חומרים מזיניםלתוך האנרגיה של ATP, הנחוצה לחיי התא והאורגניזם בכללותו.
אורז. 15. סכימה של מבנה המיטוכונדריה
פלסטידים.אלו הם אברונים שייחודיים לתאי צמחים. ישנם שלושה סוגים של פלסטידים: כלורופלסטים ירוקים, כרומופלסטים צבעוניים (אך לא ירוקים) ולקופלסטים חסרי צבע.
כלורופלסט(איור 16) מזכיר בצורתו דיסק או כדור בקוטר 4-6 מיקרון עם ממברנה כפולה - חיצונית ופנימית. בתוך הכלורופלסט יש דנ"א, ריבוזומים ומבני ממברנה מיוחדים - גרגרים המחוברים זה לזה ולמזכר הפנימי ולכלורופלסט, לכל כלורופלסט יש כ-50 גרגרים, המסודרים בתבנית דמקה ללכידת אור טובה יותר. ממברנות גראן מכילות את הפיגמנט הירוק כלורופיל. הודות לכלורופיל בכלורופלסטים, האנרגיה מומרת אוֹר שֶׁמֶשׁלתוך האנרגיה הכימית של ATP. האנרגיה של ATP משמשת בכלורופלסטים לסינתזה תרכובות אורגניותבעיקר פחמימות.
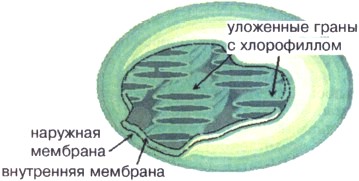
אורז. 16. סכימה של מבנה הכלורופלסט
כרומופלסטים.פיגמנטים של אדום ו צבע צהוב, הממוקם בכרומופלסטים, מעניקים לחלקים שונים של צמחים צבע אדום וצהוב. שורש גזר, פירות עגבניות צבעוניים בגלל פיגמנטים הכלולים בכרומופלסטים. השילוב של כרומופלסטים המכילים פיגמנטים שונים יוצר מגוון רחב של צבעים של פרחים ופירות של צמחים.
Leucoplastsהם מקום הצטברות של חומר תזונתי רזרבה - עמילן. ישנם לויקופלסטים רבים במיוחד בתאי פקעות תפוחי אדמה. באור, לויקופלסטים יכולים להפוך לכלורופלסטים (כתוצאה מכך פקעות תפוחי אדמה הופכות לירוקות). בסתיו, הכלורופלסטים הופכים לכרומופלסטים והעלים והפירות הירוקים הופכים לצהובים ואדומים.
התנועה המתמדת של התא. אברונים של תנועה.תאים רבים של חד תאיים ו אורגניזמים רב תאייםיש את היכולת לנוע. זה מתייחס הן לתנועת התא בחלל והן לתנועה התוך תאית של האברונים שלו. בתווך נוזלי, תנועת התאים מתבצעת על ידי תנועת דגלים וריסים (איור 10, 2, 8, 11). אורגניזמים חד תאיים רבים נעים בצורה זו, למשל, euglena ירוק, flagellate, infusoria וכו'. סוגים מסוימים של חיידקים נעים גם בעזרת flagella, ארוכות וגמישות, המסתובבות במהירות, מה שמבטיח את התקדמות התא. אמבה וכמה פרוטוזואה אחרות, כמו גם תאים מיוחדים של אורגניזמים רב-תאיים (לדוגמה, לימפוציטים) נעים בעזרת יציאות שנוצרו על פני התאים.
התא נמצא בפנים בתנועה מתמדת. במהלך phagocytosis ו-pinocytosis, קרום הפלזמה בולט לתוך התא, ליזוזומים, שלפוחיות של קומפלקס Golgi, מיטוכונדריה נע, ולבסוף, הציטופלזמה עצמה נעה.
התנועה הסלולרית מסופקת על ידי שלד ציטובולי המורכב ממיקרוטובולים, מיקרופילמנטים ו מרכז תאים. מיקרוטובולים הם גלילים חלולים ארוכים בקוטר של 25 ננומטר, שדפנותם מורכבות מחלבונים. הדגלים והריסים של תאי בעלי חיים וצמחים מורכבים ממיקרוטובולים מקבילים. Flagella שונה מסיליה רק באורך. לפיכך, לזרע של יונקים יש דגל אחד באורך של עד 100 מיקרון. ריסים קצרים מהדגלים ביותר מפי 10; יש כמה אלפי ריסים לתא. מיקרופילמנטים הם מבנים דקים מאוד המורכבים מאלפי מולקולות חלבון אקטין המחוברות זו לזו. בתאי שריר הם נכנסים, יחד עם חוטי חלבון נוספים - מיוזין, לקומפלקסים המספקים את תפקוד ההתכווצות של תאים אלו.
בציטופלזמה של התאים של כל האורגניזמים, ליד הגרעין, יש מרכז תאים שלוקח חלק בחלוקת התא. הרכב מרכז התא של תאי בעלי חיים וכמה צמחים כולל צנטרול (איור 11, 3). Centriole - היווצרות זוג. הוא מכיל שני גלילים העשויים ממיקרוטובולים ומסודרים בניצב אחד לשני.
תכלילים של תאים. לבסוף, יש לומר על התכלילים הרבים בציטופלזמה. תכלילים נקראים מבנים לא קבועים של הציטופלזמה, שבניגוד לאברונים, מופיעים או נעלמים במהלך חיי התא. תכלילים צפופים בצורת גרגירים מכילים רכיבי תזונה רזרביים (עמילן, חלבונים, סוכרים, שומנים) או תוצרי פסולת מהתאים שמסיבה זו או אחרת לא ניתן להסירם מיד. לכל הפלסטידים של תאי הצמח יש את היכולת לסנתז ולצבור חומרי הזנה רזרבה.
IN תאי צמחיםהצטברות של חומרי הזנה רזרבה מתרחשת גם ב-vacuoles - שקיות ממברנה עם תמיסה מימיתמלחים ותרכובות אורגניות, שלעתים קרובות תופסות כמעט את כל נפח התא, דוחפים את הגרעין והציטופלזמה אל קרום הפלזמה.
- מדוע המיטוכונדריה נקראות "תחנת הכוח" של התא?
- אילו מבנים של התא תורמים לתנועתו?
- מה זה תכלילים סלולריים? מה תפקידם בתא?
- תאר את המאפיינים המבניים של מיטוכונדריה וכלורופלסט בקשר לתפקודיהם בתא.
מרכז סלולר- אברון הנראה תחת מיקרוסקופ אופטי בתאי בעלי חיים ו צמחים נמוכים יותר. הוא ממוקם בדרך כלל ליד הגרעין או במרכז הגיאומטרי של התא ומורכב משני גופים דמויי מוט של צנטריולים, בגודל של כ-0.3-1 מיקרון. במיקרוסקופ אלקטרוני, נמצא שהצנטריול הוא גליל, שקירותיו בנויים על ידי תשעה זוגות של צינורות דקים מאוד. באמצע הגליל יש חלל מלא במסה הומוגנית. זוג סנטריולים מוקף באזור קל יותר - הצנטרוספירה.
מרכז התא משחק תפקיד חשובבמהלך תנועת הכרומוזומים במהלך מיטוזה. היכולת של חלק מהתאים לתנועה פעילה קשורה אליו. זה מוכח על ידי העובדה כי בבסיס של flagella או cilia של תאים תנועתיים (פרוטוזואה, spermatozoa) יש תצורות של אותו מבנה כמו מרכז התא.
אברונים של תנועה.תאים יכולים לנוע בעזרת אברונים מיוחדים, הכוללים ריסים ודגלים. ריסי התאים הם תמיד רבים (בפרוטוזואה מספרם הוא במאות ואלפים), והאורך הוא 10-15 מיקרון. Flagella הם לרוב 1-8, אורכם הוא 20-50 מיקרון. המבנה של ריסים ודגלים בתאי צמחים ובעלי חיים דומה. תחת מיקרוסקופ אלקטרונים, נמצא שמיקרוטובולים עוברים לכל אורכם. שניים מהם ממוקמים במרכז, ומסביבם לאורך הפריפריה משתרעים עוד 9 זוגות של מיקרוטובולים. כל המבנה הזה מכוסה ממברנה ציטופלזמיתשהיא הרחבה של קרום התא. תנועת הדגלים והסיליה מספקת לא רק את תנועת התאים במרחב, אלא גם את התנועה חומרים שוניםעל פני התאים, כמו גם כניסת חלקיקי מזון לתא. בבסיס הריסים והדגלים נמצאים גופים בזאליים, המורכבים גם ממיקרו-צינוריות. הוא האמין כי הגופים הבסיסיים הם מרכז היווצרות של microtubules של flagella ו cilia. גופים בזאליים, בתורם, מקורם לעתים קרובות ממרכז התא.
למספר רב של אורגניזמים חד-תאיים ולחלק מהתאים הרב-תאיים אין אברונים מיוחדים של תנועה והם נעים בעזרת פסאודופודיה (פסאודופודיה). תנועה בעזרת פסאודופודיה נקראת תנועת אמבואידים. הוא מבוסס על תנועה של מולקולות של חלבונים מיוחדים, המכונים חלבונים מתכווצים.
תכליליםנמצא לרוב בתאי צמחים ונבדלים מאברונים בכך שהם תצורות זמניות, לא קבועות. מספרם תלוי בעוצמת חילוף החומרים ובמצב הגוף. בדרך כלל הם נראים במיקרוסקופ אופטי בצורה של גרגרים או טיפות בגדלים וצורות שונות. על ידי תרכובת כימיתלהבחין: תכלילים של פחמימות, שומן וחלבון. תכלילים של פחמימות וחלבונים נראים כמו דגנים, ותכלילים שומניים נראים כמו טיפות. בתאי הצמח, פחמימות מופקדות לרוב בצורה של גרגרי עמילן, ובבעלי חיים - גליקוגן. גרגרי חלבון נמצאים בכמויות גדולות בציטופלזמה של ביצי בעלי חיים בצורה של חלמון. רבים מהם נמצאים בזרעים של צמחים. טיפות שומן קיימות בתאים רקמת חיבורבעלי חיים וזרעי צמחים. תאי צמחים מכילים תכלילים גבישיים (מלחים חומצה אורגנית). בתנאים מסוימים, כל סוגי התכלילים יכולים לשמש את התא במהלך פעילות חייו, ואז להצטבר שוב.
בציטופלזמה של תאי צמחים רבים יש תצורות מיוחדות - וואקוולים. הם טנקים מלאים בתערובת של תמיסות של כימיקלים שונים. ואקואולים גדולים במיוחד נוצרים בתאים ישנים.
|
קריטריונים לצורך השוואה |
סוגי ריבוזומים |
|
|
סוג פרוקריוטי |
סוג אוקריוטי |
|
|
מִמצָא בתאים |
ריבוזומים של אובקטריה וציאנובקטריה ריבוזומים פלסטידים מיטוכונדריה ריבוזומים Archaebacteria ריבוזומים (בדומה לאוקריוטי) |
ציטופלזמה ריבוזומים אוקריוטיים: בעלי חיים, וגם צמחים |
|
יַחַס RNA: חלבון | ||
|
כמות rRNA יחידת משנה |
1 מולקולה באורך 1500-1600 נוקלאוטידים (מינימום - 950 בריבוזומים מיטוכונדריאלים) |
1 מולקולה באורך של כ-1800 נוקלאוטידים |
|
כמות rRNA בגדול יחידת משנה |
2 מולקולות: מולקולה אחת באורך של כ-3000 נוקלאוטידים, 1 מולקולה באורך של כ-120 נוקלאוטידים |
3 מולקולות: מולקולה אחת באורך של כ-4-5 אלף נוקלאוטידים, מולקולה אחת באורך של כ-120 נוקלאוטידים, מולקולה אחת באורך של כ-160 נוקלאוטידים (מולקולה זו דומה לאחד מהאזורים של rRNA פרוקריוטי) |
|
חלבון (כמעט כל המולקולות ייחודיות) |
כ-20 מולקולות בתת-היחידה הקטנה ו-30 מולקולות בגדולה |
כ-30 מולקולות לכל תת-יחידה קטנה ו-40 מולקולות בגדול |
מרכז סלולר
מרכז התא הוא אברון השולט על היווצרות וצמיחתן של מיקרוטובוליות בשלד הציטוס של תא איקריוטי.(איור 38)
אורז. 38. מרכז סלולר 1 - שלישיות של microtubules; 2 - חישורים רדיאליים; 3 - המבנה המרכזי של "גלגל העגלה"; 4 - לוויין; 5 - ליזוזום; 6 - דיקטיוזומים של תסביך גולגי; 7 - שלפוחית גובלת; 8 - בור של רטיקולום אנדופלזמי גרגירי; 10 - מיטוכונדריה; 11 - גוף שיורי; 12 - microtubules; 13 - caroteca (לפי ר' Krstic, עם שינויים).
התפקידים העיקריים של מרכז התא הם יצירת מערכות של מיקרוטובולים של השלד הציטוניים, אברוני התנועה וציר החלוקה.
הבסיס של מרכז התא הוא זוג צנטריולים (לעיתים מספרם מגיע ל-4-6-8 בתאים פוליפלואידים), הממוקמים במרכז התא.
סנטריולים התגלו ותוארו לראשונה על ידי V. Flemming (1875), אך הם נחקרו בפירוט רק בעזרת מיקרוסקופ אלקטרונים. Centrioles קשורים קשר הדוק מעטפת גרעין, ובאאוקריוטים תחתונים רבים (פרוטוזואה, אצות, פטריות, מסוקריוטות), צנטרולים בנויים בקליפת הגרעין.
מרכז התא נמצא תמיד בתאים של בעלי חיים רב-תאיים. לפרוקריוטים חסר מרכז תאים וטובולין. בתאים של פטריות, אצות וחיות חד-תאיות לא תמיד נמצא מרכז התא, אלא בתאים צמחים גבוהים יותרלא נמצא (למעט חריגים נדירים). בהיעדר מרכז תאי, תפקידיו באאוקריוטים מבוצעים על ידי המרכז ליצירת מיקרוטובולים - CMTC.
צנטרולהוא גליל חלול בקוטר של כ-0.15 מיקרון ואורך של 0.3-0.5 מיקרון (פחות תכופות - כמה מיקרון). הקירות של הצנטריולים מורכבים מ-9 שלישיותמיקרוטובולים של טובולין המחוברים על ידי גשרי דיניין. בדרך כלל, צנטריולים מסודרים בזוגות: צנטרול אחד הוא צנטרול האב, והשני הוא צנטרול הבת; צנטרול הבת מאונך למרכז האב. צמד הצנטריולים הזה דיפלוזום- בעל צורה בצורת T או בצורת L. הדיפלוזום מוקף בשכבה צפופה של ציטופלזמה צנטרוספירה, או צנטרוזום. במהלך חלוקת התא, גדילים צפופים של מיקרוטובולים יוצאים מהצנטרוספרה, נוצרים אסטרוספרה.
הכפלה של צנטרול מתרחשת בערך באמצע השלב הבין-פאזי: הצנטרולים האימהיים והבתים מתפצלים, וניצני צנטרול חדש יוצא מכל צנטרול. בְּ קבוצות שונותאורגניזמים, לתהליך זה יש מאפיינים משלו.
בנוסף לאברונים, תאים מכילים תכלילים תאיים. הם יכולים להיכלל לא רק בציטופלזמה, אלא גם באברונים מסוימים, כגון מיטוכונדריה ופלסטידים.
מהם תכלילים של תאים?
אלו תצורות שאינן קבועות. בניגוד לאורגנואידים, הם אינם יציבים באותה מידה. בנוסף, יש להם מבנה הרבה יותר פשוט ומבצעים פונקציות פסיביות, כמו למשל גיבוי.
איך הם בנויים?
לרובם יש צורה בצורת טיפה, אך חלקם עשויים להיות אחרים, למשל, דומים לכתם. לגבי הגודל, זה עשוי להשתנות. תכלילים תאיים יכולים להיות קטנים יותר מאברונים, או באותו גודל או אפילו גדולים יותר.
הם מורכבים בעיקר מחומר אחד ספציפי, ברוב המקרים אורגני. זה יכול להיות שומן או פחמימה או חלבון. 
מִיוּן
תלוי מהיכן מגיע החומר ממנו הם מורכבים, ישנם סוגים הבאים של תכלילים תאיים:
- אקסוגני;
- אנדוגני;
- נְגִיפִי.
תכלילים תאיים אקסוגניים בנויים תרכובות כימיותשנכנסו לתא מבחוץ. אלה שנוצרים מחומרים המיוצרים על ידי התא עצמו נקראים אנדוגניים. תכלילים ויראליים, למרות שהם מסונתזים על ידי התא עצמו, עם זאת, הדבר מתרחש כתוצאה מה-DNA של הנגיף שנכנס אליו. התא פשוט לוקח אותו עבור ה-DNA שלו ומסנתז ממנו את חלבון הנגיף.
בהתאם לתפקודים שמבצעים תכלילים של תאים, הם מחולקים לפיגמנט, הפרשה וטרופי.
יתר על כן, תכלילים מחולקים לסוגים בהתאם לתרכובות הכימיות הספציפיות מהן הם מורכבים. 
תכלילים סלולריים: פונקציות
יכולות להיות להם שלוש פונקציות. שקול אותם בטבלה
כל אלו הן פונקציות של תצורות לא קבועות בתא.
תכלילים של תאי בעלי חיים
הציטופלזמה של החיה מכילה תכלילים טרופיים ופיגמנטים כאחד. לחלק מהתאים יש גם תאי הפרשה.
טרופי בתאי בעלי חיים הם הכללת גליקוגן. יש להם צורה של גרגיר בגודל של כ-70 ננומטר. 
הגליקוגן הוא חומר העתודה העיקרי של החיה. כפי ש חומר נתוןהגוף אוגר גלוקוז. ישנם שני הורמונים המווסתים את חילוף החומרים של גלוקוז וגלוקוגן: אינסולין וגלוקגון. שניהם מיוצרים על ידי הלבלב. אינסולין אחראי ליצירת גליקוגן מגלוקוז, וגלוקגון, להיפך, מעורב בסינתזה של גלוקוז.
רוב תכלילי הגליקוגן נמצאים בתאי כבד. הם גם נמצאים בכמויות גדולות בהרכב השרירים, כולל הלב. לתכלילים של גליקוגן של תאי כבד יש צורה של גרגירים בגודל של כ-70 ננומטר. הם מתאספים באשכולות קטנים. תכלילים של גליקוגן של מיוציטים ( תאי שריר) הם עגולים בצורתם. הם בודדים, מעט יותר גדולים מהריבוזומים.
כמו כן, תאים של בעלי חיים מאופיינים תכלילים של שומנים. אלו הם גם תכלילים טרופיים, שבזכותם הגוף יכול לקבל אנרגיה פנימה חירום. הם מורכבים משומנים ובעלי צורת דמעה. בעיקרון, תכלילים כאלה כלולים בתאי רקמת החיבור השומנית - ליפוציטים. ישנם שני סוגים של רקמת שומן: לבן וחום. ליפוציטים לבנים מכילים טיפה אחת גדולה של שומן, תאים חומים מכילים מספר קטן של תאים.
באשר לתכלילים של פיגמנט, תאי בעלי חיים מאופיינים באלה המורכבים ממלנין. הודות לחומר זה, לקשתית העין, לעור ולחלקים אחרים של הגוף יש צבע מסוים. ככל שיותר תכלילים של מלנין בתאים, כך מורכב התאים האלה כהה יותר.
פיגמנט נוסף שניתן למצוא בתאי בעלי חיים הוא lipofuscin. חומר זה הוא בצבע חום-צהוב. זה מצטבר בשריר הלב ובכבד עם הזדקנות האיברים.
תכלילים של תאי צמחים
תכלילים סלולריים, שהמבנה והתפקודים שלהם אנו שוקלים, כלולים גם הם
התכלילים הטרופיים העיקריים באורגניזמים אלה הם גרגירי עמילן. בצורתם, צמחים אוגרים גלוקוז. בדרך כלל, תכלילי עמילן הם עדשים, כדוריים או ביציים בצורתם. גודלם יכול להשתנות בהתאם לסוג הצמח ולאיבר שבתאיו הם נמצאים. זה יכול להיות מ-2 עד 100 מיקרון.
תכלילים ליפידיםמאפיין גם תאי צמחים. הם התכלילים הטרופיים השני בשכיחותו. יש להם צורה כדורית וקרום דק. הם נקראים לפעמים ספרוזומים.
תכלילים של חלבוןקיימים רק בתאי צמחים, הם אינם אופייניים לבעלי חיים. הם מורכבים מחלבונים פשוטים הנקראים חלבונים. תכלילי חלבון הם משני סוגים: גרגרי אלורון וגופי חלבון. גרגרי Aleurone עשויים להכיל גבישים או פשוט חלבון אמורפי. אז, הראשון נקרא מורכב, והשני - פשוט. גרגרי אלורון פשוטים, המורכבים מחלבון אמורפי, שכיחים פחות.
באשר תכלילים פיגמנט, צמחים מאופיינים על ידי פלסטוגלובולים. הם מכילים קרוטנואידים. תכלילים כאלה אופייניים לפלסידים.
תכלילים תאיים, שאת המבנה והתפקודים שלהם אנו בוחנים, מורכבים ברובם מתרכובות כימיות אורגניות, אולם בתאי הצמח יש גם כאלה הנוצרים מ. לֹא חומר אורגני. זֶה גבישי סידן אוקסלט. 
הם נמצאים רק ב-vacuoles של התא. גבישים אלה יכולים להיות מהצורה המגוונת ביותר, ולעתים קרובות זה אינדיבידואלי עבור סוגים מסוימיםצמחים.
לאורגניזמים חיים המורכבים מתא אחד יש לעתים קרובות את היכולת לנוע באופן פעיל. מנגנוני התנועה שנוצרו בתהליך האבולוציה מגוונים מאוד.
אברוני התנועה הם פסאודופודיה (בסרקודים ובכמה דגלים), דגלים (בדגלים) או ריסים (בצליות).
בנוסף, תנועה יכולה להתבצע כתוצאה מהתכווצות של מיקרופייבר מיוחדים - מיונמות, הממוקמות מתחת לכיסויים בציטופלזמה (ב-apicomples)
תנועת האמבואידים קיבלה את שמה מהאורגניזם הפשוט ביותר - האמבה. (ראה נספח מס' 6).איברי התנועה באמבה הם רגליים מזויפות - פסאודופודיה, שהן בליטות של הציטופלזמה. הם נוצרים פנימה מקומות שוניםפני השטח של הציטופלזמה. הם יכולים להיעלם ולהופיע שוב במקום אחר.
פסאודופודיה(מפסאודו... ומוגלה יוונית, סוג p. podos - כף הרגל) (פסאודופודים), יציאות ציטופלזמיות זמניות באורגניזמים חד-תאיים (קני שורש, ספורוזואנים וכו') ובחלק מהתאים (לויקוציטים, מקרופאגים וכו') של אורגניזמים רב-תאיים. . הם משמשים לנוע וללכוד מזון וחלקיקים אחרים.
תנועה בעזרת דגלים אופיינית לאצות חד-תאיות רבות (לדוגמה, כלמידומונס), פרוטוזואה (לדוגמה, יוגלנה ירוקה) וחיידקים. איברי התנועה באורגניזמים אלה הם דגלים - יציאות ציטופלזמה על פני הציטופלזמה.
פלאגלה (ראה נספח מס' 7). גדלים מ-2-5 מיקרון עד 1 מ"מ. הם נעים בעזרת דגלים אחד או רבים, שאורכם אצל פרט אחד עשוי להיות שונה. רק מעט מאוד מינים יכולים ליצור פסאודופודיה. הם בדרך כלל מתרבים באופן א-מיניכתוצאה מחלוקת תאים אורכית; חלק ראו רבייה מינית. הרוב המכריע של הדגלים הם מינים חיים חופשיים המצויים במים ימיים ומתוקים, כמו גם באדמה. צורות פלנקטוניות ממלאות תפקיד חשוב במחזור החומר; סוגים רבים הם אינדיקטורים ביולוגייםזיהום מים.
מבנה התא.
תא, מערכת חיים יסודית, בסיס המבנה והחיים של כל בעלי החיים והצמחים. תאים קיימים כאורגניזמים עצמאיים (לדוגמה, פרוטוזואה, חיידקים) וכחלק מאורגניזמים רב-תאיים, בהם יש תאי מין המשמשים לרבייה, ותאי גוף (סומטיים), שונים במבנה ובתפקודים (למשל, עצב, עצם). , שריר , הפרשה).
גודל התאים נע בין 0.1-0.25 מיקרון (חיידקים מסוימים) ל-155 מ"מ (ביצת יען בקליפה).
התא של כל אורגניזם הוא שלם מערכת חיה. הוא מורכב משלושה חלקים הקשורים זה לזה: ממברנה, ציטופלזמה וגרעין. (ראה נספח מס' 9).
1. קרום התא.
קרום התא מקיים אינטראקציה ישירה עם הסביבה החיצונית ומקיים אינטראקציה עם תאים שכנים (באורגניזמים רב-תאיים).
לדופן התא יש מבנה מורכב. הוא מורכב משכבה חיצונית וממברנת פלזמה הממוקמת מתחת.
תאי בעלי חיים וצמחים נבדלים זה מזה במבנה השכבה החיצונית שלהם (ראה נספח מס' 10).בצמחים, כמו גם בחיידקים, אצות כחולות ירוקות ופטריות, על פני התאים ממוקם קליפה צפופה, או דופן התא. ברוב הצמחים הוא מורכב מסיבים. דופן התא ממלאת תפקיד חשוב ביותר: היא מסגרת חיצונית, מעטפת הגנה, מספקת טורגור של תאים צמחיים: מים, מלחים, מולקולות של חומרים אורגניים רבים עוברות בדופן התא.
השכבה החיצונית של פני השטח של תאי בעלי חיים, בניגוד לדפנות התא של צמחים, דקה מאוד ואלסטית. הוא אינו נראה במיקרוסקופ אור ומורכב ממגוון רחב של פוליסכרידים וחלבונים. שכבת פני השטח של תאי בעלי חיים נקראת גליקוקליקס.
Glycocalyx מבצעת בעיקר את הפונקציה של חיבור ישיר של תאי בעלי חיים עם הסביבה החיצונית, עם כל החומרים הסובבים אותה. בעל עובי לא משמעותי (פחות מ-1 מיקרון) / השכבה החיצונית של תא החי אינו ממלא תפקיד תומך, האופייני לדפנות התא של צמחים. היווצרות הגליקוקאליקס, כמו גם דפנות התא של צמחים, מתרחשת עקב הפעילות החיונית של התאים עצמם.
2. ציטופלזמה.
ציטופלזמה (מציטו ... ופלזמה), המונח "ציטופלזמה" הוצע על ידי E. Strasburger (1882).
מופרד מ סביבה חיצוניתממברנת הפלזמה, הציטופלזמה היא הסביבה הפנימית הנוזלת למחצה של תאים (ראה נספח מס' 8).בציטופלזמה תאים איקריוטייםממוקמים הגרעין והאברונים השונים.
הגרעין ממוקם בחלק המרכזי של הציטופלזמה. הוא מכיל גם מגוון תכלילים - תוצרים של פעילות תאית, ואקוולים, כמו גם הצינורות והחוטים הקטנים ביותר היוצרים את שלד התא.
ואקוולים (ראה נספח מס' 8)(בצרפתית vacuole, מלטינית vacuus - ריק), חללים בתאי בעלי חיים וצמחים או אורגניזמים חד תאיים. ישנם ואקוולים עיכול ומתכווצים (פועם) המווסתים את הלחץ האוסמוטי ומשמשים להסרת תוצרי ריקבון מהגוף. (ראה נספח מס' 11).
חלבונים שולטים בהרכב החומר העיקרי של הציטופלזמה.
התהליכים המטבוליים העיקריים מתרחשים בציטופלזמה, היא מאחדת את הגרעין ואת כל האברונים למכלול אחד, מבטיחה את האינטראקציה ביניהם, את פעילות התא כמערכת חיה אינטגרלית אחת.
בניסוי, ניתן להשיג תאי ציטופלס חיים שאינם גרעיניים, שיכולים לסנתז חלבונים, שומנים, ATP תוך 1-3 ימים. ואז, כמובן, הם מתים בגלל חוסר האפשרות לסנתז RNAs חדשים בהיעדר גרעין.
נפח הציטופלזמה תאים שוניםמשתנה: בלימפוציטים הוא שווה בערך לנפח הגרעין, ובתאי כבד הציטופלזמה מהווה 94% מנפח התא הכולל.
באופן פורמלי, שלושה חלקים נבדלים בציטופלזמה: אברונים, תכלילים והיאלופלזמה.
אברונים- רכיבים, חובה לכל תא, שבלעדיהם התא אינו יכול לתמוך בקיומו.
האברונים החד-ממברניים של המערכת הוואקואולרית כוללים את הרטיקולום האנדופלזמי, ליזוזומים, מנגנון גולגי, פרוקסיזומים וממברנת הפלזמה.
אברונים דו-ממברניים הם מיטוכונדריה ופלסטידים. קבוצה זו כוללת גם גרעין התא.
רכוש משותףאברוני הממברנה היא שהם בנויים ממברנות ליפופרוטאין שסוגרות על עצמן, כך שהם יוצרים חללים סגורים ובכך מחלקים את הציטופלזמה לקבוצות של תאים שונים מבחינה תפקודית.
ריבוזומים, צנטרוזומים של תאי בעלי חיים שייכים לאברונים שאינם ממברניים.
הכללות_ _ לא תמיד נמצאים ומייצגים משקעים של חומרי רזרבה (גליקוגן, חלמון) או הצטברויות של מוצרים מטבוליים (פיגמנטים, גבישי מלח בצמחים).
היאלופלזמה(מ"hualine" - שקוף) - זו הפלזמה העיקרית, ציטוזול, נכון סביבה פנימיתתאים. הרכב ההיאלופלזמה מורכב מאוד, והעקביות מתקרבת לג'ל.
ג'לים- מערכות קולואידיות מובנות עם מדיום פיזור נוזלי, אשר בהשפעת חיצוני או גורמים פנימייםיכולים לשנות את מצב הצבירה שלהם ולעבור לפאזה נוזלית יותר - סול. מעברי ג'ל-סול דומים יכולים להתרחש בציטופלזמה בהשפעת חלבון האקטין, ומצבו משתנה בחלקים שונים של התא, מה שמבטיח את תנועת התא כולו. כאשר האקטין הפיברילרי יוצר אינטראקציה עם חלבונים מסוג פיברין, הג'ל מתייצב, וכאשר הוא נקשר לחלבונים שפעילותם תלויה בריכוז יוני Ca (למשל ג'סולין), המערכת כולה עוברת למצב נוזלי.
חומרת המרכיבים של שלד התא, כולל מרכיב האקטין שלו, יכולה להשתנות באופן משמעותי במהלך מחזור התא.
המשמעות התפקודית של הציטופלזמה גבוהה מאוד.
היאלופלזמה, בנוסף ליונים שונים של תרכובות אנאורגניות, מכילה אנזימים המעורבים בסינתזה של חומצות אמינו, נוקלאוטידים, חומצות שומן, סוכרים. על ריבוזומים ופוליריבוזומים היושבים על ממברנות, מסונתזים חלבונים שונים המספקים חילוף חומרים תאי.
3. רטיקולום אנדופלזמי.
כל האזור הפנימי של הציטופלזמה מלא בתעלות קטנות ובחללים רבים, שקירותיהם הם ממברנות הדומות במבנה לממברנת הפלזמה. ערוצים אלו מסתעפים, מתחברים זה לזה ויוצרים רשת הנקראת הרשת האנדופלזמית או הרשת האנדופלזמית.
רשת אנדופלזמית (ראה נספח מס' 12)הטרוגני במבנה. ידועים שני סוגים שלו - גרגירי (גס) וחלק.
על ממברנות התעלות והחללים של הרשת הגרעינית יש הרבה גופים עגולים קטנים - ריבוזומים, המקנים לממברנות מראה מחוספס.
הממברנות של הרשת האנדופלזמית החלקה אינן נושאות ריבוזומים על פני השטח שלהן.
הרשת האנדופלזמית מבצעת פונקציות רבות ושונות. הפונקציה העיקרית של הרטיקולום האנדופלזמי הגרגירי היא השתתפות בסינתזת חלבון, המתבצעת בריבוזומים.
על הממברנות של הרשת האנדופלזמית החלקה, מסונתזים שומנים ופחמימות. כל מוצרי הסינתזה הללו מצטברים בתעלות ובחללים, ולאחר מכן מועברים לאברוני תאים שונים, שם הם נצרכים או מצטברים בציטופלזמה כתכלילים של תאים.
הרטיקולום האנדופלזמי מחבר בין האברונים העיקריים של התא (ראה נספח מס' 8).
4. ממברנות ביולוגיות.
ממברנות ביולוגיות (לט. "ממברנה" - עור, סרט), סרטי ליפופרוטאין דקים (לא יותר מ-10 ננומטר עובי), המורכבים משכבה כפולה של מולקולות שומנים, הכוללת מולקולות של חלבונים שונים. הם ממוקמים על פני השטח של תאים (ממברנת פלזמה) וחלקיקים תוך תאיים (גרעינים, מיטוכונדריה וכו'). במונחי משקל, בהתאם לסוג הממברנה, שומנים מהווים 25-60%. וחלקם של חלבונים - 40-75%. ממברנות רבות מכילות פחמימות, שכמותן יכולה להגיע ל-2-10%.
בריאות התא, משך חייו תלוי במידה רבה במצב הממברנה.
א) מאפיינים של ממברנות.
חדירות מלאה למים. הממברנה תמיד מכניסה מים לתא או החוצה, תלוי היכן ריכוז המים גדול יותר. תנועה זו של חומר מאזור בריכוז גבוה לאזור בריכוז נמוך יותר נקראת דיפוזיה. פיזור של חומר אינו דורש אנרגיה.
מוליכות סלקטיבית של מומסים:
חלקיקים בעלי מטען שלילי חודרים את הממברנה מהר יותר ובקלות רבה יותר. חומרים מסיסים בשומנים חודרים לממברנה ביתר קלות מאשר חומרים מסיסים במים. מולקולות קטנות חודרות את הממברנה בקלות רבה יותר מאשר גדולות.
ב) הובלה פעילה של חומרים.
חלק מהחומרים מסוגלים לחדור לממברנה בכיוון המנוגד לדיפוזיה שלהם, כלומר ממקום של נמוך למקום עם יותר ריכוז גבוה. על ידי הובלה פעילה, עודפי נתרן, מימן וכלור מוסרים כל הזמן מהתא. ופוספטים, גלוקוז, חומצות אמינו, להיפך, חודרים באופן פעיל לתוך הציטופלזמה. הובלה פעילה קשורה תמיד להוצאת אנרגיה.
הממברנה עלולה ללכת לאיבוד חלקי במהלך חיי התא, כתוצאה מהיווצרות ואקוולים (שלפוחיות) עיכול. הממברנה משוחזרת באופן קבוע כתוצאה מעבודתם של אברונים מיוחדים. סינתזה של ואקואולים של ממברנות. ואקואולים אלה נדבקים בכל מקום בקרום התא, ומשחזרים את מימדיו ותכונותיו הקודמים.
ג) שומני ממברנה.
הרכב ליפידים ממברנות ביולוגיותמגוונים מאוד. נציגים אופיינייםשומנים בקרום התא הם פוספוליפידים, ספינגומיאלינים וכולסטרול (שומנים סטרואידים).
תכונה אופייניתשומני הממברנה היא חלוקת המולקולות שלהם לשני חלקים שונים בתפקוד: זנבות לא קוטביים, נטולי מטען, המורכבים מחומצות שומן וראשים קוטביים טעונים.
ראשי קוטב נושאים מטענים שלילייםאו עשוי להיות ניטרלי. נוכחותם של זנבות לא קוטביים מסבירה את המסיסות הטובה של שומנים בשומנים ובממיסים אורגניים. בניסוי, על ידי ערבוב ליפידים מבודדים ממברנות עם מים, ניתן להשיג שכבות דו-מולקולריות או ממברנות בעובי של כ-7.5 ננומטר, כאשר האזורים ההיקפיים של השכבה הם ראשי קוטב הידרופיליים, והאזור המרכזי הוא זנבות לא טעונים של מולקולות שומנים. . לכל ממברנות התא הטבעיות יש אותו מבנה.
ממברנות התא משתנות מאוד בהרכב השומנים. לדוגמה, ממברנות פלזמהתאי בעלי חיים עשירים בכולסטרול (עד 30%) ויש בהם מעט לציטין, בעוד שקרומי המיטוכונדריה עשירים בפוספוליפידים ועניים בכולסטרול.
מולקולות ליפידים יכולות לנוע לאורך שכבת השומנים, יכולות להסתובב סביב הציר שלהן, וגם לנוע משכבה לשכבה. לחלבונים הצפים ב"אגם השומנים" יש גם ניידות לרוחב. הרכב השומנים משני צידי הממברנה שונה, מה שקובע את חוסר הסימטריה במבנה השכבה הביליפידית.
ד) חלבוני ממברנה.
מולקולות חלבון, שהן הרבה יותר כבדות וגדולות ממולקולות שומנים, מפוזרות, כביכול, בשכבת הביליפידית של הממברנה. רוב מולקולות החלבון טבולות חלקית בממברנה וקשורות לליפידים באמצעות אינטראקציות הידרופוביות. אלו הם מה שנקרא חלבונים אינטגרליים.
חלבונים אחרים קשורים רק ל"קצוות" הקוטביים של מולקולות השומנים וממוקמים על פני השכבה הביליפידית, בעוד שאחרים חודרים לממברנה דרך ודרך. אסימטריה במבנה הממברנות באה לידי ביטוי בצורה הברורה ביותר ביחס לחלבונים אינטגרליים. הם מכוונים בקפדנות: מסופי ה-N שלהם מסתכלים לתוך חלל ה-vacuoles או לתוך הסביבה החוץ-תאית.
ממברנות שונות מאוד בהרכב החלבונים, אשר למעשה קובעים את תכונותיהם התפקודיות. ניתן לחלק את כל חלבוני הממברנה לשלוש קבוצות: אנזימים, חלבוני קולטן וחלבונים מבניים.
לממברנות שונות יש קבוצה אופיינית של אנזימים. לדוגמה, ATPase תלוי K-Na, המעורב בהובלת יונים, ממוקם בממברנת הפלזמה. חלבוני קולטנים נקשרים ספציפית לחומרים מסוימים וכביכול "מזהים" אותם: אלו הם חלבוני קולטן להורמונים, לזיהוי פני השטח של תאים שכנים, וירוסים וכו'.
חלבונים מבניים מספקים חוזק ממברנה וקשורים למגוון מבני חלבון בציטופלזמה. לדוגמה, בתאי אפיתל, חלבוני ממברנת פלזמה מיוחדים נקשרים לאלמנטים של השלד הציטוניים ומשתתפים ביצירת מספר קשרים בין-תאיים, כגון דסמוזומים, מגעים דביקים וכו'.
ה) פחמימות ממברנה.
הם קשורים בעיקר לממברנות. גליקופרוטאין - מולקולות של חלבונים המקושרות באופן קוולנטי לשרשרות של פחמימות. ככלל, מולקולות פחמימות ממוקמות בשכבות החיצוניות של הממברנות. לרוב, מדובר בשרשראות ליניאריות קצרות או מסועפות, הכוללות גלקטוז, מניוז, פרוקטוז, סוכרוז, N-אצטיל גלוקוזאמין. ערבינוז. קסילוס וכו'.
ו) סינתזה של ממברנות תאים.
כל ממברנות התא (למעט ממברנות מיטוכונדריה וממברנות פלסטידיות) בנויות ברשת האנדופלסמית הגרנולרית. מתפצלים ממנו ואקוולים קרומיים קטנים, המתמזגים עם הקרומים של מנגנון גולגי. שלפוחיות ממברנה, המיוצרות בתורן על ידי מנגנון גולגי, משמשות לבניית קרום הפלזמה, ואקוולי הפרשה וכל שאר מרכיבי הממברנה של התא.
5. קרום פלזמה.
קרום פלזמה (ראה נספח מס' 8)(plasmalemma, קרום התא), מתחת לגליקוקאליקס ו דופן תאממברנת הפלזמה ממוקמת, המקיפה את הפרוטופלזמה של תאי צמחים ובעלי חיים. עובי קרום הפלזמה הוא כ-10 ננומטר, חקר המבנה והתפקודים שלו אפשרי רק בעזרת מיקרוסקופ אלקטרונים.
משמש לא רק כמחסום מכני, אלא, והכי חשוב, מגביל את הזרימה הדו-כיוונית החופשית אל תוך התא ומחוצה לו של חומרים בעלי מולקולריים נמוכים וגבוהים. יתרה מכך, הפלזמלמה פועלת כמבנה ש"מזהה" שונות חומרים כימייםוויסות ההובלה הסלקטיבית של חומרים אלו לתוך התא. כמו קרומי תאים אחרים, הוא מתעורר ומתעדכן עקב פעילות סינתטיתרטיקולום אנדופלזמי ובעל מבנה דומה להם.
א) תפקיד הובלת מחסום של הפלזמה.
היציבות המכנית של ממברנת הפלזמה נקבעת לא רק על ידי תכונות הממברנה עצמה, אלא גם על ידי תכונות הגליקוקאליקס הסמוך לו והשכבה הקורטיקלית של הציטופלזמה.
פני השטח החיצוניים של קרום הפלזמה (ראה נספח מס' 13).מכוסה בשכבה סיבית רופפת של חומר בעובי 3-4 ננומטר - גליקוקליקס. הוא מורכב משרשראות פוליסכרידים מסועפות של חלבונים אינטגרליים של הממברנה, שביניהם ניתן לאתר גליקוליפידים ופרוטאוגליקנים המופרשים על ידי התא. כמה אנזימים הידרוליטיים תאיים המעורבים בפירוק חוץ תאי של חומרים (עיכול חוץ תאי, למשל, באפיתל המעי) נמצאים מיד.
שכבת קליפת המוח של הציטופלזמה, בעובי 0.1-0.5 מיקרומטר, אינה מכילה ריבוזומים ומבני ממברנה, אך עשירה במיקרופילמנטים של אקטין.
קרום הפלזמה, כמו ממברנות תאי ליפופרוטאין אחרות, חדיר למחצה. למים ולגזים המומסים בהם יש את עוצמת החדירה המקסימלית. הובלת יונים יכולה להתבצע באופן פסיבי, ללא צריכת אנרגיה. במקרה זה, חלק מחלבוני הובלת הממברנה יוצרים קומפלקסים מולקולריים, תעלות שדרכן עוברים יונים דרך הממברנה באמצעות דיפוזיה פשוטה.
במקרים אחרים, חלבוני נושאי ממברנה מיוחדים נקשרים באופן סלקטיבי ליון כזה או אחר ומעבירים אותו על פני הממברנה. סוג זה של העברה נקרא הובלה פעילה ומתבצע באמצעות משאבות יוני חלבון. לדוגמה, בהוצאת מולקולת ATP אחת, מערכת משאבת K-Na שואבת 3 יוני Na מהתא במחזור אחד ושואבת 2 יוני K כנגד שיפוע הריכוז.
בשילוב עם הובלה פעילה של יונים, סוכרים שונים, נוקלאוטידים וחומצות אמינו חודרים דרך הפלזמה. מקרומולקולות, כגון חלבונים, אינן עוברות דרך הממברנה. הם, כמו גם חלקיקים גדולים יותר של חומר, מועברים דרך התא אנדוציטוזיס . במהלך אנדוציטוזיס, קטע מסוים של הפלזמהלמה לוכד, עוטף את החומר החוץ-תאי וסוגר אותו ב-vacuole של ממברנה. ואקואול זה הוא אנדוזום - מתמזג בציטופלזמה עם הליזוזום הראשוני ומתרחש עיכול החומר הנלכד.
אנדוציטוזיס מתחלק רשמית לפאגוציטוזיס (ספיגה של חלקיקים גדולים על ידי התא) ולפינוציטוזיס (ספיגה של תמיסות).
קרום הפלזמה מעורב גם בסילוק חומרים מהתא בעזרת אקסוציטוזיס - התהליך ההפוך של אנדוציטוזיס.
ב) תפקיד הקולטן של הפלזמה.
חלבוני נשא של הממברנה החיצונית של התא הם גם קולטנים המזהים יונים מסוימים ומקיימים איתם אינטראקציה. חלבוני ממברנה או יסודות גליקוקליקס יכולים לפעול כקולטנים על פני התא. אתרים כאלה הרגישים לחומרים בודדים מפוזרים על פני התא או נאספים באזורים קטנים.
תפקידם של קולטני תאים רבים הוא לא רק לקשור חומרים ספציפיים, אלא גם להעביר אותות מהמשטח אל התא. לדוגמה, כאשר הורמון פועל על תא, שרשרת האירועים מתפתחת באופן הבא: מולקולת ההורמון מקיימת אינטראקציה ספציפית עם חלבון קולטן ממברנת הפלזמה, ומבלי לחדור לתא, מפעילה את האנזים המסנתז AMP מחזורי. האחרון מפעיל או מעכב אנזים תוך תאי או קבוצת אנזימים.
הגיוון והספציפיות של קבוצות הקולטנים על פני התא מובילים ליצירת מאוד מערכת מורכבתסמנים המאפשרים לתאים להבחין בין "שלהם" (מאותו פרט או מאותו מין) מ"זרים".
ג) קשרים בין תאיים.
באורגניזמים רב-תאיים נוצרים הרכבי תאים מורכבים עקב אינטראקציות בין-תאיות. בסמיכות של תאים זה לזה, הגליקוקליקס מבטיח הידבקות תאים עקב נוכחותם של גליקופרוטאינים טרנסממברניים קדרינים בו. זהו מגע בין תאי פשוט, שבו הפער בין התאים הוא 10-20 ננומטר.
באפיתל, יש לעתים קרובות חיבור צפוף, או נעילה, שבו השכבות החיצוניות של שני ממברנות הפלזמה קרובות ככל האפשר וכדוריות של חלבוני ממברנה אינטגרליות שוכנות בנקודות המגע שלהן. מגע כזה הוא בלתי חדיר למולקולות ויונים; הוא סוגר חללים בין-תאיים.
חיבורי עוגן, או אנשי קשר, לא רק מחברים את ממברנות הפלזמה של תאים שכנים, אלא גם נקשרים לאלמנטים הפיברילריים של השלד הציטוניים. לדוגמה, עבור דסמוזומים שנראים כמו פלאקים או כפתורים, החלל הבין תאי מאופיין בנוכחות של שכבה צפופה של גליקופרוטאין דסמוגליין.
בצד הציטופלזמי, שכבה של חלבון desmoplakin, הקשורה לחוטי הביניים של השלד הציטופלמי, צמודה לפלסמה.
גאפ junctions נחשבים לצמתי תקשורת של תאים. באזור מגע הפער יכול להיות בין 20-30 לכמה אלפים קשרים - מבני חלבון גליליים עם ערוץ פנימיקוטר של 2 ננומטר. כל קונקסון מורכב מ-6 תת-יחידות חלבון קונקטין. קונקסונים ממלאים את התפקיד של תעלות בין-תאיות ישירות שדרכן יכולים להתפזר יונים וחומרים בעלי משקל מולקולרי נמוך מתא לתא.
6. ליזוזומים.
ליזוזומים (מליז וסומא יוונית - גוף). הם גופים עגולים קטנים. כל ליזוזום מופרד מהציטופלזמה על ידי ממברנה. בתוך הליזוזום נמצאים אנזימים שיכולים לפרק (כלומר, ליזה - ומכאן השם) חלבונים, פוליסכרידים, פפטידים, חומצות גרעין (ראה נספח מס' 8).
זהו מחלקה מאוד מגוונת של שלפוחיות בגודל 0.1-0.4 מיקרומטר, מוגבלת על ידי קרום בודד (עובי של כ-7 ננומטר), עם תוכן הטרוגני בפנים. הם נוצרים על ידי פעילות הרשת האנדופלזמית ומנגנון הגולגי ומבחינה זו דומים ל-vacuoles הפרשה.
תפקידם העיקרי הוא השתתפות בתהליכי הביקוע התוך תאי של מקרומולקולות ביולוגיות אקסוגניות ואנדוגניות כאחד. תכונה אופייניתליזוזומים הוא שהם מכילים כ-40 אנזימים הידרוליטים: פרוטאינזים, נוקלאזות, פוספטאזות, גליקוזידאזות וכו', שהאופטימום שבהם מתבצע ב-pH5.
בליזוזומים, הערך החומצי של הסביבה נוצר עקב הימצאות בקרום שלהם של "משאבת" פרוטון הצורכת את האנרגיה של ATP.
בנוסף, חלבוני נשא בנויים בממברנות של הליזוזומים לצורך הובלה מהליזוזום לציטופלזמה של תוצרי הידרוליזה: מונומרים של מולקולות מפוצלות - חומצות אמינו, סוכרים, נוקלאוטידים, שומנים. כדי לא לעכל את עצמם, מרכיבי הממברנה של הליזוזומים מוגנים על ידי אוליגוסכרידים המונעים הידרולאזים לקיים איתם אינטראקציה.
בין חלקיקים ליזוזומליים בעלי מורפולוגיה שונה, מובחנים ארבעה סוגים: ליזוזומים ראשוניים ומשניים, אוטופגוזומים וגופים שיוריים.
ליזוזומים ראשוניים.
בדרך כלל מדובר בשלפוחיות קרומיות קטנות בקוטר של כ-100 ננומטר עם תכולה חסרת מבנה המכילה פוספטאז חומצה פעיל, אנזים סמן לליזוזומים. הוכח כי אנזימים ליזוזומים מסונתזים, כרגיל, ברטיקולום הגרגירי, ונארזים לתוך שלפוחיות ממברנות במנגנון גולגי. כל דרך היווצרותם של ליזוזומים ראשוניים דומה מאוד להיווצרות גרגירי הפרשה, למשל, בתאי הלבלב.
ליזוזומים משניים.
ואקוולים פגוציטים או פינוציטיים מתמזגים עם ליזוזומים ראשוניים ליצירת ליזוזומים משניים. מתחיל תהליך פיצול המצעים הנספגים בתא בפעולת ההידרולאזים הכלולים בליזוזום הראשוני. חומרים ביוגנים מתפרקים למונומרים, המועברים דרך ממברנת הליזוזום אל הציטופלזמה, שם הם מנוצלים מחדש, הכלולים במגוון סינטטי ו תהליכים מטבוליים. המגוון של הגודל והמבנה של ליזוזומים תאיים קשור בעיקר למגוון של ליזוזומים משניים - תוצרים של היתוך של ואקוולים אנדוציטיים עם ליזוזומים ראשוניים.
גופים שיוריים.
ייתכן שהפיצול של מקרומולקולות ביוגניות בתוך ליזוזומים לא ייגמר עד הסוף. במקרה זה, מוצרים לא מעוכלים מצטברים בחלל הליזוזום, והליזוזום המשני הופך לגוף שיורי (טלוליזום). התוכן שלהם דחוס ומסודר מחדש. לעתים קרובות בגופים הנותרים יש מבנה משני של שומנים לא מעוכלים, היוצרים מבנים מורכבים שכבות. שם גם מופקדים פיגמנטים.
אוטוליזוזומים.
לפי המורפולוגיה, אוטוליזוזומים (אוטופגוזומים) מסווגים כליזוזומים משניים, אך בתוך חלקיקים אלו ישנם שברים או אפילו מבנים ציטופלזמיים שלמים, כגון מיטוכונדריה, פלסטידים, ריבוזומים, יסודות רשת וכו'. ההנחה היא שתהליך האוטופגוציטוזיס הוא הקשורים לבחירה והרס של רכיבים סלולריים שהשתנו, "שבורים".
7. מנגנון גולגי.
מנגנון הגולגי (קומפלקס גולגי) על שם ק.גולגי, אורגנואיד תא.
בתאי בעלי חיים רבים, כגון תאי עצב, הוא לובשת צורה של רשת מורכבת הממוקמת סביב הגרעין. בתאי הצמחים והפרוטוזואה, מנגנון הגולגי מיוצג על ידי גופים בודדים דמויי מגל או מוטות. המבנה של אורגנואיד זה דומה בתאים של אורגניזמים צמחיים ובעלי חיים, למרות מגוון צורתו. (ראה נספח מס' 8).
הרכב מנגנון גולגי כולל: חללים מוגבלים על ידי ממברנות וממוקמים בקבוצות (5-10 כל אחד); בועות גדולות וקטנות הממוקמות בקצות החללים. כל האלמנטים הללו יוצרים קומפלקס אחד.
מנגנון גולגי מבצע פונקציות חשובות רבות.
מביא את החלבונים לצורת העבודה הסופית, למשל, "תפור" כמה חלבונים לחלבונים גדולים מתחמי חלבון, מחבר את יוני המתכת הדרושים לחלבונים מסוימים.
יוצר שלפוחיות ממברנות, אשר עוזבות את קומפלקס גולגי, או משחזרות את קרום התא או הופכות לליזוזומים. ליזוזומים הם אברוני ממברנה של התא, המייצגים שלפוחיות מיקרוסקופיות מלאות באנזימי עיכול.
הם מבצעים פונקציות עיכול והגנה. לדוגמה, בתאי הלבלב של יונקים, אנזימי עיכולהמצטברים בחללים של האורגנואיד.
הם יכולים להיצמד יחד עם ואקוול העיכול, לשפוך אליו אנזימי עיכול. במגע של תא עם חומר זר או עם תא זר, הליזוזומים נדבקים זה לזה קרום תאעל ידי שחרור האנזימים שלהם מחוץ לתא. אנזימים ליזוזומים יכולים גם לקחת חלק ב"מוות מתוכנת" של התאים שלהם.
הודות לפעילותו של מנגנון גולגי, מתרחשת חידוש וצמיחה של קרום הפלזמה.
8. ריבוזומים.
ריבוזומים נמצאים בתאים של כל האורגניזמים. אלו גופים מיקרוסקופיים בעלי צורה מעוגלת בקוטר של 15-20 ננומטר. כל ריבוזום מורכב משני חלקיקים בגודל לא שווים, קטנים וגדולים. (ראה נספח מס' 14).
תא אחד מכיל אלפים רבים של ריבוזומים, הם ממוקמים על הממברנות של הרשת האנדופלזמית הגרנורית, או שוכבים בחופשיות בציטופלזמה.
ריבוזומים מורכבים מחלבונים ו-RNA. תפקידם של הריבוזומים הוא סינתזת חלבון. סינתזת חלבון - תהליך קשה, אשר מתבצע לא על ידי ריבוזום אחד, אלא על ידי קבוצה שלמה, הכוללת עד כמה עשרות ריבוזומים משולבים. קבוצה זו של ריבוזומים נקראת פוליזום .
החלבונים המסונתזים מצטברים תחילה בתעלות ובחללים של הרשת האנדופלזמית ולאחר מכן מועברים לאברונים ולאתרי התא שם הם נצרכים.
רטיקולום אנדופלזמי וריבוזומים (ראה נספח מס' 8), הממוקם על הממברנות שלו, מייצגים מנגנון יחיד לביוסינתזה והובלה של חלבונים.
9. מיטוכונדריה.
אברונים מחולקים מעצמם. הציטופלזמה של רוב תאי החי והצומח מכילה גופים קטנים (0.2-7 מיקרון) - מיטוכונדריה (מיוונית "מיטוס" - חוט, "כונדריון" - דגן, גרגיר).
המיטוכונדריה נראים בבירור במיקרוסקופ אור, שבעזרתו ניתן לראות את צורתם, מיקומם, לספור את המספר (ראה נספח מס' 8).המבנה הפנימי של המיטוכונדריה נחקר באמצעות מיקרוסקופ אלקטרונים.
המעטפת של המיטוכונדריה מורכבת משני ממברנות - חיצונית ופנימית. הקרום החיצוני חלק, אינו יוצר קפלים וצמחים. הממברנה הפנימית, להיפך, יוצרת קפלים רבים המופנים אל חלל המיטוכונדריה. הקפלים של הממברנה הפנימית נקראים cristae (lat. "crista" - מסרק, תוצאה) מתרחש בהם תהליך החמצון של חומצת חלב, וכתוצאה מכך משתחררת אנרגיה, שנאגרת בצורה של ATP.
מספר הקריסטות אינו זהה במיטוכונדריה של תאים שונים. יכולים להיות בין כמה עשרות לכמה מאות, ובמיוחד יש הרבה קריסטלים במיטוכונדריה של תאים הפועלים באופן פעיל, למשל, תאי שריר.
המיטוכונדריה נקראות "תחנות כוח" של תאים, שכן תפקידן העיקרי הוא סינתזה של אדנוזין טריפוספט (ATP). חומצה זו מסונתזת במיטוכונדריה של התאים של כל האורגניזמים ומהווה מקור אוניברסלי לאנרגיה הנחוצה ליישום התהליכים החיוניים של התא ושל האורגניזם כולו.
מיטוכונדריה, כמו חיידקים, מתרבים בחלוקה ישירה.
10. מרכז תאים.
בתאים הסמוכים לגרעין יש אברון, הנקרא מרכז התא. החלק העיקרי של מרכז התא מורכב משני גופים קטנים (לפעמים יותר) - צנטריולים ממוקם באזור קטן של ציטופלזמה דחוסה. לכל צנטרול יש צורה של גליל באורך של עד 1 מיקרומטר, שקירותיו נוצרים על ידי מיקרוטובולים הממוקמים לאורך ציר הגליל. Centrioles מכילים חלבונים ו מספר גדול של RNA. לתא יש שני זוגות של צנטריולים. בכל זוג centrioles ממוקמים בניצב אחד לשני.
לצנטרולים תפקיד חשוב בחלוקת תאים. הם יוצרים חוטי חלבון ארוכים, מה שנקרא ציר הביקוע. במהלך חלוקת התא, הצנטריולים מתפצלים לכיוון הקטבים שלו, וקובעים את כיוון ציר החלוקה.
11. פלסטידים.
פלסטידים הם אברוני תאים מתחלקים עצמיים קרומיים. שלא כמו כל האברונים שנדונו לעיל, פלסטידים נמצאים רק בתאי צמחים.
במבנה, הם דומים למיטוכונדריה: הם נוצרים על ידי שני ממברנות, חלקים חיצוניים ופנימיים, ויוצרים שפעים שטוחים רבים - thylakoids . כל התילקואידים מוערמים כמו ערימות של מטבעות. כל מחסנית נקראת פנים . בין הגרנה נמצא נוזל פלסטידי פנימי הנקרא סטרומה . הוא מכיל DNA משלו, המבנה הדומה לזה של חיידקים.
פלסטידים מתרבים כמו חיידקים על ידי חלוקה ישירה.
על פי המאפיינים המבניים, נבדלים שלושה סוגים עיקריים של פלסטידים (ראה נספח מס' 15): ירוק - כלורופלסטים; אדום, כתום וצהוב - כרומופלסטים; חסר צבע - לויקופלסטים.
א) כלורופלסט.
אברונים אלו נמצאים בתאי העלים ואיברי צמחים ירוקים אחרים (נצרים, פירות לא בשלים), וכן במגוון אצות.
גודל הכלורופלסטים הוא 4-6 מיקרון, לרוב יש להם צורה אליפסה. בצמחים גבוהים יותר, בתא אחד יש בדרך כלל כמה עשרות כלורופלסטים.
צבע ירוקכלורופלסטים תלויים בתכולת הפיגמנט הכלורופיל שבהם.
כלורופלסט הוא האברון העיקרי של תאי הצמח שבו מתרחשת פוטוסינתזה, כלומר, יצירת חומרים אורגניים (פחמימות) מחומרים אנאורגניים (CO2 ו-H20) תוך שימוש באנרגיית אור השמש.
כלורופלסטים דומים מבחינה מבנית למיטוכונדריה. (ראה נספח מס' 16).הכלורופלסט תחום מהציטופלזמה על ידי שני ממברנות - חיצוניות ופנימיות. הקרום החיצוני חלק, ללא קפלים ויציאות, והפנימי יוצר שפעים מקופלים רבים המכוונים בתוך הכלורופלסט. לכן, מספר רב של ממברנות מרוכזים בתוך הכלורופלסט, ויוצרים מבנים מיוחדים - גרנה. מולקולות הכלורופיל ממוקמות בממברנות של הגראן, מכיוון שכאן מתרחשת הפוטוסינתזה.
ATP מסונתז גם בכלורופלסטים. בין הממברנות הפנימיות של הכלורופלסט מכיל DNA, RNA וריבוזומים. כתוצאה מכך, בכלורופלסטים, כמו גם במיטוכונדריה, יש סינתזה של החלבון הדרוש לפעילותם של האברונים הללו. כלורופלסטים מתרבים על ידי חלוקה (ראה נספח מס' 17).
ב) כרומופלסטים.
כרומופלסטים הם פלסטידים המכילים פיגמנטים דמויי שומן שצובעים את התא בצהוב, כתום ואדום. הם נמצאים בפירות בשלים של צמחים, המעניקים להם את הצבע המתאים, כמו גם בסתיו בעלים של עצים נשירים.
ג) לוקופלסטים.
Leucoplasts הם פלסטידים חסרי צבע. על הממברנה הפנימית, הם אינם מכילים פיגמנטים. בתא הם אחראים לסינתזה והצטברות של פוליסכרידים (עמילן). הם מצויים בכמות גדולה בתאי נצרים תת-קרקעיים (פקעות תפוחי אדמה, ארטישוק ירושלמי), וכן בפירות ובזרעים.
ד) טרנספורמציות פלסטיד.
כלורופלסטים יכולים בקלות להתדרדר לסוגים אחרים של פלסטידים. אנו רואים זאת עם הצהבה ואדום של פירות או עלים מבשילים בסתיו. בחושך, כלורופלסטים מסוגלים לשנות את צבעם, ולהפוך ללוקופלסטים. עם זאת, תהליכים אלה הם בלתי הפיכים: לויקופלסטים וכרומופלסטים לעולם אינם חוזרים לכלורופלסטים.
גרעין (גרעין תא) חלק חובהתאים בהרבה יצורים חד-תאיים ובכל רב-תאיים. גדלים מ-1 מיקרון (בחלק מהפרוטוזואה) ועד 1 מ"מ (בביצים של כמה דגים ודו-חיים). המונח "גרעין" (lat. Nucleus) שימש לראשונה על ידי ר' בראון בשנת 1833, כאשר תיאר את המבנים הכדוריים שצפה בתאי הצמח. (ראה נספח מס' 8).
א) מעטפת גרעינית.
החלל הפנימי של גרעין התא מופרד מהציטופלזמה על ידי קרום גרעיני המורכב משתי ממברנות. הממברנות של הממברנה הגרעינית דומות במבנהן לשאר מרכיבי הממברנה של התא ובנויות על פי אותו עיקרון: מדובר בסרטי ליפופרוטאינים דקים המורכבים משכבה כפולה של מולקולות שומנים, שבהן משובצות מולקולות חלבון.
המרווח בין הממברנה הגרעינית הפנימית והחיצונית נקרא perinuclear.
מספר רב של ריבוזומים ממוקמים בדרך כלל על פני הממברנה הגרעינית החיצונית, ולעיתים ניתן לצפות במעבר הישיר של ממברנה זו למערכת התעלות של הרשת האנדופלזמית הגרנורית של התא.
הממברנה הגרעינית הפנימית קשורה בשכבת חלבון סיבית דקה - למינה גרעינית מורכב מחלבוני למינ. רשת הפיברילים הצפופה של הלמינה הגרעינית מסוגלת להבטיח את שלמות הגרעין, גם לאחר פירוק קרומי הליפיד של מעטפת הגרעין בניסוי. עם בְּתוֹךלולאות של כרומטין הממלאות את הגרעין מחוברות ללמינה.
לממברנה הגרעינית יש חורים בקוטר של כ-90 ננומטר, הנוצרים מהתמזגות של הממברנה הגרעינית החיצונית והפנימית. חורים כאלה במעטפת הגרעינית מוקפים במבני חלבון מורכבים הנקראים קומפלקס הנקבוביות הגרעיניות.
שמונה תת-יחידות חלבון המרכיבות את הנקבובית הגרעינית ממוקמות סביב ניקוב המעטפת הגרעינית בצורת טבעות, בקוטר של כ-120 ננומטר, הנצפות במיקרוסקופ אלקטרונים משני צידי המעטפת הגרעינית. ליחידות המשנה של החלבון של קומפלקס הנקבוביות יש יציאות המכוונות למרכז הנקבוביות, שם נראה לפעמים "גרגיר מרכזי" בקוטר של 10-40 ננומטר.
גודל הנקבוביות הגרעיניות והמבנה שלהן הם סטנדרטיים עבור כל התאים האוקריוטיים. מספר הנקבוביות הגרעיניות תלוי בפעילות המטבולית של התאים: ככל שרמת התהליכים הסינתטיים בתא גבוהה יותר, כך יותר נקבוביות ליחידת שטח פנים של גרעין התא.
בתהליך של הובלה גרעינית-ציטופלסמית, נקבוביות גרעיניות מתפקדות כמעין מסננת מולקולרית, העוברות יונים ומולקולות קטנות (סוכרים, נוקלאוטידים, ATP וכו') באופן פסיבי, לאורך שיפוע הריכוז, ומבצעות הובלה סלקטיבית אקטיבית של מולקולות גדולות. של חלבונים וריבונוקלאופרוטאין, כלומר, חומצה ריבונוקלאית מחברת חומצות (RNA) עם חלבונים. כך, למשל, חלבונים המועברים לגרעין מהציטופלזמה, שם הם מסונתזים, חייבים להיות בעלי רצפים מסוימים של כ-50 חומצות אמינו, (מה שנקרא רצפי NLS), "המוכרים" על ידי קומפלקס הנקבוביות הגרעיניות. במקרה זה, קומפלקס הנקבוביות הגרעיניות, תוך שימוש באנרגיה בצורה של ATP, מעבירה באופן פעיל את החלבון מהציטופלזמה לגרעין.
ב) כרומטין.
גרעין התא הוא כלי הקיבול כמעט לכל המידע הגנטי של התא, ולכן התוכן העיקרי של גרעין התא הוא כרומטין: קומפלקס של חומצה דאוקסיריבונוקלאית (DNA) וחלבונים שונים.
בגרעין ובמיוחד בכרומוזומים מיטוטיים, ה-DNA הכרומטין מקופל שוב ושוב, ארוז בצורה מיוחדת להשגת מעלות גבוהותדחיסה. הרי את כל גדילי ה-DNA הארוכים, שאורכם הכולל, למשל, בבני אדם הוא כ-164 ס"מ, חייבים להיות ממוקמים בגרעין התא, שקוטרו מיקרומטרים בודדים בלבד. משימה זו נפתרת על ידי אריזה רציפה של DNA בכרומטין בעזרת חלבונים מיוחדים.
עיקר חלבוני הכרומטין הם חלבוני היסטון המהווים חלק מתת-יחידות כרומטין כדוריות הנקראות נוקלאוזומים . בסך הכל ישנם 5 סוגים של חלבוני היסטון.
הנוקלאוזום הוא חלקיק גלילי, המורכב מ-8 מולקולות היסטון, בקוטר של כ-10 ננומטר, שעליהן "נפצעים" קצת פחות משני סיבובים של מולקולת ה-DNA. IN מיקרוסקופ אלקטרוניכרומטין מפוקע מלאכותי כזה נראה כמו "חרוזים על חוט".
בגרעין החי, תאי נוקלאוזום מאוחדים זה עם זה בעזרת חלבון היסטון מקשר אחר, ויוצרים את מה שנקרא סיב כרומטין יסודי, בקוטר 30 ננומטר. חלבונים אחרים בעלי אופי שאינו היסטוני המרכיבים את הכרומטין מספקים דחיסה נוספת. כלומר, ערימה, כרומטין פיבריל, שמגיע שלו ערכים מקסימלייםבמהלך חלוקת התא בכרומוזומים מיטוטיים או מיוטיים.
בגרעין התא, כרומטין קיים הן בצורה של כרומטין מעובה צפוף, שבו סיבים יסודיים של 30 ננומטר ארוזים בצפיפות, והן בצורה של כרומטין מפוזר הומוגנית. היחס הכמותי בין שני סוגי הכרומטין הללו תלוי באופי הפעילות המטבולית של התא, במידת ההתמיינות שלו. לדוגמה, הגרעינים של אריתרוציטים של עופות, שבהם לא מתרחשים תהליכים פעילים של שכפול ותעתוק, מכילים למעשה רק כרומטין מעובה צפוף.
חלק מהכרומטין שומר על מצבו הדחוס והמעובה לאורך כל מחזור התא - כרומטין זה נקרא הטרוכרומטין ושונה מאוכרומטין במספר תכונות.
ג) שכפול ותעתוק.
תאים אוקריוטיים מכילים בדרך כלל כמה כרומוזומים (ממאתיים עד כמה מאות), המאבדים את צורתם הקומפקטית בגרעין (באינטרפאז, כלומר בין חלוקות מיטוטיות), משחררים וממלאים את נפח הגרעין בצורה של כרומטין. למרות המצב המעובה, כל כרומוזום תופס מיקום מוגדר בקפדנות בגרעין והוא מחובר למעטפת הגרעינית באמצעות למינה. קבוע בקפדנות על משטח פנימיקונכיות של הגרעין מבנים כאלה של כרומוזומים כמו צנטרומרים וטלומרים.
בשלב מסוים של מחזור החיים של התא, בתקופה הסינתטית, מתרחשת שכפול, כלומר, כל ה-DNA של הגרעין מוכפל, והכרומטין הופך להיות גדול פי שניים. החלבונים הדרושים לתהליך זה מגיעים כמובן מהציטופלזמה דרך הנקבוביות הגרעיניות. כך, התא מתכונן לחלוקת התא הקרובה - מיטוזה, מתי סה"כה-DNA בגרעין יחזור לרמתו המקורית.
יישום המידע הגנטי הכלול ב-DNA בצורת גנים מתחיל בתעתיק, כלומר בסינתזה של RNA שליח (i-RNA) - עותקים מדויקים של גנים, לפיהם יבנו לאחר מכן חלבונים בציטופלזמה על ריבוזומים.
תהליך זה מתרחש בנקודות שונות בנפח הגרעין, בלתי ניתנות להבחנה מורפולוגית מהכרומטין שמסביב. לרוב, ניתן לצפות בתעתיק מפוזר, כלומר. כרומטין מעובה.
בנוסף לכרומטין המרכיב את הכרומוזומים, גרעינים אוקריוטיים מכילים בדרך כלל נוקלאולי אחד או יותר.
ד) מטריצת חלבון גרעיני.
כדי לבצע תהליכי שכפול, שעתוק, וגם לשמור על מיקום מסוים של כרומוזומים בנפח הגרעין, ישנם מבני חלבון מסגרת הנקראים מטריצת החלבון הגרעיני. מטריצה כזו מורכבת משלושה מרכיבים מורפולוגיים לפחות:
שכבה סיבית היקפית - laminas;
מטריצה פנימית או interchromatin של הגרעין
והמטריצה של הגרעין.
תצפיות מראות שמרכיבי המטריצה הגרעינית אינם מבנים קפואים קשיחים, הם דינמיים ויכולים להשתנות מאוד בהתאם תכונות פונקציונליותגרעינים. הוכח כי למטריצת החלבון יש נקודות רבות של קישור חזק ל-DNA הליבה, אשר, בתורו, יש לו רצפי נוקלאוטידים מיוחדים הדרושים לכך.
13. נוקלאולי.
א) מספר הגרעינים בתא.
החל באצות ירוקות, פטריות ופרוטוזואה תחתונות וכלה באצות אורגניזמים גבוהים יותר, לכל התאים יש מבנים תוך גרעיניים חובה - נוקלאולי (ראה נספח מס' 18).לכלל זה יש מספר רב של חריגים, שרק מדגישים את החשיבות והנחיצות של הגרעין ב מעגל החייםתאים. חריגים כאלה כוללים תאים של ביציות מתפצלות, שבהם הנוקלאולי נעדרים בשלבים המוקדמים של העובר, או תאים שהשלימו התפתחות והתמחו באופן בלתי הפיך, למשל, כמה תאי דם.
מספר הגרעינים בתא יכול להשתנות, אך מספרם לגרעין תלוי באיזון הגנים של התא. נמצא כי היווצרות נוקלאולי מעורבת מקומות מסויימיםכמה כרומוזומים, שניתן לאתר היטב את הקשר שלהם עם הגרעין בטלופאז ובפרופאזה. לכרומוזומים כאלה, ככלל, יש התכווצויות משניות, שהאזורים שבהם הם מקומות שבהם מתרחשת התפתחות הנוקלאולי. McClintock (1934) קרא לקטעים אלה של כרומוזומים "מארגנים גרעיניים".
אתרי ההיצרות המשניות אופייניים במיוחד למיקומם של מארגנים גרעיניים, אך לעיתים ניתן למקם את האחרונים בקצוות הכרומוזומים או במספר מקומות לאורכו של הכרומוזום.
מספר כוללנוקלאולי לגרעין נקבע על פי מספר המארגנים הגרעיניים ועולה בהתאם לפלואידיה של הגרעין. עם זאת, לעתים קרובות מספר הגרעינים לגרעין הוא פחות ממספרמארגנים גרעיניים. הוכח שנוקלאולי יכולים להתמזג; בנוסף, כמה מארגנים משתתפים לפעמים ביצירת גרעין אחד.
אפילו בעבודותיו של M. S. Navashin (1934) הוכח כי הלוקוס הכרומוזומלי, אשר ב תנאים רגיליםיוצר גרעין גדול, הופך ללא פעיל כאשר לאחר הכלאה, לוקוס "חזק" יותר מופיע בגרעין על כרומוזום אחר. העובדה שבתנאים מסוימים ניתן לדכא את פעילותם של מארגנים גרעיניים מסוימים או להגביר את פעילותם של אחרים שהיו בעבר במצב סמוי ונסתר, מצביעה על כך שנשמר איזון מסוים בכמות החומר הגרעיני בתאים. , או, במילים אחרות, התוצרים ה"ברוטו" המיוצרים על ידי הנוקלאולי.
בהתבסס על העובדות לעיל, ניתן להסיק את המסקנות הבאות:
Þ היווצרות הגרעינים ומספרם קשורים לפעילות של חלקים מסוימים של כרומוזומים - מארגנים גרעיניים, הממוקמים לרובבאזורים של היצרות משניות;
Þ שינויים במספר הגרעינים בתאים מהסוג הזהיכול להתרחש עקב איחוי נוקלאולי או עקב שינויים באיזון הכרומוזומלי של התא.
ב) פיזיולוגיה וכימיה של הגרעין.
הגרעין, בהשוואה למרכיבים אחרים של התא, מאופיין כמבנה הצפוף ביותר עם הריכוז הגבוה ביותר של RNA, עם פעילות גבוההעבור סינתזת RNA.
ריכוז ה-RNA בנוקלאולי תמיד גבוה מריכוז ה-RNA ברכיבים אחרים של התא, כך שריכוז ה-RNA בגרעין יכול להיות גבוה פי 2-8 מאשר בגרעין, ופי 1-3 מאשר בגרעין. הציטופלזמה. היחס בין ריכוז ה-RNA בגרעין, הגרעין והציטופלזמה של תאי כבד עכברים הוא 1:7, 3:4, 1, בתאי הלבלב - 1:9, 6:6, 6.
לא נמצא DNA בגרעין, אך עם זאת, כאשר בוחנים תאים קבועים, תמיד מובחן אזור של כרומטין סביב הגרעין. הכרומטין הפרי-נוקלאולרי הזה, לפי מיקרוסקופ אלקטרונים, נראה כחלק בלתי נפרד מ מבנה מורכבגרעין.
הגרעין הוא אחד האתרים הפעילים ביותר בתא לשילוב מבשרים ב-RNA. ה-RNA הגרעיני הוא המבשר של ה-RNA הציטופלזמי.
RNA ציטופלזמי מסונתז בגרעין.
ג) RNA גרעיני.
הערכה תוכן כלליבשברים הגרעיניים של חלבונים, RNA ו-DNA, ניתן לראות ש-RNA מהווה כ-10% מהמסה הכוללת של הגרעין.
מכיוון שעיקר ה-RNA הציטופלזמי הוא RNA ריבוזמלי, אנו יכולים לומר ש-RNA גרעיני שייך למעמד זה.
הרעיון שהגרעין הוא האתר של סינתזת rRNA ויצירת ריבוזומים אושר על ידי העובדה שחלקיקי RNP בודדו מתכשירים גרעיניים, אשר, הן בהרכב ה-RNA (על פי תכונות השקיעה) והן בגודלם, ניתן לאפיין כ ריבוזומים או מבשרייהם.עם מקדמי שיקוע שונים.
ד) DNA גרעיני.
מחקרים ביוכימיים מצאו כמות מסוימת של DNA בנוקלאולים המבודדים, אותה ניתן לזהות עם כרומטין פרינוקלאולרי או עם מארגני כרומוזומים גרעיניים. תכולת ה-DNA בנוקלאולי המבודדים הוא 5-12% מהמשקל היבש ו-6-17% מסך ה-DNA של הגרעין.
ה-DNA של המארגן הגרעיני הוא אותו DNA שעליו מתרחשת סינתזה גרעינית, כלומר. ריבוזומלי, RNA.
לפיכך, ממחקרים ביוכימיים, עלו רעיונות שגנים זהים רבים לסינתזת rRNA היו מקומיים על DNA בגרעין. סינתזה של rRNA ממשיכה דרך היווצרות של מבשר ענק והפיכה נוספת שלו (התבגרות) למולקולות RNA קצרות יותר עבור תת-יחידות ריבוזומים גדולות וקטנות.
בחקר הגרעינים של ביציות ניוט, החוקרים נתקלו בתופעה מעניינת - שפע העל של נוקלאולי. במהלך צמיחת הביצית, מופיעים עד 1000 נוקלאולים קטנים שאינם קשורים לכרומוזומים. את הגרעינים האלה זיהה או. מילר, יחד עם זה, כמות ה-rDNA לכל גרעין ביצית עולה. תופעה זו קיבלה שם הַגבָּרָה . זה טמון בעובדה שמתרחשת שכפול יתר של אזור המארגן הגרעיני, עותקים רבים יוצאים מהכרומוזומים והופכים לנוקלאולים עובדים נוספים. תהליך כזה הכרחי להצטברות של מספר עצום (10 12) של ריבוזומים לתא ביצית, מה שיבטיח התפתחות עתידית של העובר על שלבים מוקדמיםאפילו בהיעדר סינתזה של ריבוזומים חדשים. הגרעינים העל-מספריים נעלמים לאחר הבשלת תא הביצית.
ה) מבנה אולטרה של הנוקלאולי.
כשלומדים מספר גדול תאים שוניםבבעלי חיים וצמחים, מצוין מבנה סיבי או רשתי של הגרעינים, סגור במסה מפוזרת צפופה פחות או יותר. הוצעו שמות לחלקים אלה: חלק סיבי - נוקלאונמה וחלק מפוזר, הומוגני - חומר אמורפי, או חלק אמורפי . מחקרים מיקרוסקופיים של אלקטרונים שנעשו כמעט במקביל לכך חשפו גם את המבנה הסיבי-חוטי של הנוקלאולי.
עם זאת, מבנה חוטי כזה של הגרעין לא תמיד בא לידי ביטוי בבירור. בתאים מסוימים, גדילים בודדים של נוקלאונמים מתמזגים, והנוקלאולי עשויים להיות הומוגניים לחלוטין.
בבדיקה מדוקדקת יותר של הגרעין, ניתן לראות שהעיקרי רכיבים מבניים nucleolus - גרגירים צפופים בקוטר של כ-15 ננומטר וסיבים דקים בעובי 4-8 ננומטר. במקרים רבים (ביציות של דגים ודו-חיים, תאי צמחים מריסטמטיים), הרכיב הפיברילרי מורכב לאזור מרכזי (ליבה) צפוף נטול גרגירים, והגרגירים תופסים את האזור ההיקפי של הגרעין. במספר מקרים (לדוגמה, תאים של שורשי צמחים), לא נצפה מבנה נוסף באזור הגרגירי הזה.
נמצא שהאזורים האמורפיים של הנוקלאולי הם הטרוגניים. במבנה שלהם מתגלים אזורים בעלי צבע נמוך - מרכזי פיברילרים - ואזורים כהים יותר המקיפים אותם, שגם להם יש מבנה פיברילרי.
בנוסף לשני המרכיבים הללו של הגרעין ב לָאַחֲרוֹנָהתשומת לב רבה הוקדשה למבנה של הכרומטין הפרינוקלאולרי. הכרומטין הזה והרשת התוך-גרעינית של DNA הם מערכת מאוחדתומהווים מרכיב אינטגרלי מהגרעין.
גרגירים וחלק פיבריל מורכבים מריבונוקלאופרוטאין.
הוכח שמרכזי הפיבריל האור הם המכילים rDNA.