Filogenija, filogenija - istorinė raida gyvų organizmų pasaulis, tiek bendras, tiek atskiras taksonomines grupes: karalystes, tipus (skyrius), klases, būrius (ordus), šeimas, gentis, rūšis. Filogenijos terminą įvedė E. Haeckelis.
Šiuolaikinė gyvūnų sistema, sukurta remiantis giminystės principais, gali būti pavaizduota kaip genealoginis (arba filogenetinis) medis, atspindintis pagrindinius gyvūnų evoliucijos etapus. Filogenezė įvairios grupės organizmai tirti netolygiai. Geriausiai ištirta stuburinių (ypač aukštesnių grupių), bestuburių – moliuskų, dygiaodžių, nariuotakojų filogenezija. Prokariotų filogenija yra menkai suprantama.
Evoliucinio medžio pagrindu gyvūnai yra vienaląsčiai. Vienaląsčiuose organizmuose visas kūno funkcijas atlieka viena ląstelė. Yra keletas hipotezių apie kilmę vienaląsčiai eukariotai s: 1) eukariotai simbiogenezės būdu išsivystė iš prokariotų. Šią hipotezę patvirtina faktas, kad eukariotinės mitochondrijos ir chloroplastai turi savo DNR; 2) eukariotai atsirado dėl prokariotinių ląstelių diferenciacijos į citoplazmą ir organelius. Visų tipų šiuolaikiniai pirmuonys yra kilę iš senovės sarkoflagellatų.
Daugialąsčiai, labiau organizuoti gyvūnai, kuriuose individualios funkcijos organizmus atlieka atskiros ląstelės arba organai. Visi daugialąsčiai organizmai individualiame vystymesi prasideda nuo vienos ląstelės – zigotos ir pradiniai etapai embrionas (blastula) primena vienaląstę koloniją.
Yra keletas hipotezių, susijusių su daugialąsčių gyvūnų kilme. Plačiausiai žinomos E. Haeckel ir I. I. hipotezės. Mechnikovas. Pagal Haeckel teoriją, visi daugialąsčiai gyvūnai kilę iš vienos protėvių formos – gastrea. I.I. Mechnikovas manė, kad daugialąsčių gyvūnų protėvis buvo vėliavėlių kolonija. Pirmas daugialąstelis organizmas buvo sferinės formos, buvo padengtas ląstelių sluoksniu su žiuželiais ir kūno viduje turėjo virškinimo ameboidines ląsteles, laisvai plūduriuojančias vandenyje. I.I. Mechnikovas tai pavadino fagocitele. Tolesnė daugialąsčių organizmų evoliucija vyko keturiomis kryptimis. Viena iš evoliucijos krypčių paskatino lamelinio tipo susidarymą, susidedantį iš vieno ląstelių sluoksnio ir vedančio šliaužiojantį gyvenimo būdą. Kita evoliucijos kryptis paskatino susiformuoti Sponge Type su dviem ląstelių sluoksniais, nesusiformavusiais į audinį. Jie dar neturi nervų sistemos. Tai nejudrūs bentoso gyvūnai. Trečioji evoliucijos kryptis paskatino Koelenteratų tipo susidarymą. Koelenteratai jau yra tikri dvisluoksniai (ektoderma ir endoderma) gyvūnai, turintys pirminį žarnyną, burną, radialinę simetriją, difuzinę nervų sistemą. Tarp jų yra laisvai plaukiančių ir prisirišusią gyvenimo būdą vedančių gyvūnų. Ketvirtoji evoliucijos kryptis paskatino dar labiau organizuotų trisluoksnių gyvūnų formavimąsi. Šių gyvūnų audiniai ir organai buvo suformuoti iš trijų ląstelių sluoksnių (gemalo sluoksnių) ektodermos, endodermos ir mezodermos.
Pirmieji trisluoksniai buvo gyvūnai, panašūs į žarnyno planarijas. Trijų sluoksnių gyvūnų evoliucija vyko trimis kryptimis. Viena kryptis vedė į formavimąsi plokščiųjų kirmėlių, kurioje nėra kūno ertmės, o parenchima yra tarp organų. Plokščiosios kirmėlės yra pirmieji dvišaliai gyvūnai (visi paskesni tipai turi tokią simetriją). Jie pirmiausia pasirodo raumenų sistema(lygus), nervų ganglijos. Tačiau susidaręs žarnynas yra aklinai uždaras, neturi išangės.
Kita kryptis paskatino pirminių ertmių gyvūnų susidarymą apvaliosios kirmėlės. Jų žarnos tampa nuo galo iki galo, susideda iš trijų skyrių su išangė. Toliau vyksta koncentracija nervų ląstelės, suformuojant perifaringinį nervinį žiedą. Plokščiosioms kirmėlėms ir anelidams būdingas odos-raumenų maišelis, virškinimo, šalinimo ir reprodukcinės sistemos.
Trečioji trijų sluoksnių evoliucijos kryptis veda į deuteruotų gyvūnų formavimąsi. Antrinės kūno ertmės susidarymas suteikė didesnį mobilumą ir stabilumą vidinė aplinka organizmas. Antrinės ertmės yra labiau organizuotos: jos turi kraujotakos sistemą. Tai apima visus kitus bestuburius ir chordatus.
Deuterostomų evoliucija paskatino protostomų ir deuterostomų susidarymą. Protostomų evoliucija. Moliuskai ir pirminiai anelidai kilę iš protostomų protėvio. Savo ruožtu šiuolaikiniai anelidai ir nariuotakojai atsirado iš pirminių anelidų.
Annelidai turi progresyvias organizacines ypatybes: antrinę kūno ertmę, metameriškai išpjaustytą kūną, uždarą kraujotakos sistemą (kituose bestuburiuose neuždarą), perifaringinį žiedą ir nervų grandinę.
Nariuotakojai pasiekia didesnį tobulumą: širdis kyla kraujotakos sistemoje; nervų sistemoje didėja ganglijų koncentracija; susiformavo egzoskeletas(chitino danga); būdinga kvėpavimo organų (žiaunų, trachėjos, plaučių) įvairovė; formuojasi sujungtos galūnės ir sparnai. Tobulesnė nariuotakojų struktūra leido jiems plačiai apgyvendinti Žemę.
Moliuskai turi antrinę kūno ertmę, kompleksą Vidaus organai(susidaro inkstai, trijų kamerų širdis, žiaunos, plaučiai), dauguma turi kalkingą apvalkalą, saugančią kūną. Tačiau moliuskų kraujotakos sistema nėra uždara, o nervų sistema yra išsibarsčiusio mazginio tipo.
Deuterostomos evoliucija. Dygiaodžiai ir chordatai kilę iš deuterostomų protėvių.
Dygiaodžiai. Jiems būdinga: antrinė kūno ertmė, kraujotakos sistemoje susidaro tarpai, savotiška vandens-motorinė (ambulakralinė) sistema, radialinė simetrija, nervų sistema žiedo su radialiniais spinduliais pavidalu.
Pirminiai chordatai tikriausiai išsivystė iš deuterostominių kirminų protėvių. Akordų izoliacija yra susijusi su mobilumo padidėjimu. Filogenetinius chordatų ryšius su bestuburiais liudija: 1) audiniai ir organai susidaro iš trijų gemalo sluoksnių (pirmą kartą susiformuoja plokščiųjų kirmėlių atveju); 2) dvišalė simetrija (atsiranda plokščiųjų kirmėlių); 3) antrinė kūno ertmė (atsiranda anneliduose); 4) antrinė burna (susijusi su dygiaodžiais); 5) uždara kraujotakos sistema (annelidai);
Iš pirminių chordatų atsirado šiuolaikiniai nekranialiniai (lancetiniai) ir stuburiniai gyvūnai. Lervų chordatai yra šoninė chordatų šaka, kurios dauguma pereina į sėslų gyvenimo ciklą, todėl chordatų požymių turi tik lervos stadijoje.
Cefalothordatai yra primityvūs chordatai. Jie turi nefridiją, panašią į anelidų metanefridiją, uždarą kraujotakos sistemą be širdies, kaip ir anelidai.
Stuburinių gyvūnų protėviai atsirado maždaug prieš 500 milijonų metų. Jų evoliucija vyko dviem kryptimis: be žandikaulio (ciklostomos) ir žandikaulio (visi kiti stuburiniai gyvūnai).
Pirmoji žandikaulė – šarvuotos žuvys išsivystė iš šarvuotų bežandikaulių. Jie jau buvo stiprūs vidinis skeletas, smegenų ir visceralinė kaukolė, žandikauliai su dantimis, suporuoti pelekai. Šios žuvys išnyko. Juos pakeitė kremzliniai ir kaulinė žuvis kurio kūnas buvo padengtas žvynais.
Kremzlinės žuvys buvo paplitusios jūrose ir pagal maisto rūšį buvo plėšrūnės.
Kaulinės žuvys iš pradžių pasirodė gėlame vandenyje. Jie sukūrė operkulu (apsaugo žiaunas nuo užsikimšimo) ir plaukimo pūslę (palengvina plaukimą gėlame vandenyje). Džiūvančiuose vandens telkiniuose (paleozojaus) atsirado skiltinių žuvų (plaučių ir skilčių žuvų), kuriose, be žiaunų, išsivystė plaučių maišeliai oro kvėpavimui, o ant pelekų susiformavę raumenys leido atsiremti. ant kieto pagrindo. Vėliau kaulinės žuvys gyveno gėluose ir jūriniuose vandens telkiniuose. Šiuo metu kaulinės žuvys yra viena iš labiausiai klestinčių stuburinių grupių (20 tūkst. rūšių).
Devone atsirado varliagyviai. Varliagyviai kilę iš senovės skiltinių žuvų. Pirmieji varliagyviai buvo Ichthyostegidae (keturpėdės žuvys). Jos, kaip ir žuvys, turėjo uodegos peleką, žiaunų dangą, žvynus. Tačiau skirtingai nei žuvys, jie turėjo dvi poras penkių pirštų galūnių ir plaučius. Jie daugiausia gyveno vandenyje.
Iš ichtiostego atkeliavo tikri varliagyviai – stegocefalai (kiaukščiagalviai). Stegocefalijų galva buvo padengta odos kaulų apvalkalu. Stegocefalai gyveno iki kreidos periodo pradžios. Tikriausiai iš jų atsirado šiuolaikiniai bešarvus varliagyviai ir pirmieji ropliai.
Nusileidimą lydėjo aromorfozė: žiauninis kvėpavimas pasikeitė į plaučių kvėpavimą, o pelekai – penkių pirštų galūnes, širdis tapo trijų kamerų, atsirado antrasis kraujotakos ratas, pakito jutimo organai, tobulėjo nervų sistema.
Pirmieji ropliai pasirodė karbone. Fosilijos Seimourijos liekanos turėjo varliagyvių ir roplių bruožų. Tikėtina, kad iš tokių formų atsirado pirmieji ropliai – kotilozaurai. Jie taip pat turėjo kaukolę kaulo dėžutės pavidalu (panašią į stegocefalus) su skylutėmis akims, šnervėms ir parietaliniam organui. Roplių klestėjimas buvo mezozojuje.
Pasitaiko: epidermio keratinizacija ir žvynelių susidarymas, arba skruostai (apsaugo organizmą nuo išsausėjimo), plaučių, širdies struktūros komplikacija, skeleto pagerėjimas, serozinių gemalo membranų susidarymas. ir amniono (suteikia galimybę daugintis sausumoje).
Ropliai sukūrė dvi naujas stuburinių gyvūnų klases: žinduolius ir paukščius.
Paukščiai kilo iš pirmųjų paukščių iš driežų dinozaurų grupės užpakalinės kojos. Iš senųjų pirmųjų paukščių buvo aptiktos ilgauodegės archeopterikso liekanos. Archeopteriksas turėjo roplių (ilga uodega, dantys ant žandikaulių, nagai ant sparnų) ir paukščių (plunksnos, sparnai, išvaizda) požymių. Tikri paukščiai atsirado nuo pirmųjų paukščių mezozojaus pabaigoje. Jie taip pat turėjo mažus dantis.
Progresuojantys paukščių bruožai: šiltakraujiškumas, keturių kamerų širdis, vienas aortos lankas (sukuria visišką sisteminės ir plaučių kraujotakos atskyrimą).
Žinduoliai pasirodė anksčiau nei paukščiai. Spėjama, kad jie kilę iš gyvulinių dantytų driežų, kurie turėjo nemažai progresuojančių požymių: dantys išsidėstę alveolėse, antrinis kaulo gomurys, galūnės buvo po kūnu, o ne šonuose, buvo pakitimų. skelete. Cynodonts buvo arčiausiai žinduolių tarp žinduolių dantytų roplių. Tolesnė žinduolių evoliucija siejama su dantų sistemos pasikeitimu. Dėl šių pokyčių susiformavo dvi gyvūnų grupės. Vienos grupės palikuonys - multituberkulioziniai buvo maži gyvūnai, turėjo daug gumbų ant krūminių dantų, stipriai išsivystę smilkiniai ir neturėjo ilčių. Gali būti, kad ši grupė sukėlė monotremus.
Kita grupė vystėsi progresyviau. Tai buvo maži gyvūnai, valgantys vabzdžius, smulkūs gyvūnai. Ateityje ši grupė skirstoma į dvi nepriklausomas šakas – žemutinę, marsupialinę ir aukštesnę, placentinę.
Marsupials atsirado kreidos periodu. Seniausia marsupialų grupė yra possumų šeima.
Kreidos periodu atsirado ir placentos žinduoliai. Seniausia placentų grupe laikomas vabzdžiaėdžių būrys. Manoma, kad jie sukėlė daugumą vėlesnių placentos grupių. Gali būti, kad primatai kilę tiesiai iš vabzdžiaėdžių.
Progresyvi Stlano žinduolių evoliucija įgyjant svarbiausias adaptacijas: šiltakraujiškumą, greitą medžiagų apykaitą, placentos formavimąsi, jauniklių maitinimą pienu, palikuonių priežiūrą. Tai palengvino progresuojantys kvėpavimo, kraujotakos, nervų sistemos. Žinduoliai turi keturių kamerų širdį, vieną aortos lanką, arterinę ir veninio kraujo, alveolinės struktūros plaučiai, gerai išvystytas antrinis nervinis skliautas – neopalija.
13… Prokariotinės ląstelės sandara. Bakterijų augimas, vystymasis, dauginimasis. Maisto bakterijų rūšys. Fotosintezė, chemosintezė, fotoredukcija. Mikroorganizmų vertė gamtoje ir šalies ūkyje prokariotinės ląstelės apima bakterijas ir melsvadumblius. At prokariotai branduolinė medžiaga neturi branduolinės membranos, jų plazminėje membranoje yra angliavandenių ir aminorūgščių. Prokariotai turi tik vieną chromosomą, kurią sudaro tik DNR. Ši chromosoma turi žiedo formą ir joje nėra šarminių baltymų – histonų. Prokariotų chromosomų rinkinys yra haploidinis.Šie vienaląsčiai organizmai neturi mitozės ir neturi mitozinio aparato. Prokariotai neturi mejozės, o lytinio proceso metu perduodama tik dalis paveldimos informacijos, o pats organizmas yra gameta. Prokariotų citoplazminė DNR yra lokalizuota plazmidėse, o citoplazminių membranų nėra. Prokariotų ląstelių sienelėse yra peptidoglikanų. Prokariotuose mitochondrijų nepastebima, kvėpavimo fermentai lokalizuoti mezosomose, kurios yra vidiniame plazminės membranos paviršiuje.Fotosintezės procesai vyksta naudojant bakteriochlorofilo pigmentą, o sieros junginiai tarnauja kaip reduktoriai. Prokariotinės ribosomos yra mažesnės nei eukariotinės ribosomos. Citoplazma nejuda, vakuolės joje yra labai retos. Prokariotų žvyneliai yra submikroskopinio dydžio, žvyneliuose yra vienas ar keli mikrovamzdeliai (fibrilės) Prokariotai jautrūs antibiotikams, atsparūs rentgeno ir pakilusi temperatūra, atlaiko kaitinimą iki + 75 ... + 90 ° С. būdingas bruožas prokariotai yra tai, kad jie negali egzistuoti kartu su kitų tipų ląstelėmis. bakterijos- vienaląsčiai organizmai; juos galima pamatyti tik pro mikroskopą. Todėl jie vadinami mikrobais arba mikroorganizmais. Bakterijos įvaldė pačias įvairiausias buveines: gyvena dirvoje, dulkėse, vandenyje, ore, ant išorinių gyvūnų ir augalų dangalų bei kūno viduje. Jų galima rasti net karštuose šaltiniuose, kur jie gyvena maždaug 60 °C ar aukštesnėje temperatūroje. Mikroorganizmų veikla yra svarbą visoms kitoms gyvoms būtybėms, nes bakterijos ir grybai ardo organines medžiagas ir dalyvauja medžiagų cikle gamtoje. Be to, bakterijos įgyja viską didesnę vertęžmonių gyvenime, ir ne todėl, kad sukelia įvairias ligas, o todėl, kad iš jų galima gauti daug reikalingų produktų.
LĄSTELĖS STRUKTŪRA. Prokariotai: nesusiformavęs branduolys, mitochondrijos, plastidai. Citoplazmos centre yra nukleoidas, kuriame yra viena "chromosoma" su apskrita DNR molekule; nėra atskirtas nuo citoplazmos membrana. Ląstelės viduje membranos ataugos atlieka EPS, Golgi aparato, funkcijas; mezosoma dalyvauja kvėpavime; yra ribosomos. Citoplazmą dengia išorinė membrana ir tanki ląstelės sienelė. Kartais ląstelė panardinama į pusiau skystą koloidinę kapsulę. Pagal ląstelės sienelės sandarą bakterijas galima suskirstyti į dvi grupes.Vienos yra gramneigiamos, todėl vadinamos gramteigiamos, o kitos nuplaunant dažus tampa bespalvės ir todėl vadinamos gramneigiamomis. Augimas ir dauginimasis.Individualus ląstelių augimas ir nelytinis dauginimasis. Bakterijų ląstelių paviršiaus ir tūrio santykis yra labai didelis. Tai prisideda greitas perėmimas maistinių medžiagų iš aplinką per difuziją ir aktyvų transportą. IN palankiomis sąlygomis bakterijos auga labai greitai. Augimas pirmiausia priklauso apie terpės temperatūrą ir pH, maistinių medžiagų prieinamumą ir jonų koncentraciją. Privalomiesiems aerobams taip pat reikia deguonies, o privalomiems anaerobams, priešingai, jo visiškai nėra. Pasiekusios tam tikrą dydį, bakterijos pereina į nelytinis dauginimasis(dvejetainis dalijimasis), t.y., jie pradeda dalytis susidarant dviem dukterinėms ląstelėms. Perėjimą prie dalijimosi lemia branduolio tūrio ir citoplazmos tūrio santykis. Prieš ląstelių dalijimąsi vyksta DNR replikacija, kurios metu mezosomos išlaiko genomą tam tikroje padėtyje.Mezosomos taip pat gali prisitvirtinti prie naujų pertvarų tarp dukterinių ląstelių ir kažkaip dalyvauti ląstelės sienelės medžiagų sintezėje. Sparčiausiai augančios bakterijos dalijasi kas 20 minučių.
4 bakterijų mitybos rūšys ir kai kurios jų savybės.
Yra žinomos trys pagrindinės grupės fotosintetinis(bakterijos, kurios energiją gauna iš saulės): 1. Žaliosios sieros bakterijos . Anaerobinės bakterijos, kurios naudoja vandenilio sulfidą (H2S) ir kitus redukuotus sieros junginius kaip vandenilio ir elektronų donorus.
Pavyzdys: šviesa
2H2S + CO2 →→→→→→→→ [CH2O] + 2S + H2O.
bakteriochlorofilas
Šiuo atveju siera nusėda.
2. Purpurinės sieros bakterijos. Ląstelės yra tamsiai raudonos spalvos, nes bakteriochlorofilo yra daug mažiau nei raudonųjų ir rudųjų pigmentų. Tai dažniausiai anaerobai; fotosintezė juose vyksta taip pat, kaip ir ankstesnėje grupėje.
3. Violetinės nesierinės bakterijos . Šioms bakterijoms įvairūs organiniai junginiai tarnauja kaip vandenilio šaltinis, kuris naudojamas CO2 (fotoautotrofuose) arba kai kurių organinių junginių (fotoheterotrofų) redukavimui.
Chemosintetinis organizmai (chemoautotrofai) – tai bakterijos, kurios CO2 naudoja kaip anglies šaltinį, tačiau energiją gauna ne iš saulės, o padedamos cheminės reakcijos. Energija gali išsiskirti, tarkime, oksiduojant vandenilį, sieros vandenilį, sierą, geležį (II), amoniaką, nitritus ir kitus neorganinius junginius. Geležies bakterijose : Deguonis
Fe2+ →→→→→Fe3+ + Energija;
Bespalvėse sieros bakterijose : Deguonis arba
S→→→→→→→ SO42-+ Energija;
Sieros sulfatas
Anaerobinėmis sąlygomis kai kurios rūšys naudoja nitratą kaip vandenilio akceptorių ir taip vykdo denitrifikaciją.
Nitrifikuojančiose bakterijose: deguonis
2NH4+→→→→→ NO-2 + Energija
Amoniako nitritas
Visos ką tik aptartos bakterijos yra aerobinės, nes deguonis yra elektronų (ir vandenilio) akceptorius.
Chemosintetinės bakterijos atlieka labai svarbų vaidmenį biosferoje; jie daugiausia dalyvauja azoto cikle ir taip palaiko dirvožemio derlingumą.
Fotoredukcija – žalioms ir violetinėms serobakterijoms., kurios kaip šaltinį naudoja ne H2O, o H2S
GAMTOS IR ŽMONIŲ EKONOMINĖS VEIKLOS REIKŠMĖ.
pieno fermentacijos bakterijos(heterotrofiniai saprotrofai), kurie skaido angliavandenius: Teigiamas: pieno rūgšties produktų (varškės, rūgpienio, sviesto, grietinės) ruošimas, pašarų silosavimas, raugintų kopūstų, agurkų ir pomidorų rauginimas. Neigiamas: produktų sugadinimas.
Acto fermentacijos bakterijos(heterotrofiniai saprotrofai), kurie skaido angliavandenius: Teigiamas: alkoholio oksidacija iki acto rūgštis, kuris naudojamas vaisiams ir daržovėms marinuoti, konservuoti. Neigiamas: produktų sugadinimas.
puvimo bakterijos(heterotrofiniai saprotrofai), kurie skaido baltymus: Teigiamas: sanitarinis vaidmuo – organinių likučių mineralizacija. Neigiamas: maisto gedimas; Siekiant išvengti puvimo, naudojamas džiovinimas, sūdymas, marinavimas, sterilizavimas, pasterizavimas ir cukravimas.
Mazgelių bakterijos(heterotrofiniai simbiotai): Rhizobium mazginės bakterijos prasiskverbia pro ankštinių augalų (dobilų, lubinai, liucernos ir kt.) šaknis ir su jais į simbiozę. Dėl to ant šaknų formuojasi navikai – mazgeliai, užpildyti bakterijomis, kurios iš atmosferos azoto sintetina azoto junginius, prieinamus tiek augalui šeimininkui, tiek kitiems augalams. Į šį natūralų dirvožemio praturtinimą azoto trąšomis atsižvelgiama rengiant lauko sėjomainas, kuriose būtinai yra ankštinių augalų.
14. Eukariotinės ląstelės sandara. trijų sluoksnių membranaRibosomosGolgi kompleksasMitochondrijosLizosomosChloroplastaiChromoplastaiLeukoplastaiLąstelių centras arba centrosomaInkliuzaiVakuolėscitoskeletasKarioplazmabranduolys1. 2 3 4 5
15.Ontogenija ir jos periodai. ankstyva embriogenezė. Tiesioginė ir netiesioginė plėtra.(51)
Ontogenezė- individualus organizmo vystymasis nuo gimimo iki mirties. Jis tęsis skirtingoms organizacijoms nėra tas pats ir gali trukti nuo kelių mindos šimtus metų (vėžlys). Ontogeniškumas yra glaudžiai susijęs su filogenezija, bet nėra absoliutus filogenijos pasikartojimas. Aplinkos įtakoje jis keičiasi ir gali atsirasti mutacijų kintamumas. ontogeniškumo periodai:
1 ikiembrioninis
2. gemalinė (embrioninė)
3. poembrioninis
1 periodu lytinių ląstelių vystymasis ir paruošimas tolesniam vystymuisi po apvaisinimo (spermatogenezė ir ovogenezė) spermatogenezė iš 4 periodų: 1. reprodukcija; 2. augimas; 3. nokinimas; 4. formavimas
Spermatogenezė: Spermatogonija 2n "1 eilės spermatocitai 2n" spermatocitai 2 poros 2n "spermatozoidai n.
Ovogenezė: O vogonia 2n "oocitai 1 pora2n" oocitai 2 pora2n "1 kiaušinis + 3 kryptiniai kūnai (sumažinti)
2 periode embrionas vystėsi gimdoje (motinos įsčiose) arba išorinėje aplinkoje. „Har-Xia“ greiti formavimo procesai trumpalaikis sukelti daugybinių org-ma atsiradimą. Turi 3 laikotarpius: 1. pradinis- 1 savaitė embriono vystymasis prasideda nuo apvaisinimo momento ir baigiasi embriono implantavimu į motinos gleivinę. 2 gemalai. Laikas nuo pradžios 2 iki 8 savaičių nuo implantacijos iki organogenezės pabaigos. 3 vaisiaus- vaisiaus vystymasis nuo 9 savaičių iki gimimo. Padidėjęs augimas, formavimas, hematotrofinė mityba.
3-asis poebr periodas prasideda nuo gimimo momento, t.y. organizmo išlaisvinimas iš lukštų užuomazgų. Šiuo vystymosi laikotarpiu atsirado specifinis anatominis ir funkcinis bruožas, vadinamas amžiaus. Periodizavimas:
1-10 dienų - naujagimis
11d - 1 metų kūdikis
1-11 metų vaikai (mergaitės ir berniukai)
11-15m (d) ir 12-16 (m) - paauglys
16-20 (d) ir 17-21 (m) - jaunimas
21-55 (w) ir 22-60 (m) - brandus
56, 61-74 - senyvo amžiaus
75-90 senatvės, per 90 šimtamečių
Tiesioginis vystymasis (ne lervos tipo), susijęs su > trynio skaičiumi kiaušinyje arba org. Motinos organizme būdinga žuvims, žinduoliams, paukščiams, ropliams.
Netiesioginis vystymasis – (lerva) su metamorfoze – charakteris. d / nenuoseklumas, vėžiagyviai, vabzdžiai, kirminai, charakteris. Trumpas embrioninis laikotarpis katės metu. Lerva susidaro. Save, bet egzistuoti. Ne pilna transformacija – kiaušinėlis-lerva-imago. Visiškas virsmas – kiaušinis – lerva – lėliukė – imago. Ankstyvoji embriogenezė: spermatozoidas patenka į kiaušinėlio ląstelę, citoplazma ištirpsta - sinkarijos pulkas (branduoliai susilieja "diploidinio branduolio" zigota 2n-div. Ant blastomerų-4, 8,16,32,64,128 ..-arr. blastula (embrionas su ertme viduje) - invaginacijos būdu - dvisluoksnis - gastrula, ect.ent) - (blastocoel, blastopore) - išsivystė. 3 gemalas. Listok. endoderma, mezoderma, ektoderma
Smulkinimas gali būti vienodas ir nelygus (diskoidinis)
Gastruliacija vyksta 4 būdais:
1 invaginacija (invaginacija)
2 epiboliai (užteršimas)
3 delaminacija (sluoksniacija)
4 imigracija
Nervinio vamzdelio, stygos, vaisiaus vandenų ir kamieno raukšlių neuruliacijos angažas
ektoderma formuoja nervinę plokštelę-st. neurula (nervinio vamzdelio klojimas, stygos), klausa, uoslė, išorinis apvalkalas.Endoderma – žarnyno, kepenų, plaučių, kasos liaukos pradžia. Mezodermos forma. Raumenys (dermotomas, miotomas), raumeninis audinys, kraujotakos sistema, visi kremzliniai ir kauliniai skeleto elementai, paryškinimas, lytinė sistema.
16.pagrindiniai filogenezės keliai. Divergencija, konvergencija, paralelizmas.(52)
Evoliucinis vystymasis gali vykti Skirtingi keliai priklausomai nuo objekto ir jo santykio su aplinka. Yra keletas filogenezės formų.
Filetinė evoliucija – tai laipsniškas evoliucinis tam tikros taksonominės grupės persitvarkymas, dėl kurio atsiranda nauja grupė, kuri skiriasi nuo pradinės. Filetinės evoliucijos pavyzdžiu gali būti keletas arklių protėvių: eogippus – epigippus – merik-hippus – gashogippus – šiuolaikinis arklys.
Divergencija - pagrindinis evoliucijos kelias, kuri buvo įtvirtinta Darvino teorijos pagrindu. Divergentinę evoliuciją lemia taksono išsiskyrimas į kelias grupes, veikiamas elementarių evoliucijos ir prisitaikymo prie jos veiksnių. skirtingos sąlygos egzistavimas. Esant divergencijai, panašumą lemia bendra kilmė, o skirtumus – prisitaikymas prie kitokios aplinkos. Divergencijos pradžia rūšies viduje formuojantis ekotipams, ekologinėms rasėms, geografiniams porūšiams. Kadangi charakteriai skiriasi, veislės tampa rūšimis, rūšys toliau diferencijuojasi ir formuoja gentis, gentys tampa šeimomis ir pan.
Skirtumas gali būti kelių formų: dichotominis išsišakojimas (rūšies suskaidymas į dvi vaikų rūšis), radiacija (rojaus kritimas, rūšies į kelias vaikų rūšis), atsiskyrimas (kartu su dukterinėmis įmonėmis egzistuoja protėvių darinys, suformuotas iš izoliuotų skirtingas laikas populiacijos). Divergencija vystosi homologinių organų pagrindu. Klasikinis pavyzdys Divergencija gali būti vadinama Galapagų archipelago kikilių charakterių skirtumais.
Divergencijos biologinė reikšmė slypi tame, kad ji lemia gyvų organizmų įvairovę, vienodą rūšių pasiskirstymą ekologinėse nišose ir išlygina kovos už būvį įtampą.
Konvergencija yra priešingas divergencijos procesas. Tai reiškia, kad tomis pačiomis sąlygomis savarankiškas panašių simbolių įgijimas formomis, kurios iš esmės nėra susijusios panašiais organais ar struktūromis. Pavyzdžiui, kurmio meškos rausvos galūnės panašios; banginių šeimos ir žuvys turi tą pačią kūno formą. Konvergencija lemia bendrą „kulkšnių“ bradančių paukščių (gervių ir garnių) panašumą. Konvergencijos reiškiniai apima „geografines“ (klimatines) „paraleles“, kurios išreiškiamos didesniu šaltųjų šalių šiltakraujų gyvūnų skaičiumi, palyginti su artimiausiais vidutinio klimato regionų atstovais, didesniu paukščių sankabų skaičiumi ir žinduolių išmatų skaičiumi. Arktyje dėl padidėjusios tropinių paukščių pigmentacijos.
Tolimose gyvūnų grupėse tą konvergenciją lemia išorinės sąlygos giminaičiai taip pat turi atsižvelgti į giminystę. Šiuo atžvilgiu klasėje pastebima konvergencija.
Lygiagretumas.Jei anksčiau skirtingos grupės patenka į skirtingas egzistavimo sąlygas, jos gali savarankiškai, lygiagrečiai suformuoti panašius prisitaikymo bruožus. Pavyzdžiui, giliavandenėse žuvyse liuminescencijos organai atsirado lygiagrečiai ir nepriklausomai; įvairių kačių kardadantiškumo išsivystymas Kartais paralelizmas apima visas organų sistemas arba visos organizacijos dalį. Pavyzdžiui, lygiagretus varliagyvių dvikvėpių žuvų vystymasis. Tikėtina, kad dvikvėpių ir varliagyvių panašumus nulemia šių grupių santykiai. Ypač reikšmingi yra širdies struktūros paraleliai ir kraujotakos sistema. Kuo labiau susiję ryšiai tarp organizmų, tuo dažniau lygiagretus vystymasis apima reikšmingas visos organizacijos dalis. Tai liudija ir paleontologiniai duomenys, pavyzdžiui, proboscių vystymasis. Kuo organizmai arčiau vienas kito, tuo panašesnės jų reakcijos į tą patį aplinkos pasikeitimą, tuo panašesni (lygiagretesni) yra jų struktūros pokyčiai. Lygiagrečiam vystymuisi svarbi ir išorinė aplinka, ir vidiniai veiksniai (t.y. istoriškai susiklosčiusios paveldimos organizmo struktūros). Santykis vidinių veiksnių su išorine lemia organizmų evoliuciją. Pavyzdžiui, skirtingose klimato zonose gyvenantys kanopiniai gyvūnai bėga nuo plėšrūnų. Todėl kanopiniams gyvūnams pastebimas itin didelio kojų pirštų laipsnio išsivystymas ir laipsniškas kraštinių pirštų mažėjimas.
Genetiniai lygiagrečios evoliucijos pagrindai. Pasikeitus aplinkos veiksniams, panašiai kinta ir giminingi organizmai. Kovoje už būvį per natūralią atranką šie pokyčiai įgauna „paveldimą“ pavidalą. Atrankos medžiaga yra individualūs struktūros nukrypimai, ty mutacijos. Giminingos ar artimos formos, kaip taisyklė, suteikia tas pačias mutacijas (pavyzdžiui, skirtingų tipų Drosophila). Tačiau tai nereiškia, kad lygiagrečios mutacijos yra pagrįstos tais pačiais tų pačių genų pokyčiais.
Yra žinoma, kad skirtingi to paties organizmo genai sukelia panašias mutacijas. Ypač jei šie pokyčiai yra nedideli. Ir kadangi evoliucijos eigoje atrenkami tik mažų mutacijų deriniai, lygiagrečiai pokyčiai gali būti kuriami remiantis skirtingu genetiniu pagrindu.
KURSINIS DARBAS
biologijoje
„Eukariotų kilmė
Užbaigė: Severova Vera
10 klasė, mokykla N55
Patikrintas: Strukova N.A.
Riazanė 1999 m
1 . Simbiotinė teorija:
1.1 Pagrindinis skirtumas tarp prokariotų ir eukariotų;
1.2 Prokariotiniai organizmai;
1.3 Eukariotinės ląstelės;
1.4 Mitochondrijos ir plastidai;
1.5 eukariotų gebėjimas fotosintezei;
2. Gyvybės pirmtakai;
3. Fermentuojantys organizmai:
3.1 Ankstyvųjų ląstelių energijos šaltiniai;
3.2 Metanogenezė kaip energijos šaltinis;
3.3 Melsvabakterijos ir chloroksibakterijos;
4. Fotosintezė ir oras;
5. Asociacijos ir eukariotai:
5.1 nuoseklių simbiozių teorijos;
5.2 Simbiotinis mobilumo įgijimas;
5.3 Eukariotai – fotosintezės;
6. Žvilgsnis į ląstelių evoliuciją:
6.1 Tiesioginės giminystės hipotezė;
6.2 Hipotezė: nuo cianobakterijų iki dumblių;
6.3 2 hipotezių suderinamumas: tiesioginė giminystė ir
endosimbiozė;
7. Išvados;
8. Literatūra.
ENDOSIMBIOZĖS TEORIJA
Visi gyvi organizmai Žemėje paprastai skirstomi į prokariotus ir eukariotus. Pagrindinis bruožas prokariotai, skirtingai nei eukariotai, jiems trūksta pilno ląstelės branduolys padengtas membrana. Prokariotinės ląstelės yra labai mažo dydžio, maždaug 1 µm. Apimtis eukariotinės ląstelės turintis pilnavertį branduolį, 800–1000 kartų didesnį už prokariotinių ląstelių tūrį. Šiuo atžvilgiu ląstelėms – eukariotams gyvybinėms funkcijoms atlikti reikalingas daug didesnis membraninis aparatas. Iš tiesų, eukariotų ląstelėje buvo nustatytos tarpląstelinės struktūros, apsuptos membranų, nepriklausomų nuo plazmalemos ir viena kitos. Šios struktūros vadinamos organelėmis. Kiekviena organelė: mitochondrijos, chloroplastas, branduolys, lizosomos, Golgi aparatas ir kt. turi savo specifines funkcijas ląstelės gyvenime.
Molekulinių-biologinių metodų pagalba buvo gauta įtikinamų įrodymų, kad visos gyvos būtybės turėtų būti suskirstytos į 3 karalystes: bakterijas, archajas ir eukariotus. Bakterijos ir archėjos Žemėje atsirado iš bendro protėvio – vadinamojo progenoto – maždaug prieš 4 milijardus metų, o paleontologų teigimu, eukariotai Žemėje atsirado praėjus 500 milijonų metų po prokariotų. Populiariausia ir pakankamai pagrįsta eukariotinių ląstelių kilmės idėja šiuo metu yra įvairių prokariotų daugialypės endosimbiozės idėja. Pasak žinomų mikrobiologų A. Kluyverio ir K. Van Nielio, būtent iš senovės prokariotų buvo atrenkamos ekonomiškiausios ir dažnai polifunkcinės biocheminės reakcijos pasižyminčios ląstelės, kurios tapo pagrindu. svarbiausi būdai medžiagų apykaitą.
Simbiotinė ląstelių atsiradimo ir evoliucijos teorija remiasi dviem naujomis biologijoje sampratomis.Pagal pirmąją iš šių sąvokų esminis skirtumas gyvojoje gamtoje yra skirtumas tarp prokariotų ir eukariotų, t.y. tarp bakterijų ir organizmų, susidedančių iš ląstelių, turinčių tikrus branduolius – protistų, gyvūnų, grybų ir augalų. Antroji koncepcija yra ta, kad kai kurios eukariotinių ląstelių dalys atsirado dėl simbiozių evoliucijos, t.y. nuolatinių asociacijų tarp organizmų susidarymas skirtingi tipai. Daroma prielaida, kad trys organelių klasės – mitochondrijos, blakstienos ir fotosintetinės plastidės – atsirado iš laisvai gyvenančių bakterijų, kurios dėl simbiozės pateko į tam tikrą seką prokariotinių šeimininkų ląstelėse. Ši teorija labai remiasi neodarvinizmo idėjomis, kurias sukūrė genetikai, ekologai, citologai ir kiti mokslininkai, kurie susiejo Mendelio genetiką su Darvino idėja. natūrali atranka Joje taip pat remiamasi visiškai naujomis arba neseniai atgimusiomis mokslo sritimis: molekuline biologija, ypač baltymų struktūra ir aminorūgščių seka, mikropaleontologija, tiriančia ankstyviausius gyvybės pėdsakus Žemėje, ir net fizikos bei chemijos pagrindu. atmosferą, nes šie mokslai yra susiję su biologinės kilmės dujomis. Visi ląsteliniai organizmai gali būti suskirstyti į penkias karalystes: prokariotų karalystę (Monera, kuri apima bakterijas) ir keturias eukariotų karalystes (Protoctista, Animalia, Fungi, Plantae). Protokistai yra eukariotiniai organizmai, kurie nėra gyvūnai, grybai ar augalai. Protokistų karalystei priklauso dumbliai, pirmuonys, gleivių pelėsiai ir kiti neaiškios kilmės eukariotiniai organizmai. Protistai labiau apibrėžiami kaip vienaląsčiai eukariotai. Taigi, karalystė Protoctista apima ne tik protistus – vienaląsčius eukariotus, bet ir artimiausius daugialąsčius jų palikuonis, tokius kaip raudonieji ir rudieji dumbliai ir taip pat daug mikroorganizmų, panašių į grybus, pavyzdžiui, chitridai. Remiantis tradiciniu tiesioginės filiacijos požiūriu, ląstelių organelės, tokios kaip mitochondrijos ir plastidai, atsirado suskaidžius pačią ląstelę.
FORMUOJA KELI PROKARIOTAI
VIENAS EUKARIOTAS
Ląstelės yra membranomis surištos sistemos, mažiausios iš biologinių vienetų, galinčių savarankiškai daugintis. Net mažiausiose ląstelėse yra genų DNR molekulių pavidalu (viena ar daugiau) ir baltymų sintezės aparatas, susidedantis iš kelių tipų RNR ir daugybės baltymų. Visose ląstelėse yra ribosomų, maždaug 0,02 µm skersmens kūnų, sudarytų iš mažiausiai trijų tipų RNR ir apie penkiasdešimt skirtingų baltymų. Prokariotiniai organizmai yra mažiausi biologiniai vienetai, atitinkantys šį ląstelės apibrėžimą. Jie neturi branduolių. Kita vertus, pavienės eukariotinės (turinčios branduolį) ląstelės tam tikra prasme nėra vienetai. Tai įvairių baltymus sintezuojančių vienetų (nukleocitoplazmos, mitochondrijų ir plastidų) kompleksai, kurių tipas ir skaičius skiriasi priklausomai nuo rūšies. Taigi, remiantis šia teorija, eukariotinės ląstelės atsirado bendradarbiaujant iš pradžių nepriklausomiems elementams, susijungusiems tam tikra tvarka. Taylor šią idėją apie atskirą eukariotinės ląstelės dalių kilmę ir vystymąsi bei vėlesnį jų suvienijimą pavadino nuoseklių endosimbiozių teorija. Sąlygos aukštos temperatūros o rūgštingumas paskatino nukleocitoplazmoje gamintis specialios klasės baltymai – histonai – apsaugoti jos DNR. Mitochondrijos sugebėjo suskaidyti trijų anglies junginius į anglies dioksidą ir vandenį. Simbiotinė nukleocitoplazmos ir mitochondrijų asociacija pradėjo keistis abipusės priklausomybės stiprėjimo kryptimi. Šis kompleksas apėmė motorines organeles - blakstienas ir žiuželes. Po šios transformacijos sekė morfogenetinė transformacija, kurią sudarė ryški ląstelių asimetrija. Integravus tarpusavyje priklausomas nukleocitoplazmas, mitochondrijas, undulipodijas, buvo baigta sukurti eukariotinės ląstelės poligenominė struktūra. Pirmoji eukariotinė ląstelė buvo heterotrofinė. Ji valgė jau paruoštus organinius junginius. Šiuose eukariotuose atsirado mitozės procesai, o vėliau - mejozė. Ir galiausiai, eukariotai įgijo gebėjimą fotosintezei tapti pagrindine eukariotų organizacija, o po jos užbaigimo - paskutiniu šio epo įvykiu. Simbiozės vis labiau integravosi. Šiuo metu kiekvienos organelės priklausomybė nuo kitų organelių medžiagų apykaitos produktų tapo tokia visiška, kad tik šiuolaikiniai metodai analizė leidžia atsekti pirminių partnerių medžiagų apykaitos kelius. Eukariotų struktūra yra įvairi, tačiau metaboliškai jie išlieka vienodi. Buvo sukurti eukariotų medžiagų apykaitos gudrybės įvairios bakterijos dar prieš tą laiką, kai jie susivienijo, virsdami šeimininku ir jo organelėmis. Prokariotai pasiekė didelę energijos gavimo būdo ir biosintezės sudėtingumo įvairovę. Jie paliko savo egzistavimo pėdsaką gerokai anksčiau, nei iš eukariotinės ląstelės susiformavo tokie dideli organizmai kaip gyvūnai su skeletu.
Schema pro Ir eukariotinių ląstelių konvergencija simbiozės būdu
GYVYBĖS PREKURSAI
Šiandieninių organizmų protėviai atsirado daugiau nei prieš tris milijardus metų iš organinių junginių, susidariusių Žemės paviršiuje. Šios koncepcijos patrauklumą iš dalies lemia jos patikrinamumas.
S. Milleris ir H. Urey parodė, kad tokiomis atmosferos sąlygomis, kurios vyko jaunoje Žemėje, gali susidaryti aminorūgštys ir kitos organinės molekulės. Įvairių tipų organinės molekulės susiformavo savaime per pirmuosius milijardus žemės gyvavimo metų. Daug organinių molekulių buvo rasta tarpžvaigždinėje erdvėje ir meteorituose. Reiškia, organiniai junginiai atsiranda šviesos elementų sąveikos metu, esant energijos šaltiniams. Universalumas cheminis mechanizmas bioreprodukcija reiškia, kad visa gyvybė Žemėje yra kilusi iš bendrų protėvių: visa dabartinė gyvybė kilo iš ląstelių su informacinėmis sistemomis, pagrįstomis DNR replikacija ir baltymų sinteze, kuriai vadovauja pasiuntinio RNR.
RŪGINGI ORGANIZMAI
Gali būti, kad nukleorūgštys nuo pat pradžių buvo atskirtos nuo aplinkos lipoproteininėmis membranomis. Informaciją pradėjo koduoti nukleotidų seka DNR molekulėse. Seniausios ląstelės kaip tiesioginius energijos šaltinius naudojo ATP ir aminorūgštis, kurios yra baltymų dalis. konstrukciniai komponentai. Taigi - seniausia gyvybė egzistavo heterotrofinių bakterijų pavidalu, kurios maistą ir energiją gaudavo iš abiotinės kilmės organinių medžiagų. Neodarvinistinei organinei evoliucijai reikalingi trys reiškiniai: dauginimasis, mutacija ir selektyvus aplinkos spaudimas. Dauginimasis yra nepritaikoma biologinės evoliucijos sąlyga. Pakankamai tiksliai dauginantis, mutacijos ir natūrali atranka yra neišvengiamos. Paaiškinti tokio dauginimosi kilmę reiškia suprasti pačios gyvybės kilmę. Mutacija, pagrindinis evoliucijos kintamumo šaltinis, sukelia paveldimus pokyčius savaime besidauginančioje sistemoje. Sukūrus patikimą reprodukciją, prasidėjo neodarvinizmo evoliucija. Organizmai, kurie išsivystė sintetiniu keliu reikalingas narvas komponentų, turėjo pranašumą prieš kitus. Seniausi autotrofai, tai yra organizmai, galintys patenkinti savo energijos ir energijos poreikius organinės medžiagos ah dėl neorganinių šaltinių buvo anaerobinių metaną gaminančių bakterijų. Fotosintezės gebėjimas pirmiausia buvo sukurtas deguoniui jautriose bakterijose. Sugeriantis saulės šviesa bakterinio chlorofilo pagalba jie atmosferos anglies dioksidą pavertė organinėmis medžiagomis.Anaerobinės fotosintezės atsiradimas negrįžtamai pakeitė Žemės paviršių ir atmosferą.
Anglies dioksidas buvo pašalintas iš atmosferos ir paverčiamas organine medžiaga pagal bendrąją fotosintezės lygtį: 2H2X+CO2=CH2O+2X+H2O. Fotosintetinės bakterijos, kurios naudojo vandenilio sulfidą CO2 nusėdusios elementinės sieros mažinimui. Iš jų atsirado bakterijų, galinčių fotosintezuoti, išskirdamos deguonį. Šio tipo fotosintezė tikriausiai atsirado dėl fotosintetinėse sieros bakterijose susikaupusių mutacijų. Šis kelias paskatino melsvadumblių arba melsvadumblių atsiradimą. Neseniai atrastas ir ištirtas nauja grupė fotosintetiniai prokariotai, gaminantys O2 – chloroksibakterijas. Galima daryti prielaidą, kad dumblių ir augalų ląstelėse randamų plastidų protėviai buvo ciano ir chloroksibakterijos. O2 susikaupus dėl mikrobų fotosintezės, žemės atmosfera oksidavosi, o tai sukėlė visos gyvybės krizę – iki tol pasaulyje gyveno anaerobinės bakterijos. Norėdami išgyventi, mikrobai turėjo vengti deguonies, slėpdamiesi pelkėse, arba jie turėjo sukurti medžiagų apykaitą, galinčią apsaugoti juos nuo didėjančio deguonies kiekio.
FOTOSINTEZĖ IR ORO
Maždaug prieš 2 milijardus metų vienas po kito vyko įvairūs evoliuciniai procesai, sukuriantys medžiagų apykaitos įvairovę. Gebėjimas gaminti specifinius fermentus gali būti perduodamas toje pačioje rūšyje naudojant į virusą panašias daleles, vadinamas plazmidėmis. Prokariotų biosintetiniai gebėjimai stebina: iš anglies dioksido, kelių druskų ir redukuotų neorganinių junginių, kurie tarnauja kaip energijos šaltiniai, chemoautotrofiniai mikroorganizmai gali susintetinti deguonies turinčiame neapšviestame vandenyje visas sudėtingiausias augimui ir dauginimuisi reikalingas makromolekules. Šie biocheminiai procesai paskatino elementų cirkuliacijos sistemas net tada, kai nebuvo augalų, grybų, gyvūnų.
asociacijos ir eukariotai.
Žvilgsnis į ląstelės evoliuciją.
Tiesioginis filialas.
Tradicinis požiūris į eukariotinių ląstelių kilmę ir evoliuciją yra toks, kad visi gyvi organizmai yra tiesiogiai kilę iš vienos protėvių populiacijos dėl natūralios atrankos įtakoje susikaupusių mutacijų.
Taškinės mutacijos, dubliacijos, delecijos ir kiti paveldimi pokyčiai, kurie, kaip žinoma, turėjo įtakos gyvūnų ir augalų evoliucijai, taip pat yra atsakingi už eukariotinių ląstelių diferenciaciją nuo prokariotinių – tiesioginės giminės teorija. y., nesimbiotinės ląstelių kilmės samprata. Galbūt žalieji dumbliai yra visų augalų protėvių palikuonys. Daugelis šių dumblių savybių rodo, kad jie ir augalai turi bendrą protėvį. Gyvūnų filogenija yra sudėtingesnė, tačiau tiesioginės giminystės sąvoka tikriausiai taikoma ir jų evoliucijos keliams. Kempinės, matyt, išsivystė iš protistų, nepriklausomai nuo kitų gyvūnų grupių. Taigi, bent jau gyvūnams ir augalams, hipotezė, kad sudėtingi eukariotiniai organizmai išsivystė iš paprasčiau išsidėsčiusių eukariotų, susikaupus individualioms mutacijoms, veikiant natūraliai atrankai, yra gana pagrįsta. Evoliucinės naujovės remiasi labai tobulų palankių genų išsaugojimu. Taigi, nauji organizmai, turintys didesnį gebėjimą kontroliuoti savo aplinką, labiau išsivystė išlaikant labai pritaikytų genų dubliavimąsi ir rekombinaciją, o ne įgydami naujų savybių per paprastas, atsitiktines taškines mutacijas. Tiesioginės giminės hipotezė nepaaiškina eukariotų kilmės iš prokariotų. Pavyzdžiui, pertrauka tarp melsvadumblių, negalinčių susirgti mitoze, ir tokių formų, kurių mitozė yra visiškai išsivysčiusi, kaip raudonieji dumbliai, yra sunkiai suprantamas tiesioginės filiacijos požiūriu, tačiau tai natūraliai išplaukia iš simbiotinės teorijos.
Remiantis tiesioginės giminės teorija ir simbiozės teorija, visi organizmai Žemėje kilę iš biochemiškai panašių bakterijų protėvių, o iš eukariotinių mikrobų atsirado augalai ir gyvūnai. Dėl fotosintetinių bakterijų atsirado dumbliai, o galiausiai ir augalai, o kai kurie dumbliai prarado plastidus ir tapo grybų ir gyvūnų protėviais. Tiesioginės giminystės teoretikai sutinka, kad plastidai ir kitos eukariotinės organelės, įskaitant branduolį, susidarė diferencijuojant pačiose ląstelėse, ir vadina tai botaniniu mitu.
Augalų rūšių filogenezė
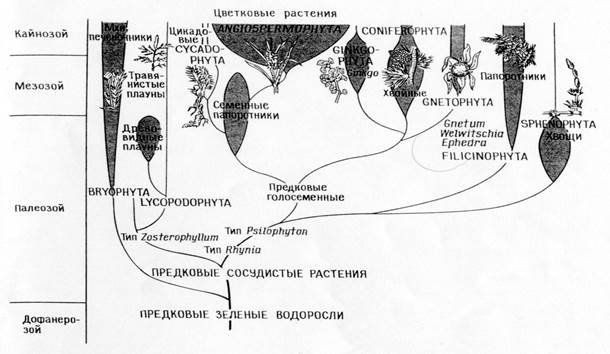
Filogenijų, atitinkančių tiesioginės giminės teoriją, palyginimas (A)
ir simbiozės teorija (B)
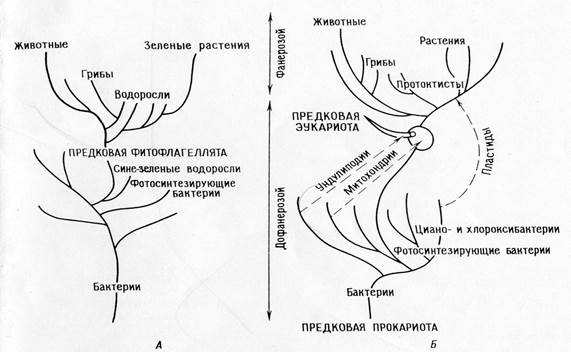
Skirtumai tarp tiesioginės giminės teorijos ir simbiotinės teorijos:
Tiesioginės giminės teorija.
Pagrindinė dichotomija: gyvūnai – augalai
Fotosintetiniai eukariotai (dumbliai ir žalieji augalai) išsivystė iš fotosintetinių prokariotų (mėlynadumblių)
Augalai ir jų fotosintezės sistemos išsivystė monofiletiškai Prekambrijoje.
Gyvūnai ir grybai išsivystė iš autotrofų, kurie prarado plastidus.
Mitochondrijos tarpląsteliniu būdu diferencijuojasi fotosintetiniuose dumbliuose.
Dešiniosiose piktžolėse undolipodijos, mitozinis aparatas ir visi kiti eukariotų organeliai diferencijuojasi tarpląsteliniu būdu.
Visi organizmai kilę iš savo protėvių – prokariotų, susikaupę pavienių mutacijų.
PASEKMĖ: Pagal teoriją turi būti dešiniosios piktžolės; sunku susieti šią teoriją su iškastiniais įrašais ir sukurti atitinkamą filogeniją.
Viena po kitos einančių endosimbiozių teorija:
Pagrindinė dichotomija: prokariotai – eukariotai.
Fotosintetiniai eukariotai (branduoliniai dumbliai ir augalai) ir nefotosintetiniai eukariotai (gyvūnai, grybai, pirmuonys) išsivystė iš bendrų heterotrofinių protėvių (ameboflagelatų).
Fotosintezės evoliucija Archėjuje buvo monofilinė bakterijose (įskaitant ciano ir chloroksibakterijas), fotosintetiniai protistai staiga atsirado, kai jų heterotrofiniai protistai-protėviai įsitraukė į simbiozę su fotosintetiniais prokariotais (protoplastidais).
Gyvūnai ir dauguma dumblių bei grybų yra tiesiogiai kilę iš heterotrofinių protistų.
Mitochondrijos jau buvo heterotrofiniuose protistuose, kai pastarieji dėl simbiozės įgijo plastides. Promitochondrijos anksčiau buvo įgytos simbiotiniu būdu.
Mitozė išsivystė heterotrofiniams eukariotų protistams diferencijuojant undolipodinę sistemą.
Prokariotai išsivystė iš ankstyvųjų prokariotų protėvių per keletą pavienių mutacijų įvykių, eukariotai taip pat išsivystė iš eukariotų protėvių per pavienius mutacijos įvykius, o eukariotų protėviai išsivystė iš prokariotų per daugybę simbiozių.
Pasekmės: teorija paaiškina biocheminius kelius ir jų perteklius organelėse. Tai leidžia sukurti nuoseklią filogeniją, paaiškinančią „biologinį atotrūkį“ tarp proterozojaus ir fanerozojaus laikų. Teorija numato, kad organelės turi išlaikyti replikacijos sistemų likučius.
botanikos mitas
Nuo Jussieu laikų žemesni augalai vadinami talofitais.o aukštesnieji – bryofitai ir tracheofitai. Remiantis botanikos mitais, bendri fotosintetinių bakterijų ir melsvadumblių protėviai dėl tiesioginės giminės sukėlė eukariotinių dumblių, įskaitant jūrinius, pradžią. Organizmų grupės, kurių filogeniją labiausiai veikia tiesioginės giminės sąvoka, atsidūrė botanikos srityje – tai bakterijos, dumbliai, grybai ir augalai.
Hipotetiniai organizmai, galintys būti jungtimi tarp melsvadumblių ir žaliųjų dumblių, buvo vadinami protėvių fitomonadais (protėvių fitomonadais) arba dešiniosiomis piktžolėmis. Kai kurie autoriai teigė, kad Cyanidium ir Cyanophora eukariotai, kurių pigmentų sistemos yra panašios į cianobakterijas, gali būti tinkami kandidatai į dešiniųjų piktžolių vaidmenį. Talofituose nerasta tarpinių formų tarp nemitozinės melsvadumblių būklės ir visiškai išsivysčiusios tipinės žaliųjų dumblių ir augalų mitozės. Nors grybai ir žydintys augalai visiškai neturi undulipodijų, jiems būdinga mitozė ir mejozė, kurioms reikalingi verpstės, susidedančios iš mikrovamzdelių. Visuotinai sutariama, kad visi daugialąsčiai organizmai, turintys mikrovamzdelius, yra kilę iš eukariotinių mikroorganizmų su undolipodijomis, tačiau niekas nežino, iš kurios protistų grupės. Botanikos mitą sustiprino fotosintezės senumas ir sudėtingumas, taip pat tai, kad daugelis organizmų (bakterijų, euglenoidų, parazitinių augalų) tiek gamtoje, tiek laboratorijoje dėl mutacijų prarado fotosintezę. Kadangi fotosintezė yra anaerobinis procesas, galiausiai užtikrinantis visų organizmų egzistavimą, ji turėjo išsivystyti labai anksti mūsų planetos gyvybės istorijoje, dar prieš atsirandant mitozei, kuriai reikalingas deguonis. fotosintezėje dalyvauja daug pigmentų, lipidų ir fermentų, kurie tvarkingai išsidėstę labai sudėtingose membranose. Tuo pačiu metu dumblių ir augalų fotosintezės mechanizmų detalės, viena vertus, ir cianobakterijos, iš kitos pusės, yra stebėtinai panašios. Visi šie organizmai iš vandens išskiria deguonies dujas, kurios yra vandenilio atomų, naudojamų CO2 redukavimui į ląstelės organines medžiagas, šaltinis. Daugelis dumblių ir pirmuonių neturi nei mitozės, nei genetinės sistemos, panašios į gyvūnų ir augalų. Blakstienos turi specialią genetinę sistemą su 2 tipų branduoliais, o ląstelės dažnai būna daugiabranduolės. Vieno tipo branduoliai yra makrobranduoliai, o kiti – daug mažesni mikrobranduoliai. Mikrobranduoliai yra fiziologiškai nereikšmingi. tačiau jie saugo genų kopijas ir lytinio proceso metu patiria mejozę. Fiziologiškai esminiai makrobranduoliai yra RNR transkripcijos vieta ir taip valdo ląstelių fiziologiją. Vienas iš sunkumų išaiškinant ryšį tarp melsvadumblių ir eukariotinių dumblių yra duomenų apie undolipodijų kilmę trūkumas: tarp cianobakterijų, kurios niekada neturi undolipodijų, ir daugelio dumblių, kurios turi, nerasta. Kita problema yra visiškas nebuvimas lytinis procesas melsvadumbėse: tarp nelytinių melsvadumblių ir dumblių, kurių daugumai būdingas lytinis procesas, tarpinių formų nebuvo. Nustatyta, kad dauguma dumblių tam tikru etapu turi undolipodiją gyvenimo ciklas. Dumblių, pirmuonių, gyvūnų ir augalų judrios struktūros pasirodė esančios vienodos: jos visos susideda iš 0,024 mikrono skersmens mikrovamzdelių. Paaiškėjo, kad mitozinė verpstė, net ir grybuose, kurie neturi undolipdijų, susideda iš tokio pat skersmens baltymų mikrotubulių, kaip ir undolipdijų. Pusę amžiaus atrodė labai tikėtina, kad eukariotų dumbliai išsivystė iš melsvadumblių. Šis botaninis mitas lėmė prielaidas, kurios daugelį metų buvo ir kaip stimulas tyrimams, ir koordinačių sistema, in kuriame buvo atliktas gautų rezultatų vertinimas. Teiginys, kad ryšiai tarp cianobakterijų ir eukariotinių dumblių išnyko, nepalikdami fosilijų pėdsakų ir gyvų relikvijų, tapo vis mažiau įtikinami, nes atsirado naujų duomenų apie gyvus mikrobus ir fosilijas. Pagrindinės tarpinės grandys tarp fotosintetinių prokariotų ir eukariotų neišnyko be pėdsakų, kaip reikalauja botanikos mitas – jų niekada nebuvo. Remiantis nuoseklių endosimbiozių teorija, cianobakterijos iš tikrųjų yra protėviai, bet tik plastidai, o ne likusi eukariotinės ląstelės dalis ir tikrai ne jos branduolys. Ir vis dėlto, sprendžiant branduolio kilmės klausimą, tiesioginės giminės hipotezė ir simbiozės teorija dera. Įmanomos ekstremalios simbiozės teorijos versijos modifikacijos apima, pavyzdžiui, plastidžių, bet ne mitochondrijų, simbiotinės kilmės idėją arba plastidų ir mitochondrijų simbiotinę kilmę, bet ne undolipodiją. Tiesioginės giminės šalininkai taip pat pateikia prielaidą apie branduolio kilmę per simbiozę.
Kritiniai chloroplastų ir mitochondrijų endosimbiotinės kilmės įrodymai.
Reikšmingiausi įrodymai apie svarbiausių eukariotinių ląstelių organelių – chloroplastų ir mitochondrijų kilmę iš prokariotų buvo gauti m. pastaraisiais metais tiriant ribosomų RNR struktūrą. Molekulinis biologas
K. Wozas jau devintajame dešimtmetyje parodė, kad RNR iš augalų chloroplastų ribosomų struktūra yra beveik identiška kai kurių cianobakterijų RNR struktūrai. Tuo pačiu metu grupė mokslininkų nustatė, kad ribosomų RNR, gauta iš skirtingų eukariotų mitochondrijų, yra panaši į kai kurių bakterijų, ypač Paracoccus genties bakterijų, RNR. Šios bakterijos kvėpavimo fermentų rinkinys labai panašus į
gyvūnų mitochondrijų „kvėpavimo takų ansamblis“.
Šie duomenys aiškiai patvirtina labiau tikėtina eukariotinių ląstelių endosimbiotinė kilmė. Endosimbiozės reiškinys vis dar itin plačiai paplitęs gamtoje. Yra žinoma, kad ant kai kurių kūno jūrinės žuvys Dažnai apsigyvena šviečiančios bakterijos, kurios ne tik gauna didelę naudą savo gyvybei, bet ir yra labai naudingos šeimininkams. Visa tai, kas išdėstyta aukščiau, įtikinamai liudija, kad eukariotinės ląstelės yra endosimbiotinės kilmės iš daug mažesnių prokariotinių ląstelių. Tačiau pagrindinis klausimas yra nustatyti ląstelės šeimininkės prigimtį, nes eukariotinės ląstelės turi gerai suformuotą branduolį su apvalkalu, o prokariotai tokio branduolio neturi. Pavyzdžiui, L. Margelis mano, kad ląstelių šeimininkų vaidmenį galėjo atlikti prokariotai – pirmtakai šiuolaikinės bakterijos susiję su mikoplazmomis – labai primityviai sukonstruoti prokariotai, praktiškai neturintys ląstelių sienelių. IN Pastaruoju metu pasirodė ir plačiai aptarinėjami japonų mokslininko darbai
T. Oshima. Remdamasis nuodugniais daugelio pro- ir eukariotų ribosomų RNR struktūros tyrimais, įskaitant daugelį iki šiol mažai ištirtos archajų superkaralystės atstovų, šis autorius daro išvadą, kad ląstelė šeimininkė buvo viena iš šiuolaikinės archejos protėvių. Reikia pasakyti, kad šios superkaralystės atstovai, skirtingai nei bakterijos, turi daug biocheminių ir molekulinių biologinių savybių, bendrų su eukariotais. Naujausiuose T. Oshimos darbuose pateikiami argumentai už tai, kad ląstelė šeimininkė eukariotų atsiradimo metu buvo archėjos protėvis, panašus į vieną iš šiuolaikinių šių prokariotų atstovų – termoplazmą. Koks bus galutinis problemos, susijusios su šeimininko ląstelės, kuri sukėlė eukariotinę ląstelę daugybinės endosimbiozės procese, nustatymo problemos sprendimas, parodys tolesni tyrimai.
Šiuo metu pateikiama populiariausia ir pagrįsta eukariotinių ląstelių kilmės hipotezė
įvairių prokariotų, priklausančių bakterijoms ir archejoms, endosimbiozė, daug kartų pasireiškusi evoliucijos procese per pirmuosius 1,5–2 milijardus gyvų ląstelių egzistavimo Žemėje metų. Lemiamų įrodymų apie endosimbiotinę chloroplastų ir mitochondrijų kilmę gavo pirmaujantys pasaulio biologai, tirdami ribosomų RNR struktūrą. Šie duomenys aiškiai patvirtina didelė tikimybė endosimbiotinė eukariotinių ląstelių kilmė iš daug mažesnių (keliais dydžiais) prokariotinių ląstelių.
LITERATŪRA
1. Sorošo edukacinis žurnalas. Biologija, chemija, geomokslai, fizika, matematika. N 5 1998
2 Margelis L. Simbiozės vaidmuo ląstelių evoliucijoje. Per. iš anglų kalbos, M. MIR 1983 m
3 Mikrobiologijos pagrindai. M. "Medicina" 1992m.
4 Khakhina L.N. Sąvoka A.S. Famintsyn apie simbiozės svarbą L. evoliucijoje; Mokslas, 1981, p. 165–181.
5 Alberts B. Ląstelės molekulinė biologija; Per. iš anglų kalbos. M. Mir 1986 v.1
Terminas Prokariotae (ikibranduoliniai) reiškia mikroorganizmus, turinčius primityvią branduolinių struktūrų organizaciją. Šį pavadinimą 1968 metais pasiūlė kanadiečių mokslininkas Murray.
XX amžiaus antroje pusėje. tapo aišku, kad visi gyvi organizmai su ląstelių struktūra, galima priskirti vienai iš dviejų grupių – prokariotams arba eukariotams. Prokariotinės ląstelės yra paprastesnės – jose nėra branduolio, apsupto apvalkalu. Kurį laiką buvo manoma, kad terminai „prokariotai“ ir „bakterijos“ yra sinonimai ir kad bakterijos yra nepriklausoma ir vieninga gyvų organizmų evoliucijos šaka. Tačiau palaipsniui kaupėsi įrodymai, kad kai kurių bakterijų skirtumai gali būti reikšmingi ir rodo, kad jos neturi glaudaus evoliucinio ryšio. Ypač didelę reikšmę prijungtas prie duomenų apie ribosomų ir transportuojančių ribonukleino rūgščių (RNR) struktūrą. Šiuo metu remiantis ribosomų RNR molekulių bazinių sekų palyginimo rezultatais, sprendžiama apie organizmų ryšį. Tokie duomenys, kaip ir kitos prokariotų savybės, parodė, kad iš tikrųjų prokariotai turėtų būti suskirstyti į bent dvi grupes, turinčias skirtingą kilmę ir evoliucijos kelius. Be to, eukariotinės ląstelės yra ne tik sudėtingiau organizuotos, bet ir turi elementų, kurių negalėjo gauti iš šiuolaikinių prokariotų protėvių. Tai visų pirma citoplazminės ribosomos, kurios skiriasi nuo prokariotinių ribosomų. Esminis panašumas genetinis kodas, makromolekulių organizacija ir biocheminis baltymų sintezės aparatas rodo visų gyvų organizmų kilmės vienovę. Spėjama, kad egzistavo tam tikras bendras protėvis – „progenotas“, bet koks jis buvo, nežinoma. Šis progenotas gali sukelti tris nepriklausomas evoliucinio medžio šakas. Hierarchinės sistematikos požiūriu šias šakas siūloma laikyti sritimis, turinčiomis aukštesnį rangą nei tradicinės karalystės. Šios sritys (imperijos) yra eubakterijos, archebakterijos, eukariotos (eubakterijos, archebakterijos ir eukariotai). Bendrosios ir skiriamosios archebakterijų, eubakterijų ir eukariotų savybės pateiktos lentelėje.
Pagrindinis prokariotinės ląstelės bruožas yra branduolio, kurį nuo citoplazmos riboja dviguba membrana, nebuvimas. Paveldima medžiaga yra koncentruota bakterijų chromosomoje, paprastai vaizduojama kaip apskrita dvigrandė DNR molekulė. Kai kurios spirochetos turi linijines chromosomas, o aktinomicetai turi „pseudožiedo“ chromosomas. Į siūlą panaši DNR molekulė veikia kaip branduolys ir yra centrinėje ląstelės zonoje, vadinamoje „nukleoidu“. Elektroniniame mikroskope ant bakterinės ląstelės dalies ši zona atrodo šviesesnė nei likusi citoplazmos dalis.
Prokariotinė ląstelė turi keletą pagrindinių savybių, susijusių tiek su jos ultrastruktūrine, tiek su cheminė organizacija. Visų pirma, bakterijų ląstelėje nėra membraninio tinklo – endoplazminio tinklo. Prokariotinėse ląstelėse citoplazminė membrana, esanti po ląstelės sienele ir iš išorės apgaubianti protoplastą, yra vienintelė ląstelės membrana.
Kiekvieną ląstelę supanti citoplazminė membrana lemia jos dydį ir užtikrina, kad išliktų reikšmingi ląstelių turinio ir aplinkos skirtumai. Tačiau membrana nėra tik mechaninė pertvara. Jis taip pat tarnauja kaip labai selektyvus filtras, kuris palaiko skirtingą jonų koncentraciją abiejose membranos pusėse ir leidžia maistinėms medžiagoms patekti į ląstelę ir pašalinti atliekas. Citoplazminė membrana yra lipidų ir baltymų molekulių, laikomų kartu nekovalentine sąveika, visuma. IN elektroninis mikroskopas membranos atrodo kaip maždaug 7 nm storio lakštai su ryškia trijų sluoksnių struktūra. Dvigubas lipidų molekulių sluoksnis lemia membranos struktūrines ypatybes, o baltymai yra atsakingi už daugumą membranos funkcijų.
Pagrindinės archebakterijų, eubakterijų ir eukariotų savybės
| ženklai | archebakterijos | eubakterijos | eukariotų |
| Tipiški organizmai | metanogenai, ekstremalūs termofilai, halofilai | Pagrindinės bakterijų grupės | protistai, grybai, augalai, gyvūnai |
| Tipiniai matmenys, mikronai | 0,5-4 | 0,5-4 | virš 5 |
| Genomas | žiedinė chromosoma | žiedinė chromosoma | branduolys su daugybe sudėtingų chromosomų |
| Membraninės struktūros: endoplazminis tinklas Golgi kompleksinės lizosomos | ne ne ne | ne ne ne | taip taip taip |
| Mitochondrijos | Nr | Nr | Yra |
| Chloroplastai | nėra (kartais tilakoidai) | Nr | ne (grybuose ir gyvūnuose), taip (augaluose ir dumbliuose) |
| Ląstelių sienelės | baltymai, pseudomureinas | mureinas, lipopolisacharidas | įvairių |
| Ribosomos | 70S* | 70S | 80S (citoplazmoje) + 705 organelių ribosomos, kaip ir bakterijose |
| RNR polimerazė | kompleksas | paprastas | kompleksas |
| Flagella | paprastas | paprastas, sudarytas iš vienos ar kelių fibrilių | kompleksinė struktūra (2х9)+2 |
| Azoto fiksacija | Yra | Yra | Nr |
| Anaerobinis kvėpavimas | Yra | Yra | Nr |
| Chemosintezė | Yra | Yra | Nr |
S* – Svedbergo sedimentacijos vienetas, apibūdinantis ribosomos dydį
Prokariotų citoplazminėje membranoje yra daugybinės invaginacijos – invaginacijos, kurios atlieka skirtingas funkcijas. Juose gali būti fermentų ir jie atrodo kaip pūslelės arba yra mezosomos ir dalyvauja ląstelių dalijimuisi. Prokariotų citoplazminė membrana yra energijos apykaitos fermentų lokalizacijos vieta, todėl ji atlieka mitochondrijų, kurių nėra prokariotinėse ląstelėse, funkciją. Todėl bakterijos su aukštas lygis kvėpavimo medžiagų apykaita, ypač išvystyta citoplazminė membrana, t.y. jis turi daug invaginacijų, todėl padidėja jo paviršius. Citoplazminė membrana taip pat dalyvauja dalijantis nukleoidą. Žiedinė DNR fiksuojama ant membranos ir po replikacijos (dvigubėjimo) dėl membranos augimo išsiskiria į dvi naujas ląsteles. Endosporų dangteliai susidaro iš citoplazminės membranos. Tai taip pat yra pagrindinė kliūtis medžiagoms prasiskverbti į ląstelę. Citoplazminė membrana nepraleidžia didelių molekulių, ji nepajėgi pinocitozės ir egzocitozės.
Už citoplazminės membranos ribų esančios struktūros – ląstelės sienelė, kapsulė, gleivinė, žvyneliai, fimbrijos, piliai, gaureliai – yra paviršinės struktūros.
Ląstelės sienelė yra svarbus ir privalomas daugumos prokariotinių ląstelių (tikrųjų bakterijų, neskaitant archebakterijų ir mikoplazmų) struktūrinis elementas, esantis po kapsule ar gleivine arba, jei jos nėra, kontaktuojantis su aplinka. Ląstelės sienelė tarnauja kaip mechaninis barjeras tarp protoplasto ir išorinė aplinka ir suteikia ląstelei tam tikrą jai būdingą formą.
Pagal struktūrą ir cheminę sudėtį prokariotų ląstelių sienelė labai skiriasi nuo eukariotinių organizmų. Prokariotų ląstelės sienelė susideda iš kelių sluoksnių, iš kurių pagrindinis – karkasinis – sluoksnis, atsakingas už stiprumą, susidaro iš specialios medžiagos – mureino peptidoglikano, kurį sintetina tik prokariotinė ląstelė, o kituose organizmuose nėra. Tikrųjų prokariotų grupėje išskiriami du pagrindiniai ląstelės sienelės sandaros tipai. Šie du tipai atitinka gramteigiamas ir gramneigiamas bakterijas. Šių grupių pavadinimai kilę iš gebėjimo ar nesugebėjimo skirtingos bakterijos dėmė pagal gramą. 1884 metais danų gydytojas K. Gramas, užsiėmęs audinių dažymu, pasiūlė dažymo būdą, kuris vėliau buvo panaudotas bakterijoms. Jei fiksuotos eubakterijų ląstelės pirmiausia apdorojamos krištolinio violetinio, o po to jodo tirpalu, susidaro spalvotas kompleksas. Vėlesnio gydymo alkoholiu metu, priklausomai nuo ląstelės sienelės sandaros, komplekso likimas skiriasi: vadinamosiose gramteigiamosiose rūšyse šis kompleksas išlaikomas ląstelėje, o pastarosios išlieka spalvotos, gramneigiamos. rūšių, spalvotas kompleksas išplaunamas iš ląstelės, ir jie pasikeičia. Nustatyta, kad spalvotas kompleksas susidaro ant protoplasto, tačiau jo sulaikymą ląstelėje arba išplovimą iš jos vėlesnio apdorojimo alkoholiu metu lemia ląstelės sienelės struktūriniai ypatumai. Rastas gramo dažymo metodas platus pritaikymas mikrobiologinėje praktikoje bakterijoms diferencijuoti.
Gramteigiamų ir gramneigiamų eubakterijų ląstelių sienelės labai skiriasi tiek chemine sudėtimi, tiek ultrastruktūra.
Didžiąją gramteigiamų bakterijų ląstelės sienelės dalį sudaro specifinis heteropolimeras – peptidoglikano mureinas. Molekulės polisacharidinis pagrindas sudarytas iš kintančių N-acetilgliukozamino ir N-acetilmuramo rūgšties liekanų, tarpusavyje sujungtų glikozidinėmis jungtimis. Be peptidoglikano, gramteigiamų eubakterijų ląstelių sienelėse yra techoinės rūgštys ir polisacharidai. Teichoinės rūgštys yra polimerai, kurių pagrindą sudaro polihidroksiliai alkoholiai(ribitolis ir glicerolis), tarpusavyje sujungti fosfodiesterio ryšiais.
Gramneigiamos eubakterijos turi daug sudėtingesnę ląstelių sienelių struktūrą nei gramteigiamos. Jame yra daug daugiau skirtingų makromolekulių cheminis tipas. Peptidoglikanas sudaro tik vidinį ląstelės sienelės sluoksnį, laisvai prilipdamas prie citoplazminės membranos. Už peptidoglikano yra papildomas ląstelės sienelės sluoksnis - išorinė membrana, kuris susideda iš fosfolipidų (būdingų elementarioms membranoms), baltymų, lipoproteinų ir lipopolisacharido.
Nustatyta, kad empiriškai nustatyti skirtumai tarp dviejų bakterijų grupių, susijusių su Gramo dėmėmis, koreliuoja su daugeliu kitų svarbių bruožų. Pavyzdžiui, gramteigiamos bakterijos dauginasi dvejetainiu dalijimusi dėl to, kad ląstelėje susidaro skersinė pertvara, gramneigiamos – susidarius susiaurėjimui. Pirmieji jautrūs penicilinui, antrieji – ne. Gram-neigiamos bakterijos nesudaro endosporų, o kai kurios gramteigiamos – formuoja.
Prokariotų ląstelės sienelė atlieka įvairias funkcijas: mechaniškai apsaugo ląstelę nuo aplinkos poveikio, išlaiko išorinę formą, leidžia ląstelei egzistuoti hipotoniniuose tirpaluose. Visų pirma, tai yra peptidoglikano „nuopelnas“. Medžiagų pralaidumas ir transportavimas į ląstelę yra susijęs su ląstelės sienele.
Po citoplazmine membrana bakterijose yra citoplazma. Tai koloidinė sistema, susidedanti iš vandens, baltymų, riebalų, angliavandenių, mineralinių junginių ir kitų medžiagų, kurių santykis kinta priklausomai nuo bakterijų rūšies ir jų amžiaus. Prokariotinės ląstelės citoplazma nėra pavaldi cikliniam judėjimui, kaip augaluose, ar susitraukimo reiškiniams, kaip amebose. bakterijų ląstelės atlieka judėjimą dėl kitų mechanizmų: arba paprasčiausiai išdėstytų žvynelių, kurie skiriasi nuo sudėtingų eukariotų žvynelių, pagalba jie aktyviai plaukia skystoje terpėje arba slysta, išskirdami gleives ir judėdami tankaus substrato paviršiumi.
Judrios bakterijų ląstelės aktyviai juda tam tikrų veiksnių nustatyta kryptimi. Tokie nukreipti bakterijų judėjimai vadinami „taksi“. Yra chemotaksis, fototaksis, magnetotaksis, termotaksis, viskozitaksis (atsakas į tirpalo klampumo pasikeitimą).
Ramias bakterijų ląsteles gramteigiamose bakterijose reprezentuoja endosporos, gramneigiamose – cistos. Bakterijų sporos turi unikalų atsparumą neigiamam poveikiui. Ypač į akis krenta bakterijų sporų atsparumas karščiui: kai kurios sporos pakenčia virimą 1 valandą ar ilgiau ir nepraranda gyvybingumo. Labai atsparus sporoms ir džiūvimui. Sporų stabilumas yra susijęs su specialaus sluoksnio buvimu jų ląstelės sienelėje - žievėje.
Sporų susidarymo metu bakterijų ląstelės kartais įgauna neįprastą verpstės, citrinos ar būgnų lazdelė. Kiekvienoje ląstelėje paprastai yra viena endospora, tačiau yra bakterijų, turinčių daugiau nei keturias sporas. Kai kurie anaerobinės bakterijos ant sporų susidaro ląsteliniai dangteliai, sudaryti iš dujų vakuolių arba įvairių rūšių ataugų
Bakterijų sporos nėra vienintelės prokariotų ramybės formos. Gramą skaitančios bakterijos, kurios nesugeba susidaryti endosporų, kai kuriais atvejais virsta cistomis - specialios ląstelės su storomis išorinėmis dangomis (pvz. , Azotobacter chroococcum)
Išorėje prokariotų ląstelių sienelės dažnai yra apsuptos gleivinės medžiagos. Tokie dariniai, priklausomai nuo struktūrinių ypatybių, vadinami kapsulėmis, gleiviniais sluoksniais, apvalkalais arba apvalkalais (gijinėse bakterijose). Visi jie yra prokariotų organinių polimerų biosintezės ir jų nusėdimo aplink ląsteles rezultatas.
Kapsulėse, gleivių dariniuose ir apvalkaluose gali būti komponentų, kurie yra tokie patys kaip ląstelės sienelė, bet jie cheminė sudėtis nėra tapatus. Paprastai bakterijų suformuotų kapsulių cheminė sudėtis yra būdinga genčiai ir rūšiai. Pagrindiniai daugumos prokariotinių kapsulių cheminiai komponentai yra polisacharidai. Išimtis yra kai kurių rūšių bacilų kapsulės, pagamintos iš polipeptido, kuris yra glutamo rūgšties polimeras. Bylų – daugiau sudėtingos struktūros, jie yra sudėtingesnės cheminės sudėties ir juose yra cukrų, baltymų, lipidų.
Kapsulės yra skirtingo storio ir lengvai atpažįstamos
po mikroskopu su neigiamu preparato kontrastu juodu rašalu.
Kapsulės ir dangteliai apsaugo ląstelę nuo pažeidimų ir išsausėjimo, sukuria papildomą osmosinį barjerą, trukdo fagams prasiskverbti, aplink ląsteles sukuria ypatingą aplinką, kuri suteikia tam tikrą vandens režimas. Kapsulės atlieka specifines funkcijas. Pavyzdžiui, gumbelinėse bakterijose jos lemia selektyvų prisirišimą prie įvairių rūšių augalų šaknų plaukų. Kartais kapsulės gali būti naudojamos kaip atsarginės maistinės medžiagos. Gleivių pagalba vyksta ryšys tarp kolonijos ląstelių, ląstelių pritvirtinimas prie įvairių paviršių
Kapsuliniai polisacharidai sąveikauja su dirvožemio tirpalų katijonais, gali sunaikinti dirvožemio mineralus, dalyvauti agregacijos procesuose ir formuojant vandeniui atsparią dirvožemio struktūrą. Galbūt mikrobų tarpląstelinių gleivių įtraukimas į huminių medžiagų susidarymo procesą.
Prokariotų ląstelių forma yra labai įvairi. Žinomi prokariotai yra sferiniai, lazdelės formos, susukti spirale, kablelio pavidalo, žiedo formos – uždari arba atviri, su ataugomis – paprasti. Ląstelės gali būti kvadratinės arba žvaigždės formos. Daugialąsčiai prokariotai yra gijų arba sliekų formos.
Visi prokariotai priklauso mikroskopinių organizmų grupei, jų ląstelių dydis matuojamas mikronais (mikrometrais) (1 μm = 10 3 mm), o jų detalės matuojamos nanometrais (1 nm = 10 mm). Beveik 40% bakterijų dirvožemyje nepatenka į šviesos mikroskopą, jos matomos tik elektroniniu mikroskopu. Esant tokiems mažiems dydžiams, paviršiaus ir tūrio santykis yra labai didelis, todėl galimas didelis medžiagų apykaitos greitis.
Visi prokariotai yra suskirstyti į keturias grupes pagal išorinio ląstelės apvalkalo struktūrą: gramneigiamas (Gracilicutes) eubakterijas, gramteigiančias (Firmicutes) eubakterijas, mikoplazmas (Mollicutes), neturinčias ląstelių sienelių ir archebakterijas arba archejas. (archebakterijos). Pirmosios trys grupės yra sujungtos kaip eubakterijos (Eubacteria), archebakterijos yra atskira organizmų grupė (Archaebacteria), kurios skiriasi nuo kitų ribosomų ir pernešančių RNR molekulių sandara, RNR polimerazėmis, lipidais, ląstelės sienelės sandara.
Gramneigiamos bakterijos turi ląstelių sieneles, kuriose, be pagrindinio mureino sluoksnio, yra ir išorinė membrana bei baltymo ir (arba) glikoproteino sluoksnis, dengiantis jį iš viršaus. Tarp citoplazminės membranos ir mureino sluoksnio yra periplazminė erdvė. Ši bakterijų grupė yra pati didžiausia ir įvairiausia pagal rūšių skaičių ir fiziologines funkcijas. Prokariotų grupė jungia fototrofines ir chemotrofines bakterijas. Tarp jų yra paprasti saprotrofai, kopiotrofai, oligotrofai, kitų bakterijų viduląsteliniai parazitai, patogeninės formos žmonėms, gyvūnams ir augalams, turintiems sudėtingų mitybos poreikių. Pagal morfologiją tai yra įvairios bakterijos nuo paprastų lazdelės formos pseudomonadų iki miksobakterijų, turinčių sudėtingą vystymosi ciklą.
Mikoplazmos – tai ląstelės sienelės neturinčios bakterijos, kurių funkciją atlieka citoplazminė membrana.
Archebakterijos vienija skirtingą išorinio apvalkalo struktūrą turinčius prokariotus, kuriems trūksta tikrosioms bakterijoms būdingo mureino. Kai kurių archebakterijų atstovų ląstelės sienelėje yra pseudomureino, o kitų – ląstelės sienelė, sudaryta iš rutulinių baltymų arba glikoproteinų sluoksnio.
Bakterijų skirstymas į grupes pagal fenotipinius požymius (morfologiją, fiziologiją) patogus praktiniais tikslais greitam organizmų identifikavimui. Organizmai yra sugrupuoti į dirbtines grupes arba "skyrius", kurie neturi taksonominio statuso ir dažnai yra žymimi būdinga rūšis arba titulą. Pagal šį principą yra sukurtas Burgey bakterijų determinantas.
Tyrinėdami Žemėje gyvenusių ir gyvenančių organizmų įvairovę, taksonomikai siekia sukurti filogenetinę sistemą, kuri atspindėtų į vieną ar kitą grupę – taksoną – susijungusių organizmų genealoginį ryšį ir bendrą kilmę. Šiam tikslui pasitarnauja molekulinė sistematika, pagrįsta biopolimerų – DNR, RNR, baltymų – sandaros panašumu. Dėl galimybės nustatyti genų panašumą jis dažnai vadinamas genų sistematika. Vienas svarbiausių genų sistematikos pasiekimų yra naujos atskiroje karalystėje išskirtos mikroorganizmų grupės – archebakterijų – atradimas. Bakterijų genomų panašumo laipsnis nustatomas lyginant nukleotidų sekas 16S rRNR molekulėje. Šio metodo panaudojimas bakterijų taksonomijoje buvo pagrindas K. Wese'ui sukurti filogenetinį bakterijų medį.
Labai sunku palyginti filogenetinę ir fenotipinę sistematiką. Tose pačiose filogenetinėse šakose yra fiziologijos požiūriu labai įvairių organizmų. Pereinant nuo didelių gramteigiamų ir gramneigiamų bakterijų šakų į žemesnius hierarchijos lygius, tokių neatitikimų vis daugėja.
KURSINIS DARBAS
biologijoje
„Eukariotų kilmė
Užbaigė: Severova Vera
10 klasė, mokykla N55
Patikrintas: Strukova N.A.
Riazanė 1999 m
1 . Simbiotinė teorija:
1.1 Pagrindinis skirtumas tarp prokariotų ir eukariotų;
Prokariotiniai organizmai;
eukariotinės ląstelės;
mitochondrijos ir plastidai;
eukariotų gebėjimas fotosintezei;
2. Gyvybės pirmtakai;
3. Fermentuojantys organizmai:
Ankstyvųjų ląstelių energijos šaltiniai;
Metanogenezė kaip energijos šaltinis;
cianobakterijos ir chloroksibakterijos;
Fotosintezė ir oras;
Asociacijos ir eukariotai:
Viena po kitos einančių simbiozių teorijos;
Simbiotinis mobilumo įgijimas;
Eukariotai yra fotosintetiniai;
Žvilgsnis į ląstelės evoliuciją:
6.1 Tiesioginės giminystės hipotezė;
Hipotezė: nuo cianobakterijų iki dumblių;
2 hipotezių suderinamumas: tiesioginė giminystė ir
endosimbiozė;
Literatūra.
ENDOSIMBIOZĖS TEORIJA Visi gyvi organizmai Žemėje paprastai skirstomi į prokariotus ir eukariotus. Pagrindinis prokariotų bruožas, skirtingai nuo eukariotų, yra pilnaverčio ląstelės branduolio, padengto membrana, trūkumas. Prokariotinės ląstelės yra labai mažo dydžio, maždaug 1 µm. Eukariotinių ląstelių, kuriose yra pilnavertis branduolys, tūris yra 800–1000 kartų didesnis nei prokariotinių ląstelių tūris. Šiuo atžvilgiu ląstelėms – eukariotams gyvybinėms funkcijoms atlikti reikalingas daug didesnis membraninis aparatas. Iš tiesų, eukariotų ląstelėje buvo nustatytos tarpląstelinės struktūros, apsuptos membranų, nepriklausomų nuo plazmalemos ir viena kitos. Šios struktūros vadinamos organelėmis. Kiekviena organelė: mitochondrijos, chloroplastas, branduolys, lizosomos, Golgi aparatas ir kt. turi savo specifines funkcijas ląstelės gyvenime. Molekulinių-biologinių metodų pagalba buvo gauta įtikinamų įrodymų, kad visos gyvos būtybės turėtų būti suskirstytos į 3 karalystes: bakterijas, archajas ir eukariotus. Bakterijos ir archėjos Žemėje atsirado iš bendro protėvio – vadinamojo progenoto – maždaug prieš 4 milijardus metų, o paleontologų teigimu, eukariotai Žemėje atsirado praėjus 500 milijonų metų po prokariotų. Populiariausia ir pakankamai pagrįsta eukariotinių ląstelių kilmės idėja šiuo metu yra įvairių prokariotų daugialypės endosimbiozės idėja. Žinomų mikrobiologų A. Kluyverio ir K. Van Nielio teigimu, būtent senovėje prokariotuose vyko ekonomiškiausių ir dažnai polifunkcinių biocheminių reakcijų pasižyminčių ląstelių atranka, kuri tapo svarbiausių medžiagų apykaitos kelių pagrindu.
Simbiotinė ląstelių atsiradimo ir evoliucijos teorija remiasi dviem naujomis biologijoje sampratomis.Pagal pirmąją iš šių sąvokų esminis skirtumas gyvojoje gamtoje yra skirtumas tarp prokariotų ir eukariotų, t.y. tarp bakterijų ir organizmų, susidedančių iš ląstelių, turinčių tikrus branduolius – protistų, gyvūnų, grybų ir augalų. Antroji koncepcija yra ta, kad kai kurios eukariotinių ląstelių dalys atsirado dėl simbiozių evoliucijos, t.y. nuolatinių asociacijų tarp skirtingų rūšių organizmų susidarymas. Daroma prielaida, kad trys organelių klasės – mitochondrijos, blakstienos ir fotosintetinės plastidės – atsirado iš laisvai gyvenančių bakterijų, kurios dėl simbiozės pateko į tam tikrą seką prokariotinių šeimininkų ląstelėse. Ši teorija labai remiasi neodarvinistinėmis idėjomis, kurias sukūrė genetikai, ekologai, citologai ir kiti mokslininkai, kurie susiejo Mendelio genetiką su darvinietiška natūralios atrankos idėja. Ji taip pat remiasi visiškai naujomis arba neseniai atgaivintomis mokslo kryptimis: molekuline biologija. , ypač dėl baltymų struktūros ir aminorūgščių sekos duomenų, apie mikropaleontologiją, tiriančią ankstyviausius gyvybės pėdsakus Žemėje, ir net apie atmosferos fiziką bei chemiją, nes šie mokslai yra susiję su biologinės kilmės dujomis. Visi ląsteliniai organizmai gali būti suskirstyti į penkias karalystes: prokariotų karalystę (Monera, kuri apima bakterijas) ir keturias eukariotų karalystes (Protoctista, Animalia, Fungi, Plantae). Protokistai yra eukariotiniai organizmai, kurie nėra gyvūnai, grybai ar augalai. Protokistų karalystei priklauso dumbliai, pirmuonys, gleivių pelėsiai ir kiti neaiškios kilmės eukariotiniai organizmai. Protistai labiau apibrėžiami kaip vienaląsčiai eukariotai. Taigi į karalystę Protoctista priklauso ne tik protistai – vienaląsčiai eukariotai, bet ir artimiausi daugialąsčiai jų palikuonys, tokie kaip raudonieji ir rudieji dumbliai, taip pat daugybė į grybus panašių mikroorganizmų, tokių kaip chitridai. Remiantis tradiciniu tiesioginės filiacijos požiūriu, ląstelių organelės, tokios kaip mitochondrijos ir plastidai, atsirado suskaidžius pačią ląstelę.
KELI PROKARIOTAI SUDARIA VIENĄ EUKARIOTĄ
Ląstelės yra membranomis surištos sistemos, mažiausios iš biologinių vienetų, galinčių savarankiškai daugintis. Net mažiausiose ląstelėse yra genų DNR molekulių pavidalu (viena ar daugiau) ir baltymų sintezės aparatas, susidedantis iš kelių tipų RNR ir daugybės baltymų. Visose ląstelėse yra ribosomų, maždaug 0,02 µm skersmens kūnų, sudarytų iš mažiausiai trijų tipų RNR ir apie penkiasdešimt skirtingų baltymų. Prokariotiniai organizmai yra mažiausi biologiniai vienetai, atitinkantys šį ląstelės apibrėžimą. Jie neturi branduolių. Kita vertus, pavienės eukariotinės (turinčios branduolį) ląstelės tam tikra prasme nėra vienetai. Tai įvairių baltymus sintezuojančių vienetų (nukleocitoplazmos, mitochondrijų ir plastidų) kompleksai, kurių tipas ir skaičius skiriasi priklausomai nuo rūšies. Taigi, remiantis šia teorija, eukariotinės ląstelės atsirado bendradarbiaujant iš pradžių nepriklausomiems elementams, susijungusiems tam tikra tvarka. Taylor šią idėją apie atskirą eukariotinės ląstelės dalių kilmę ir vystymąsi bei vėlesnį jų suvienijimą pavadino nuoseklių endosimbiozių teorija. Aukštos temperatūros ir rūgštingumo sąlygos paskatino nukleocitoplazmoje gamintis specialios klasės baltymai – histonai – apsaugoti jos DNR. Mitochondrijos sugebėjo suskaidyti trijų anglies junginius į anglies dioksidą ir vandenį. Simbiotinė nukleocitoplazmos ir mitochondrijų asociacija pradėjo keistis abipusės priklausomybės stiprėjimo kryptimi. Šis kompleksas apėmė motorines organeles - blakstienas ir žiuželes. Po šios transformacijos sekė morfogenetinė transformacija, kurią sudarė ryški ląstelių asimetrija. Integravus tarpusavyje priklausomas nukleocitoplazmas, mitochondrijas, undulipodijas, buvo baigta sukurti eukariotinės ląstelės poligenominė struktūra. Pirmoji eukariotinė ląstelė buvo heterotrofinė. Ji valgė jau paruoštus organinius junginius. Šiuose eukariotuose atsirado mitozės procesai, o vėliau - mejozė. Ir galiausiai, eukariotai įgijo gebėjimą fotosintezei tapti pagrindine eukariotų organizacija, o po jos užbaigimo - paskutiniu šio epo įvykiu. Simbiozės vis labiau integravosi. Šiuo metu kiekvienos organelės priklausomybė nuo kitų organelių medžiagų apykaitos produktų tapo tokia išsami, kad tik šiuolaikiniai analizės metodai leidžia atsekti pirminių partnerių medžiagų apykaitos kelius. Eukariotų struktūra yra įvairi, tačiau metaboliškai jie išlieka vienodi. Eukariotų medžiagų apykaitos gudrybės buvo sukurtos įvairių bakterijų dar prieš tai, kai jos susijungė ir tapo šeimininku ir jo organelėmis. Prokariotai pasiekė didelę energijos gavimo būdo ir biosintezės sudėtingumo įvairovę. Jie paliko savo egzistavimo pėdsaką gerokai anksčiau, nei iš eukariotinės ląstelės susiformavo tokie dideli organizmai kaip gyvūnai su skeletu.
Eukariotinių ląstelių kilmės simbiozės būdu schema
GYVYBĖS PREKURSAI
Šiandieninių organizmų protėviai atsirado daugiau nei prieš tris milijardus metų iš organinių junginių, susidariusių Žemės paviršiuje. Šios koncepcijos patrauklumą iš dalies lemia jos patikrinamumas.
S. Milleris ir H. Urey parodė, kad tokiomis atmosferos sąlygomis, kurios vyko jaunoje Žemėje, gali susidaryti aminorūgštys ir kitos organinės molekulės. Per pirmuosius milijardus Žemės egzistavimo metų spontaniškai susiformavo įvairių tipų organinės molekulės. Daug organinių molekulių buvo rasta tarpžvaigždinėje erdvėje ir meteorituose. Tai reiškia, kad organiniai junginiai atsiranda sąveikaujant šviesos elementams, esant energijos šaltiniams. Cheminio biologinio dauginimosi mechanizmo universalumas reiškia, kad visa gyvybė Žemėje kilusi iš bendrų protėvių: visa dabartinė gyvybė kilo iš ląstelių su informacinėmis sistemomis, pagrįstomis DNR replikacija ir baltymų sinteze, kuriai vadovauja pasiuntinio RNR.
RŪGINGI ORGANIZMAI
Gali būti, kad nukleorūgštys nuo pat pradžių buvo atskirtos nuo aplinkos lipoproteininėmis membranomis. Informaciją pradėjo koduoti nukleotidų seka DNR molekulėse. Seniausios ląstelės ATP ir aminorūgštis, kurios yra baltymų dalis, naudojo kaip tiesioginius energijos šaltinius ir kaip struktūrinius komponentus. Taigi - seniausia gyvybė egzistavo heterotrofinių bakterijų pavidalu, kurios maistą ir energiją gaudavo iš abiotinės kilmės organinių medžiagų. Neodarvinistinei organinei evoliucijai reikalingi trys reiškiniai: dauginimasis, mutacija ir selektyvus aplinkos spaudimas. Dauginimasis yra nepritaikoma biologinės evoliucijos sąlyga. Pakankamai tiksliai dauginantis, mutacijos ir natūrali atranka yra neišvengiamos. Paaiškinti tokio dauginimosi kilmę reiškia suprasti pačios gyvybės kilmę. Mutacija, pagrindinis evoliucijos kintamumo šaltinis, sukelia paveldimus pokyčius savaime besidauginančioje sistemoje. Sukūrus patikimą reprodukciją, prasidėjo neodarvinizmo evoliucija. Organizmai, sukūrę ląstelei reikalingų komponentų sintezės būdus, turėjo pranašumą prieš kitus. Anaerobinės metaną gaminančios bakterijos buvo seniausi autotrofai, tai yra organizmai, galintys patenkinti savo energijos ir organinių medžiagų poreikius iš neorganinių šaltinių. Fotosintezės gebėjimas pirmiausia buvo sukurtas deguoniui jautriose bakterijose. Sugerdami saulės šviesą bakterinio chlorofilo pagalba, jie atmosferos anglies dioksidą pavertė organinėmis medžiagomis.Anaerobinės fotosintezės atsiradimas negrįžtamai pakeitė Žemės paviršių ir atmosferą.
Anglies dioksidas buvo pašalintas iš atmosferos ir paverčiamas organine medžiaga pagal bendrąją fotosintezės lygtį: 2H2X+CO2=CH2O+2X+H2O. Fotosintetinės bakterijos, kurios naudojo vandenilio sulfidą CO2 nusėdusios elementinės sieros mažinimui. Iš jų atsirado bakterijų, galinčių fotosintezuoti, išskirdamos deguonį. Šio tipo fotosintezė tikriausiai atsirado dėl fotosintetinėse sieros bakterijose susikaupusių mutacijų. Šis kelias paskatino melsvadumblių arba melsvadumblių atsiradimą. Neseniai buvo atrasta ir ištirta nauja fotosintetinį O2 gaminančių prokariotų grupė – chlorooksibakterijos. Galima daryti prielaidą, kad dumblių ir augalų ląstelėse randamų plastidų protėviai buvo ciano ir chloroksibakterijos. O2 susikaupus dėl mikrobų fotosintezės, žemės atmosfera oksidavosi, o tai sukėlė visos gyvybės krizę – iki tol pasaulyje gyveno anaerobinės bakterijos. Norėdami išgyventi, mikrobai turėjo vengti deguonies, slėpdamiesi pelkėse, arba jie turėjo sukurti medžiagų apykaitą, galinčią apsaugoti juos nuo didėjančio deguonies kiekio.
FOTOSINTEZĖ IR ORO
Maždaug prieš 2 milijardus metų vienas po kito vyko įvairūs evoliuciniai procesai, sukuriantys medžiagų apykaitos įvairovę. Gebėjimas gaminti specifinius fermentus gali būti perduodamas toje pačioje rūšyje naudojant į virusą panašias daleles, vadinamas plazmidėmis. Prokariotų biosintetiniai gebėjimai stebina: iš anglies dioksido, kelių druskų ir redukuotų neorganinių junginių, kurie tarnauja kaip energijos šaltiniai, chemoautotrofiniai mikroorganizmai gali susintetinti deguonies turinčiame neapšviestame vandenyje visas sudėtingiausias augimui ir dauginimuisi reikalingas makromolekules. Šie biocheminiai procesai paskatino elementų cirkuliacijos sistemas net tada, kai nebuvo augalų, grybų, gyvūnų.
asociacijos ir eukariotai.
Žvilgsnis į ląstelės evoliuciją.
Tiesioginis filialas.
Tradicinis požiūris į eukariotinių ląstelių kilmę ir evoliuciją yra toks, kad visi gyvi organizmai yra tiesiogiai kilę iš vienos protėvių populiacijos dėl natūralios atrankos įtakoje susikaupusių mutacijų.
Taškinės mutacijos, dubliacijos, delecijos ir kiti paveldimi pokyčiai, kurie, kaip žinoma, turėjo įtakos gyvūnų ir augalų evoliucijai, taip pat yra atsakingi už eukariotinių ląstelių diferenciaciją nuo prokariotinių – tiesioginės giminės teorija. y., nesimbiotinės ląstelių kilmės samprata. Galbūt žalieji dumbliai yra visų augalų protėvių palikuonys. Daugelis šių dumblių savybių rodo, kad jie ir augalai turi bendrą protėvį. Gyvūnų filogenija yra sudėtingesnė, tačiau tiesioginės giminystės sąvoka tikriausiai taikoma ir jų evoliucijos keliams. Kempinės, matyt, išsivystė iš protistų, nepriklausomai nuo kitų gyvūnų grupių. Taigi, bent jau gyvūnams ir augalams, hipotezė, kad sudėtingi eukariotiniai organizmai išsivystė iš paprasčiau išsidėsčiusių eukariotų, susikaupus individualioms mutacijoms, veikiant natūraliai atrankai, yra gana pagrįsta. Evoliucinės naujovės remiasi labai tobulų palankių genų išsaugojimu. Taigi, nauji organizmai, turintys didesnį gebėjimą kontroliuoti savo aplinką, labiau išsivystė išlaikant labai pritaikytų genų dubliavimąsi ir rekombinaciją, o ne įgydami naujų savybių per paprastas, atsitiktines taškines mutacijas. Tiesioginės giminės hipotezė nepaaiškina eukariotų kilmės iš prokariotų. Pavyzdžiui, pertrauka tarp melsvadumblių, negalinčių susirgti mitoze, ir tokių formų, kurių mitozė yra visiškai išsivysčiusi, kaip raudonieji dumbliai, yra sunkiai suprantamas tiesioginės filiacijos požiūriu, tačiau tai natūraliai išplaukia iš simbiotinės teorijos.
Remiantis tiesioginės giminės teorija ir simbiozės teorija, visi organizmai Žemėje kilę iš biochemiškai panašių bakterijų protėvių, o iš eukariotinių mikrobų atsirado augalai ir gyvūnai. Dėl fotosintetinių bakterijų atsirado dumbliai, o galiausiai ir augalai, o kai kurie dumbliai prarado plastidus ir tapo grybų ir gyvūnų protėviais. Tiesioginės giminystės teoretikai sutinka, kad plastidai ir kitos eukariotinės organelės, įskaitant branduolį, susidarė diferencijuojant pačiose ląstelėse, ir vadina tai botaniniu mitu.
Augalų rūšių filogenezė
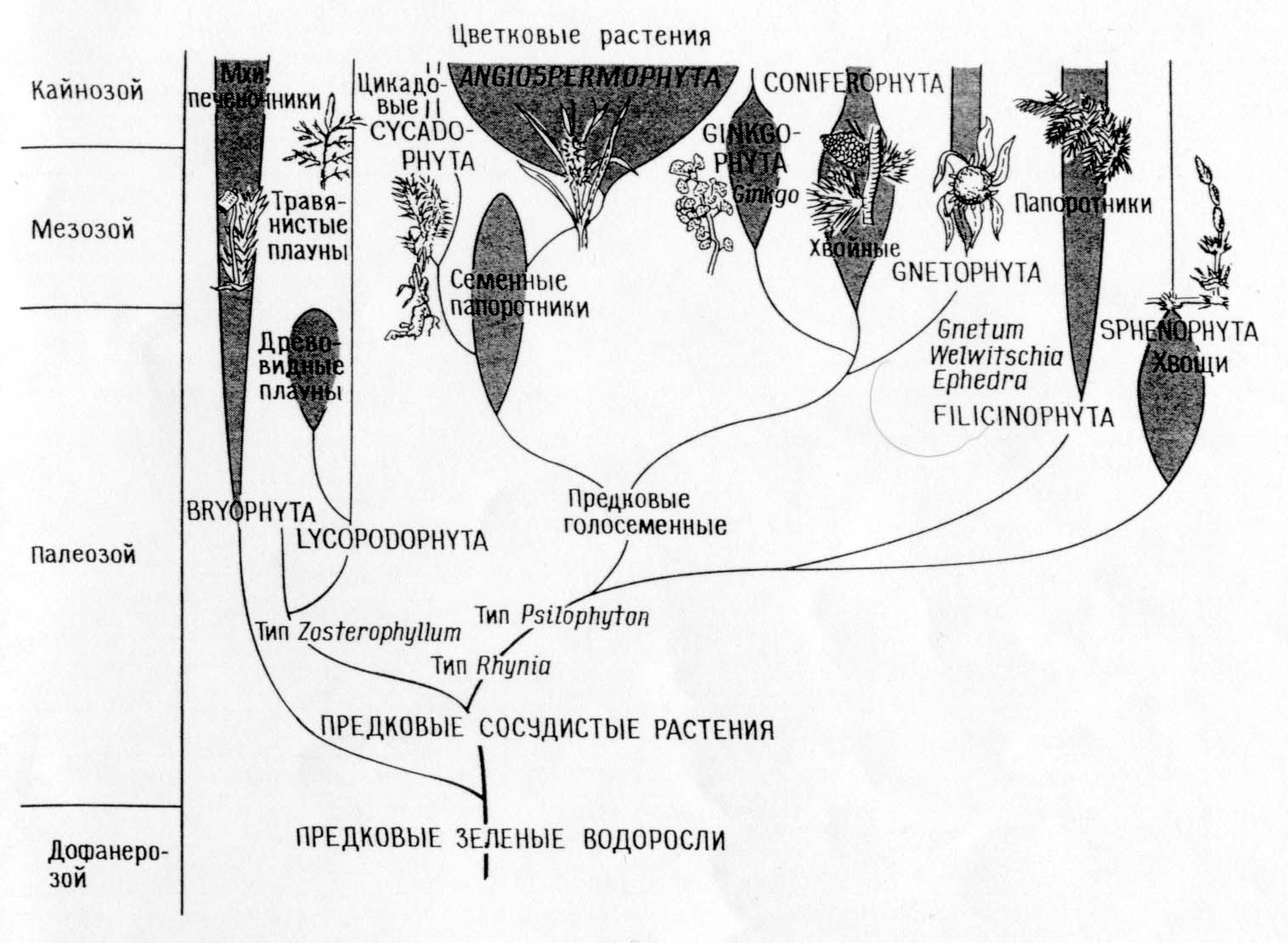
Filogenijų, atitinkančių tiesioginės giminės teoriją, palyginimas (A)
ir simbiozės teorija (B)
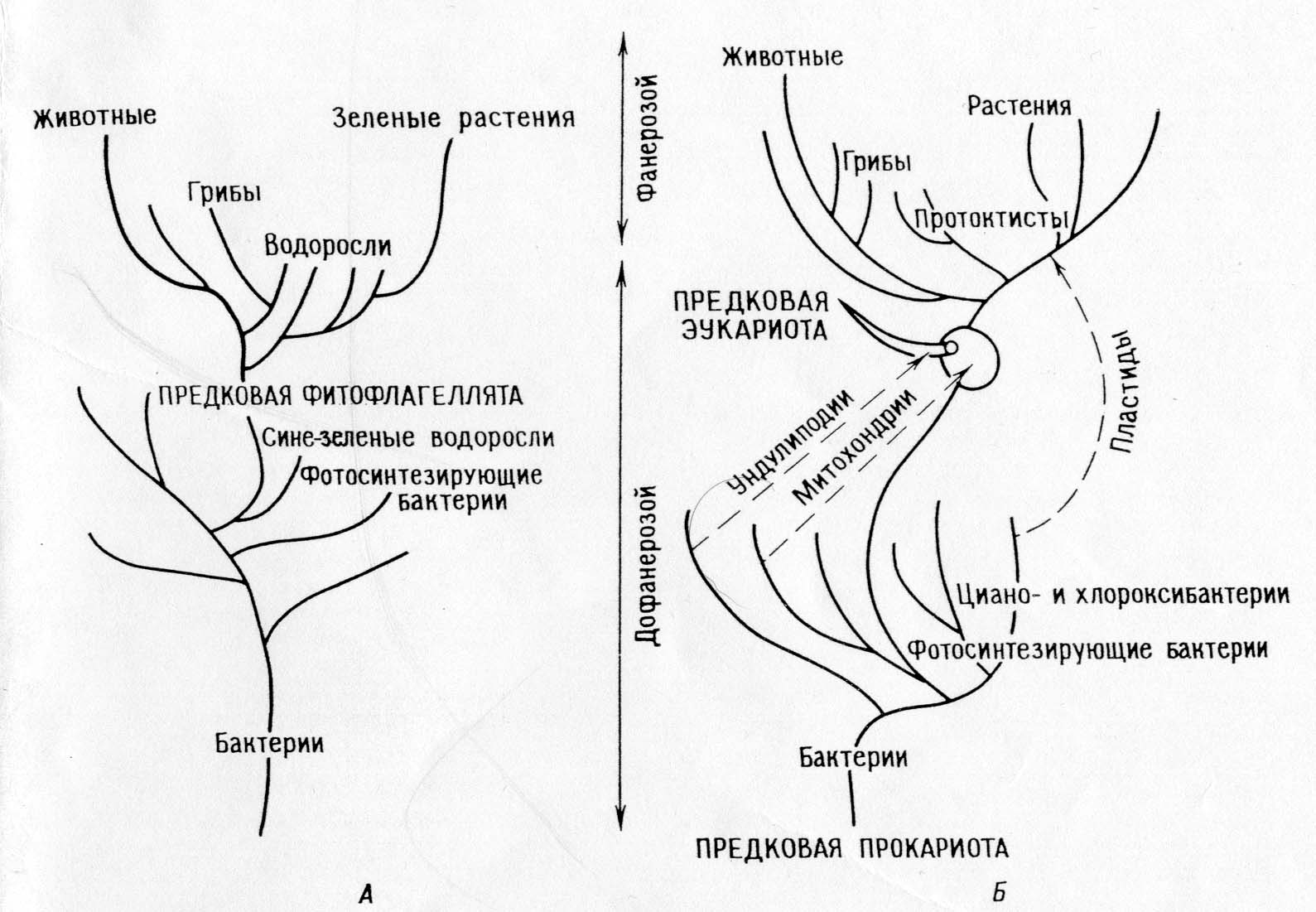
Skirtumai tarp tiesioginės giminės teorijos ir simbiotinės teorijos:
Tiesioginės giminės teorija.
Pagrindinė dichotomija: gyvūnai – augalai
Fotosintetiniai eukariotai (dumbliai ir žalieji augalai) išsivystė iš fotosintetinių prokariotų (mėlynadumblių)
Augalai ir jų fotosintezės sistemos išsivystė monofiletiškai Prekambrijoje.
Gyvūnai ir grybai išsivystė iš autotrofų, kurie prarado plastidus.
Mitochondrijos tarpląsteliniu būdu diferencijuojasi fotosintetiniuose dumbliuose.
Dešiniosiose piktžolėse undolipodijos, mitozinis aparatas ir visi kiti eukariotų organeliai diferencijuojasi tarpląsteliniu būdu.
Visi organizmai kilę iš savo protėvių – prokariotų, susikaupę pavienių mutacijų.
PASEKMĖ: Pagal teoriją turi būti dešiniosios piktžolės; sunku susieti šią teoriją su iškastiniais įrašais ir sukurti atitinkamą filogeniją.
Viena po kitos einančių endosimbiozių teorija:
Pagrindinė dichotomija: prokariotai – eukariotai.
Fotosintetiniai eukariotai (branduoliniai dumbliai ir augalai) ir nefotosintetiniai eukariotai (gyvūnai, grybai, pirmuonys) išsivystė iš bendrų heterotrofinių protėvių (ameboflagelatų).
Fotosintezės evoliucija Archėjuje buvo monofilinė bakterijose (įskaitant ciano ir chloroksibakterijas), fotosintetiniai protistai staiga atsirado, kai jų heterotrofiniai protistai-protėviai įsitraukė į simbiozę su fotosintetiniais prokariotais (protoplastidais).
Gyvūnai ir dauguma dumblių bei grybų yra tiesiogiai kilę iš heterotrofinių protistų.
Mitochondrijos jau buvo heterotrofiniuose protistuose, kai pastarieji dėl simbiozės įgijo plastides. Promitochondrijos anksčiau buvo įgytos simbiotiniu būdu.
Mitozė išsivystė heterotrofiniams eukariotų protistams diferencijuojant undolipodinę sistemą.
Prokariotai išsivystė iš ankstyvųjų prokariotų protėvių per keletą pavienių mutacijų įvykių, eukariotai taip pat išsivystė iš eukariotų protėvių per pavienius mutacijos įvykius, o eukariotų protėviai išsivystė iš prokariotų per daugybę simbiozių.
Pasekmės: teorija paaiškina biocheminius kelius ir jų perteklius organelėse. Tai leidžia sukurti nuoseklią filogeniją, paaiškinančią „biologinį atotrūkį“ tarp proterozojaus ir fanerozojaus laikų. Teorija numato, kad organelės turi išlaikyti replikacijos sistemų likučius.
botanikos mitas
Nuo Jussieu laikų žemesni augalai vadinami talofitais, o aukštesni – briofitais ir tracheofitais. Remiantis botanikos mitais, bendri fotosintetinių bakterijų ir melsvadumblių protėviai dėl tiesioginės giminės sukėlė eukariotinių dumblių, įskaitant jūrinius, pradžią. Organizmų grupės, kurių filogeniją labiausiai veikia tiesioginės giminės sąvoka, atsidūrė botanikos srityje – tai bakterijos, dumbliai, grybai ir augalai.
Hipotetiniai organizmai, galintys būti jungtimi tarp melsvadumblių ir žaliųjų dumblių, buvo vadinami protėvių fitomonadais (protėvių fitomonadais) arba dešiniosiomis piktžolėmis. Kai kurie autoriai teigė, kad Cyanidium ir Cyanophora eukariotai, kurių pigmentų sistemos yra panašios į cianobakterijas, gali būti tinkami kandidatai į dešiniųjų piktžolių vaidmenį. Talofituose nerasta tarpinių formų tarp nemitozinės melsvadumblių būklės ir visiškai išsivysčiusios tipinės žaliųjų dumblių ir augalų mitozės. Nors grybai ir žydintys augalai visiškai neturi undulipodijų, jiems būdinga mitozė ir mejozė, kurioms reikalingi mikrovamzdeliai. Visuotinai sutariama, kad visi daugialąsčiai organizmai, turintys mikrovamzdelius, yra kilę iš eukariotinių mikroorganizmų su undolipodijomis, tačiau niekas nežino, iš kurios protistų grupės. Botanikos mitą sustiprino fotosintezės senumas ir sudėtingumas, taip pat tai, kad daugelis organizmų (bakterijų, euglenoidų, parazitinių augalų) tiek gamtoje, tiek laboratorijoje dėl mutacijų prarado fotosintezę. Kadangi fotosintezė yra anaerobinis procesas, galiausiai užtikrinantis visų organizmų egzistavimą, ji turėjo išsivystyti labai anksti mūsų planetos gyvybės istorijoje, dar prieš atsirandant mitozei, kuriai reikalingas deguonis. fotosintezėje dalyvauja daug pigmentų, lipidų ir fermentų, kurie tvarkingai išsidėstę labai sudėtingose membranose. Tuo pačiu metu dumblių ir augalų fotosintezės mechanizmų detalės, viena vertus, ir cianobakterijos, iš kitos pusės, yra stebėtinai panašios. Visi šie organizmai iš vandens išskiria deguonies dujas, kurios yra vandenilio atomų, naudojamų CO2 redukavimui į ląstelės organines medžiagas, šaltinis. Daugelis dumblių ir pirmuonių neturi nei mitozės, nei genetinės sistemos, panašios į gyvūnų ir augalų. Blakstienos turi specialią genetinę sistemą su 2 tipų branduoliais, o ląstelės dažnai būna daugiabranduolės. Vieno tipo branduoliai yra makrobranduoliai, o kiti – daug mažesni mikrobranduoliai. Mikrobranduoliai yra fiziologiškai nereikšmingi. tačiau jie saugo genų kopijas ir lytinio proceso metu patiria mejozę. Fiziologiškai esminiai makrobranduoliai yra RNR transkripcijos vieta ir taip valdo ląstelių fiziologiją. Vienas iš sunkumų išaiškinant ryšį tarp melsvadumblių ir eukariotinių dumblių yra duomenų apie undolipodijų kilmę trūkumas: tarp cianobakterijų, kurios niekada neturi undolipodijų, ir daugelio dumblių, kurios turi, nerasta. Kita problema – visiškas melsvadumblių lytinio proceso nebuvimas: tarp nelytinių melsvadumblių ir dumblių, kurių daugumai būdingas lytinis procesas, tarpinių formų nebuvo. Nustatyta, kad dauguma dumblių tam tikru gyvenimo ciklo etapu turi undolipodijų. Dumblių, pirmuonių, gyvūnų ir augalų judrios struktūros pasirodė esančios vienodos: jos visos susideda iš 0,024 mikrono skersmens mikrovamzdelių. Paaiškėjo, kad mitozinė verpstė, net ir grybuose, kurie neturi undolipdijų, susideda iš tokio pat skersmens baltymų mikrotubulių, kaip ir undolipdijų. Pusę amžiaus atrodė labai tikėtina, kad eukariotų dumbliai išsivystė iš melsvadumblių. Šis botaninis mitas lėmė prielaidas, kurios daugelį metų buvo ir stimulas tyrimams, ir atskaitos sistema, pagal kurią buvo vertinami rezultatai. Teiginys, kad ryšiai tarp cianobakterijų ir eukariotinių dumblių išnyko, nepalikdami fosilijų pėdsakų ir gyvų relikvijų, tapo vis mažiau įtikinami, nes atsirado naujų duomenų apie gyvus mikrobus ir fosilijas. Pagrindinės tarpinės grandys tarp fotosintetinių prokariotų ir eukariotų neišnyko be pėdsakų, kaip reikalauja botanikos mitas – jų niekada nebuvo. Remiantis nuoseklių endosimbiozių teorija, cianobakterijos iš tikrųjų yra protėviai, bet tik plastidai, o ne likusi eukariotinės ląstelės dalis ir tikrai ne jos branduolys. Ir vis dėlto, sprendžiant branduolio kilmės klausimą, tiesioginės giminės hipotezė ir simbiozės teorija dera. Įmanomos ekstremalios simbiozės teorijos versijos modifikacijos apima, pavyzdžiui, plastidžių, bet ne mitochondrijų, simbiotinės kilmės idėją arba plastidų ir mitochondrijų simbiotinę kilmę, bet ne undolipodiją. Tiesioginės giminės šalininkai taip pat pateikia prielaidą apie branduolio kilmę per simbiozę.
Kritiniai chloroplastų ir mitochondrijų endosimbiotinės kilmės įrodymai.
Reikšmingiausi įrodymai apie svarbiausių eukariotinių ląstelių organelių – chloroplastų ir mitochondrijų kilmę iš prokariotų gauti pastaraisiais metais tiriant ribosomų RNR struktūrą. Molekulinis biologas
K. Wozas jau devintajame dešimtmetyje parodė, kad RNR iš augalų chloroplastų ribosomų struktūra yra beveik identiška kai kurių cianobakterijų RNR struktūrai. Tuo pačiu metu grupė mokslininkų nustatė, kad ribosomų RNR, gauta iš skirtingų eukariotų mitochondrijų, yra panaši į kai kurių bakterijų, ypač Paracoccus genties bakterijų, RNR. Šios bakterijos kvėpavimo fermentų rinkinys labai panašus į
gyvūnų mitochondrijų „kvėpavimo takų ansamblis“.
Šie duomenys gana aiškiai liudija didesnę eukariotinių ląstelių endosimbiotinės kilmės tikimybę. Endosimbiozės reiškinys vis dar itin plačiai paplitęs gamtoje. Yra žinoma, kad ant kai kurių jūrų žuvų kūno dažnai nusėda šviečiančios bakterijos, kurios ne tik labai praverčia savo gyvybei, bet ir yra labai naudingos šeimininkams. Visa tai, kas išdėstyta aukščiau, įtikinamai liudija, kad eukariotinės ląstelės yra endosimbiotinės kilmės iš daug mažesnių prokariotinių ląstelių. Tačiau pagrindinis klausimas yra nustatyti ląstelės šeimininkės prigimtį, nes eukariotinės ląstelės turi gerai suformuotą branduolį su apvalkalu, o prokariotai tokio branduolio neturi. Pavyzdžiui, L. Margelis mano, kad ląstelių šeimininkų vaidmenį galėjo atlikti prokariotai – šiuolaikinių bakterijų pirmtakai, priklausantys mikoplazmoms – labai primityviai sukonstruoti prokariotai, praktiškai neturintys ląstelių sienelių. Pastaruoju metu pasirodė ir plačiai aptarinėjami japonų mokslininko darbai.
T. Oshima. Remdamasis nuodugniais daugelio pro- ir eukariotų ribosomų RNR struktūros tyrimais, įskaitant daugelį iki šiol mažai ištirtos archajų superkaralystės atstovų, šis autorius daro išvadą, kad ląstelė šeimininkė buvo viena iš šiuolaikinės archejos protėvių. Reikia pasakyti, kad šios superkaralystės atstovai, skirtingai nei bakterijos, turi daug biocheminių ir molekulinių biologinių savybių, bendrų su eukariotais. Naujausiuose T. Oshimos darbuose pateikiami argumentai už tai, kad ląstelė šeimininkė eukariotų atsiradimo metu buvo archėjos protėvis, panašus į vieną iš šiuolaikinių šių prokariotų atstovų – termoplazmą. Koks bus galutinis problemos, susijusios su šeimininko ląstelės, kuri sukėlė eukariotinę ląstelę daugybinės endosimbiozės procese, nustatymo problemos sprendimas, parodys tolesni tyrimai.
Šiuo metu pateikiama populiariausia ir pagrįsta eukariotinių ląstelių kilmės hipotezė
įvairių prokariotų, priklausančių bakterijoms ir archejoms, endosimbiozė, daug kartų pasireiškusi evoliucijos procese per pirmuosius 1,5–2 milijardus gyvų ląstelių egzistavimo Žemėje metų. Lemiamų įrodymų apie endosimbiotinę chloroplastų ir mitochondrijų kilmę gavo pirmaujantys pasaulio biologai, tirdami ribosomų RNR struktūrą. Šie duomenys gana aiškiai liudija didelę tikimybę, kad eukariotinės ląstelės bus endosimbiotinės kilmės iš daug mažesnių (keliais dydžiais) prokariotinių ląstelių.
LITERATŪRA
Soroso edukacinis žurnalas. Biologija, chemija, geomokslai, fizika, matematika. N 5 1998
Margelis L. Simbiozės vaidmuo ląstelių evoliucijoje. Per. iš anglų kalbos, M. MIR 1983 m
Iš pagrindinių kinetikos skyrių jis tiria biologinių reakcijų kinetiką; Šis skyrius vadinamas biokinetika. Biokinetika yra paribių mokslas, atsiradęs biochemijos ir cheminės kinetikos sankirtoje (žr. 1 pav.). Biokinetikos, kaip atskiros disciplinos, pasirinkimas nėra atsitiktinis, logiškai pagrįstas ir siejamas su išskirtine kinetinių procesų svarba visiems gyviems organizmams. Biokinetika...
Genai, bet ir sukūrė prielaidas juose kauptis pokyčiams, genų divergencijai, jais valdomų produktų įvairovei didėti. 2. Mobilieji genetiniai elementai Tam tikras vaidmuo tiek pro-, tiek eukariotinių ląstelių genomų evoliucijoje tenka vadinamiesiems mobiliesiems genetiniams elementams – transpozonams. Jie yra savarankiški vienetai, turintys nukleotidų ...
Keletas galimi požiūriai ateities vystymuisi Bendroji teorija Dinaminės sistemos. 10 skyriuje „Žiūriniai gyvenimo procesai“ nagrinėjame būdingą ciklišką saviorganizavimosi organizavimą. dissipacinės sistemos. Apibendrinta schema pateikia transformavimo ciklų ir katalizinių reakcijų ciklų, taip pat katalizinių hiperciklų generavimo charakteristikas pagal...