Filogeneza, filogeneza - rozwój historycznyświat organizmów żywych, zarówno w ogólnych, jak i indywidualnych grupach taksonomicznych: królestwach, typach (podziałach), klasach, rzędach (rzędach), rodzinach, rodzajach, gatunkach. Termin filogeneza wprowadził E. Haeckel.
Współczesny system zwierząt, zbudowany na zasadach pokrewieństwa, można przedstawić jako drzewo genealogiczne (lub filogenetyczne), odzwierciedlające główne etapy ewolucji zwierząt. Filogeneza różne grupy organizmy badane nierównomiernie. Filogeneza kręgowców (zwłaszcza wyższych grup) została najlepiej zbadana; wśród bezkręgowców najlepiej zbadana została filogeneza mięczaków, szkarłupni i stawonogów. Filogeneza prokariotów jest słabo poznana.
U podstawy drzewa ewolucyjnego zwierząt znajdują się zwierzęta jednokomórkowe. W organizmach jednokomórkowych wszystkie funkcje organizmu są wykonywane przez jedną komórkę. Istnieje kilka hipotez na temat pochodzenia jednokomórkowe eukarionty s: 1) eukarionty wyewoluowały z prokariotów w wyniku symbiogenezy. Hipotezę tę potwierdza fakt, że eukariotyczne mitochondria i chloroplasty mają własne DNA; 2) eukarionty powstały w wyniku różnicowania się komórek prokariotycznych w cytoplazmę i organelle. Wszystkie typy współczesnych pierwotniaków wywodzą się od starożytnych sarkoflagellatów.
Wielokomórkowe, lepiej zorganizowane zwierzęta, w których poszczególne funkcje organizmy są wykonywane przez pojedyncze komórki lub narządy. Wszystkie organizmy wielokomórkowe w indywidualnym rozwoju zaczynają się od pojedynczej komórki - zygoty i początkowe etapy zarodek (blastula) przypomina jednokomórkową kolonię.
Istnieje wiele hipotez dotyczących pochodzenia zwierząt wielokomórkowych. Najbardziej znane są hipotezy E. Haeckela i I.I. Miecznikow. Zgodnie z teorią Haeckela wszystkie zwierzęta wielokomórkowe wywodzą się od jednej formy przodków - gastrea. I.I. Miecznikow założył, że przodkiem zwierząt wielokomórkowych była kolonia wiciowców. Pierwszy organizm wielokomórkowy miał kulisty kształt, był pokryty warstwą komórek z wici i miał wewnątrz ciała komórki ameboidalne przewodu pokarmowego, swobodnie unoszące się w wodzie. I.I. Miecznikow nazwał to fagocytellą. Dalsza ewolucja organizmów wielokomórkowych przebiegała w czterech kierunkach. Jeden z kierunków ewolucji doprowadził do powstania typu blaszkowatego, składającego się z pojedynczej warstwy komórek i prowadzącego pełzający tryb życia. Inny kierunek ewolucji doprowadził do powstania typu gąbczastego z dwiema warstwami komórek nie uformowanych w tkankę. Nie mają jeszcze układu nerwowego. Są to nieruchome zwierzęta bentosowe. Trzeci kierunek ewolucji doprowadził do powstania Typu Coelenterates. Koelenteraty są już prawdziwymi zwierzętami dwuwarstwowymi (ektoderma i endoderma) z jelitem pierwotnym, ustami, symetrią promieniową, rozproszonym układem nerwowym. Wśród nich są zwierzęta swobodnie pływające i prowadzące przywiązany tryb życia. Czwarty kierunek ewolucji doprowadził do powstania jeszcze lepiej zorganizowanych zwierząt trójwarstwowych. Tkanki i narządy tych zwierząt powstały z trzech warstw komórek (listków zarodkowych): ektodermy, endodermy i mezodermy.
Pierwszymi trójwarstwowymi były zwierzęta podobne do planarian jelitowych. Ewolucja zwierząt trójwarstwowych przebiegała w trzech kierunkach. Jeden kierunek doprowadził do powstania płazińce, w którym nie ma jamy ciała, a miąższ znajduje się między narządami. Płazińce są pierwszymi zwierzętami dwustronnymi (wszystkie kolejne typy mają tę symetrię). Pojawiają się jako pierwsi system mięśniowy(gładki), zwoje nerwowe. Jednak uformowane jelito jest ślepo zamknięte, nie ma odbytu.
Inny kierunek doprowadził do powstania pierwotnych zwierząt jamistych glisty. Ich jelita stają się od końca do końca, składają się z trzech odcinków z odbyt. Jest dalsza koncentracja komórki nerwowe, tworząc pierścień nerwu okołogardłowego. Wspólną cechą płazińców i pierścienic jest obecność worka skórno-mięśniowego, układu pokarmowego, wydalniczego i rozrodczego.
Trzeci kierunek ewolucji trójwarstwowej prowadzi do powstania zwierząt deuterowanych. Utworzenie wtórnej jamy ciała zapewniło większą mobilność i stabilność środowisko wewnętrzne organizm. Wnęki wtórne są lepiej zorganizowane: mają układ krążenia. Należą do nich wszystkie inne bezkręgowce i akordy.
Ewolucja deuterostomów doprowadziła do powstania protostomów i deuterostomów. Ewolucja protostomów. Mięczaki i pierwotne pierścienice pochodzą od przodka protostomów. Z kolei współczesne pierścienice i stawonogi wywodzą się z pierścienic pierwotnych.
Pierścienie mają postępujące cechy organizacyjne: wtórną jamę ciała, metamerycznie wypreparowane ciało, zamknięty układ krążenia (niezamknięty u innych bezkręgowców), pierścień okołogardłowy i łańcuch nerwowy.
Stawonogi osiągają większą doskonałość: serce powstaje w układzie krążenia; w układzie nerwowym wzrasta koncentracja zwojów; uformowany egzoszkielet(osłona chitynowa); charakterystyczna jest różnorodność narządów oddechowych (skrzela, tchawica, płuca); tworzą się połączone kończyny i skrzydła. Bardziej doskonała budowa stawonogów pozwoliła im na szerokie zaludnienie Ziemi.
Mięczaki mają wtórną jamę ciała, złożoną narządy wewnętrzne(tworzą się nerki, trójkomorowe serce, skrzela, płuca), większość ma wapienną skorupę, która chroni ciało. Jednak u mięczaków układ krążenia nie jest zamknięty, a układ nerwowy ma rozproszony typ guzkowy.
Ewolucja Deuterostomu. Szkarłupnie i strunowce wywodzą się od przodków deuterostomów.
Szkarłupnie. Charakteryzują się: wtórną jamą ciała, układ krążenia tworzy szczeliny, rodzajem układu motorycznego (ambulakralnego), symetrią promienistą, układem nerwowym w postaci pierścienia z promieniowymi promieniami.
Pierwotne akordy prawdopodobnie wyewoluowały z deuterostomicznych przodków przypominających robaki. Izolacja akordów wiąże się ze wzrostem ruchomości. O pokrewieństwie filogenetycznym strunowców z bezkręgowcami świadczą: 1) tkanki i narządy zbudowane są z trzech listków zarodkowych (powstałych po raz pierwszy u płazińców); 2) dwustronna symetria (występuje u płazińców); 3) wtórna jama ciała (występuje w pierścieniach); 4) wtórne usta (spokrewnione ze szkarłupniami); 5) zamknięty układ krążenia (pierścienie);
Z pierwotnych strunowców wywodzą się współczesne zwierzęta nieczaszkowe (lancetowate) i kręgowce. Struny larwalne są boczną gałęzią strunowców, które w większości przechodzą do osiadłego cyklu życiowego, dlatego ślady strunowców mają dopiero w stadium larwalnym.
Cephalothordates to prymitywne akordy. Mają nephridia, podobne do metanephridia pierścienic, zamknięty układ krążenia bez serca, jak pierścienice.
Przodkowie kręgowców pojawili się około 500 milionów lat temu. Ich ewolucja przebiegała w dwóch kierunkach: bezszczękowy (cyklostomy) i szczękowy (wszystkie inne kręgowce).
Pierwsza szczękowa - opancerzona ryba wyewoluowała z opancerzonej bezszczękowej. Mieli już moc szkielet wewnętrzny, czaszka mózgowa i trzewna, szczęki z zębami, sparowane płetwy. Te ryby są wymarłe. Zostały one zastąpione przez chrzęstne i oścista ryba którego ciało było pokryte łuskami.
Ryby chrzęstnoszkieletowe były powszechne w morzach i ze względu na rodzaj pożywienia były drapieżnikami.
Ryby kostnoszkieletowe pierwotnie pojawiały się w wodach słodkich. Wykształciły wieczko (chroni skrzela przed zatykaniem) i pęcherz pławny (ułatwia pływanie w słodkiej wodzie). W wysychających zbiornikach wodnych (w paleozoiku) pojawiły się ryby płetwiaste (płetwiaste i płetwiaste), w których oprócz skrzeli rozwinęły się worki płucne do oddychania powietrzem, a mięśnie uformowane na płetwach umożliwiały pochylanie się na twardym podłożu. Następnie ryby kostnoszkieletowe zamieszkiwały wody słodkie i morskie. Obecnie ryby kostnoszkieletowe są jedną z najlepiej prosperujących grup kręgowców (20 tys. gatunków).
Płazy pojawiły się w dewonie. Płazy wywodzą się od starożytnych ryb płetwiastych. Pierwszymi płazami były Ichthyostegidae (czworonogi). Oni, podobnie jak ryby, mieli płetwę ogonową, osłonę skrzelową, łuski. Ale w przeciwieństwie do ryb miały dwie pary pięciopalczastych kończyn i płuc. Żyli głównie w wodzie.
Z ichtiostega pochodzą prawdziwe płazy - stegocefale (muszlowate). Głowa stegocefalów była pokryta skorupą kości skórnych. Stegocefalianie żyli do początku okresu kredowego. Prawdopodobnie dały one początek współczesnym płazom bez pancerza i pierwszym gadom.
Lądowaniu towarzyszyła aromorfoza: oddychanie skrzelowe zmieniło się w oddychanie płucne, a płetwy w pięciopalczaste kończyny, serce stało się trójkomorowe, powstał drugi krąg krążenia, zmieniły się narządy zmysłów, a układ nerwowy stał się doskonalszy.
Pierwsze gady pojawiły się w karbonie. Skamieniałe szczątki Seymourii miały cechy płazów i gadów. Jest prawdopodobne, że pierwsze gady, kotylozaury, wywodzą się z takich form. Mieli też czaszkę w formie kościanej skrzyni (podobnej do stegocefalów) z otworami na oczy, nozdrza i narząd ciemieniowy. Okres rozkwitu gadów przypadał na mezozoik.
Występuje: rogowacenie naskórka i tworzenie się łusek lub łusek (chroni organizm przed wysychaniem), powikłanie budowy płuc, serca, poprawa kośćca, tworzenie się błon rozrodczych surowiczego i owodniowy (pod warunkiem zdolności do rozmnażania się na lądzie).
Gady dały początek dwóm nowym klasom kręgowców: ssakom i ptakom.
Ptaki wywodzą się od pierwszych ptaków z grupy biegających dinozaurów jaszczurek tylne nogi. Spośród starożytnych pierwszych ptaków znaleziono szczątki archaeopteryksa długoogoniastego. Archaeopteryx nosił znamiona gadów (długi ogon, zęby na szczękach, pazury na skrzydłach) i ptaków (pióra, skrzydła, wygląd). Prawdziwe ptaki pojawiły się od pierwszych ptaków pod koniec mezozoiku. Mieli też małe zęby.
Cechy progresywne ptaków: stałocieplność, serce czterokomorowe, jeden łuk aorty (powoduje całkowite oddzielenie krążenia systemowego i płucnego).
Ssaki pojawiły się wcześniej niż ptaki. Przypuszcza się, że pochodziły one od jaszczurek zwierzęcozębnych, które posiadały szereg cech progresywnych: zęby znajdują się w pęcherzykach płucnych, kości wtórnej podniebienia, kończyny znajdowały się pod tułowiem, a nie po bokach, występowały zmiany w szkielecie. Cynodonty były najbliżej ssaków spośród gadów ssakozębnych. Dalsza ewolucja ssaków wiąże się ze zmianami w systemie uzębienia. Zmiany te doprowadziły do powstania dwóch grup zwierząt. Potomkowie jednej grupy - wielogruźlicze byli małymi zwierzętami, mieli liczne guzki na zębach trzonowych, silnie rozwinięte siekacze i nie mieli kłów. Możliwe, że ta grupa dała początek monotremom.
Druga grupa rozwijała się bardziej progresywnie. To były małe zwierzęta, które zjadały owady, małe zwierzęta. W przyszłości ta grupa zostanie podzielona na dwie niezależne gałęzie - Dolną, Torbaczową i Wyższą, Łożyskową.
Torbacze powstały w okresie kredy. Najstarszą grupą torbaczy jest rodzina oposów.
Ssaki łożyskowe pojawiły się także w okresie kredowym. Za najstarszą grupę łożyskowców uważa się grupę owadożerców. Przypuszcza się, że dały one początek większości późniejszych grup łożyskowych. Możliwe, że naczelne pochodzą bezpośrednio od owadożerców.
Postępująca ewolucja ssaków rasy Stlan z nabyciem najważniejszych przystosowań: stałocieplność, wysoka przemiana materii, tworzenie łożyska, karmienie młodych mlekiem, opieka nad potomstwem. Sprzyjały temu postępujące zmiany w układzie oddechowym, krążeniowym, układy nerwowe. Ssaki mają czterokomorowe serce, jeden łuk aorty, tętnicę i krew żylna, płuca o strukturze pęcherzykowej, wtórne sklepienie nerwowe - neopalium jest dobrze rozwinięte.
13… Budowa komórki prokariotycznej. Wzrost, rozwój, reprodukcja bakterii. Rodzaje żywności bakterii. Fotosynteza, chemosynteza, fotoredukcja. Wartość mikroorganizmów w przyrodzie i gospodarce narodowej komórki prokariotyczne obejmują bakterie i niebiesko-zielone algi. Na prokarioty substancja jądrowa nie ma błony jądrowej; ich błona plazmatyczna zawiera węglowodany i aminokwasy. Prokarioty mają tylko jeden chromosom, który składa się wyłącznie z DNA. Ten chromosom ma kształt pierścienia i nie zawiera białek zasadowych - histonów. Zestaw chromosomów u prokariotów jest haploidalny.Te organizmy jednokomórkowe nie mają mitozy i nie mają aparatu mitotycznego. Prokarionty nie mają mejozy, a podczas procesu płciowego przekazywana jest tylko część informacji dziedzicznej, a sam organizm jest gametą. Cytoplazmatyczny DNA u prokariotów jest zlokalizowany w plazmidach, a błony cytoplazmatyczne są nieobecne. Ściana komórkowa prokariotów zawiera peptydoglikany. U prokariotów nie obserwuje się mitochondriów, enzymy oddechowe są zlokalizowane w mezosomach, które znajdują się na wewnętrznej powierzchni błony plazmatycznej.Procesy fotosyntezy przeprowadzane są przy użyciu pigmentu bakteriochlorofilowego, a związki siarki służą jako czynniki redukujące. Rybosomy prokariotyczne są mniejsze niż rybosomy eukariotyczne. Nie ma ruchu cytoplazmy, wakuole są w niej bardzo rzadkie. Wici prokariotów są wielkości submikroskopowych, wici zawierają jedną lub więcej mikrotubul (fibryli).Prokarioty są wrażliwe na antybiotyki i są odporne na promieniowanie rentgenowskie i podniesiona temperatura, wytrzymujące nagrzewanie do + 75 ... + 90 ° С. charakterystyczna cecha prokariontów polega na tym, że nie są one zdolne do współistnienia z innymi typami komórek. bakteria- Jednokomórkowe organizmy; można je zobaczyć tylko pod mikroskopem. Dlatego nazywane są mikrobami lub mikroorganizmami. Bakterie opanowały różnorodne siedliska: żyją w glebie, kurzu, wodzie, powietrzu, na zewnętrznych powłokach zwierząt i roślin oraz wewnątrz ciała. Można je znaleźć nawet w gorących źródłach, gdzie żyją w temperaturze około 60°C lub wyższej. Aktywność mikroorganizmów jest znaczenie dla wszystkich innych istot żywych, ponieważ bakterie i grzyby niszczą materię organiczną i uczestniczą w obiegu substancji w przyrodzie. Ponadto bakterie nabywają wszystko większa wartość w życiu ludzi, i to nie dlatego, że powodują różne choroby, ale dlatego, że można z nich uzyskać wiele potrzebnych produktów.
STRUKTURA KOMÓRKOWA. Prokariotyczny: brak uformowanego jądra, mitochondria, plastydy. W centrum cytoplazmy znajduje się nukleoid zawierający jeden „chromosom” z kolistą cząsteczką DNA; nie jest oddzielony od cytoplazmy błoną. Wewnątrz komórki wyrostki błony pełnią funkcje EPS, aparatu Golgiego; mezosom bierze udział w oddychaniu; są rybosomy. Cytoplazma jest pokryta błoną zewnętrzną i gęstą ścianą komórkową. Czasami komórka jest zanurzona w półpłynnej kapsułce koloidalnej. Ze względu na budowę ściany komórkowej bakterie można podzielić na dwie grupy: jedne są barwione metodą Grama, dlatego nazywane są gram-dodatnimi, inne zaś stają się bezbarwne po wypłukaniu barwnika i dlatego nazywane są gram-ujemnymi. Wzrost i rozmnażanie.Indywidualny wzrost i rozmnażanie bezpłciowe komórek. Stosunek powierzchni do objętości komórek bakteryjnych jest bardzo wysoki. Przyczynia się szybkie przejęcie składniki odżywcze z środowisko poprzez dyfuzję i transport aktywny. W korzystne warunki bakterie rozmnażają się bardzo szybko. Wzrost zależy przede wszystkim od temperatury i pH pożywki, dostępności składników odżywczych i stężenia jonów. Obowiązkowe tlenowce również potrzebują tlenu, a bezwzględne beztlenowce wręcz przeciwnie, potrzebują go całkowicie nieobecnego. Po osiągnięciu określonego rozmiaru bakterie przenoszą się do rozmnażanie bezpłciowe(rozszczepienie binarne), tj. zaczynają się dzielić z utworzeniem dwóch komórek potomnych. Przejście do podziału jest podyktowane stosunkiem objętości jądra do objętości cytoplazmy. Przed podziałem komórki następuje replikacja DNA, podczas której mezosomy utrzymują genom w określonej pozycji.Mezosomy mogą również przyczepiać się do nowych podziałów między komórkami potomnymi i w jakiś sposób uczestniczyć w syntezie substancji ściany komórkowej. Najszybciej rosnące bakterie dzielą się co 20 minut.
4 rodzaje odżywiania się bakterii i niektóre ich cechy.
Znane są trzy główne grupy Fotosyntetyczny(bakterie czerpiące energię ze słońca): 1. Bakterie zielonej siarki . Bakterie beztlenowe, które wykorzystują siarkowodór (H2S) i inne zredukowane związki siarki jako donory wodoru i elektronów.
Przykład: światło
2H2S + CO2 →→→→→→→→ [CH2O] + 2S + H2O.
bakteriochlorofil
W takim przypadku osadza się siarka.
2. Purpurowe bakterie siarkowe. Komórki mają ciemnoczerwony kolor, ponieważ jest znacznie mniej bakteriochlorofilu niż czerwonych i brązowych pigmentów. Są to głównie beztlenowce; fotosynteza w nich zachodzi w taki sam sposób jak w poprzedniej grupie.
3. Fioletowe bakterie bezsiarkowe . Dla tych bakterii różne związki organiczne służą jako źródło wodoru, który jest wykorzystywany do redukcji CO2 (w fotoautotrofach) lub niektórych związków organicznych (w fotoheterotrofach).
Chemosyntetyczny organizmy (chemoautotrofy) to bakterie, które wykorzystują CO2 jako źródło węgla, ale pobierają energię nie ze słońca, ale za pomocą reakcje chemiczne. Energia może zostać uwolniona, powiedzmy, podczas utleniania wodoru, siarkowodoru, siarki, żelaza (II), amoniaku, azotynów i innych związków nieorganicznych. W bakteriach żelaza : Tlen
Fe2+ →→→→→Fe3+ + Energia;
W bezbarwnych bakteriach siarkowych : tlen lub
S→→→→→→→ SO42-+ Energia;
Siarczan Siarki
W warunkach beztlenowych niektóre gatunki wykorzystują azotany jako akceptory wodoru i przeprowadzają w ten sposób denitryfikację.
W bakteriach nitryfikacyjnych: Tlen
2NH4+→→→→→ NO-2 + Energia
Azotyn amoniaku
Wszystkie rozważane bakterie są tlenowe, ponieważ tlen służy jako akceptor elektronów (i wodoru).
Bakterie chemosyntetyczne odgrywają bardzo ważną rolę w biosferze; biorą udział głównie w obiegu azotu, a tym samym utrzymują żyzność gleby.
Fotoredukcja - dla zielonych i fioletowych serobacter., które nie wykorzystują H2O, ale H2S jako źródło
ZNACZENIE W PRZYRODZIE I DZIAŁALNOŚCI GOSPODARCZEJ CZŁOWIEKA.
bakterie fermentacji mlekowej(heterotroficzne saprotrofy), które rozkładają węglowodany: Pozytywny: przygotowywanie produktów kwasu mlekowego (twaróg, zsiadłe mleko, masło, śmietana), zakiszanie pasz, zakwas, ogórki kiszone i pomidory. Negatywny: zepsucie produktów.
Bakterie fermentacji octowej(heterotroficzne saprotrofy), które rozkładają węglowodany: Pozytywny: utlenianie alkoholu do kwas octowy, który służy do marynowania, konserwowania owoców i warzyw. Negatywny: zepsucie produktów.
bakterie gnilne(heterotroficzne saprotrofy), które rozkładają białka: Pozytywny: rola sanitarna - mineralizacja pozostałości organicznych. Negatywny: psucie się żywności; aby uniknąć gnicia, stosuje się suszenie, solenie, marynowanie, sterylizację, pasteryzację i cukier.
Bakterie guzkowe(heterotroficzne symbionty): bakterie brodawkowe rhizobium przenikają do korzeni roślin strączkowych (koniczyna, łubin, lucerna itp.) i wchodzą z nimi w symbiozę. W efekcie na korzeniach tworzą się guzki – guzki wypełnione bakteriami, które syntetyzują związki azotu z azotu atmosferycznego, dostępnego zarówno dla rośliny żywicielskiej, jak i dla innych roślin. To naturalne wzbogacenie gleby w nawozy azotowe jest brane pod uwagę przy opracowywaniu płodozmianów polowych, które koniecznie obejmują rośliny strączkowe.
14. Budowa komórki eukariotycznej. 3 warstwowa membranaRybosomykompleks GolgiegomitochondriaLizosomyChloroplastychromoplastyLeukoplastyCentrum komórkowe lub centrosomInkluzjewakuolecytoszkieletKaryoplazmająderko1. 2 3 4 5
15.Ontogeneza i jej okresy. wczesna embriogeneza. Rozwój bezpośredni i pośredni.(51)
Ontogeneza- indywidualny rozwój organizmu od urodzenia do śmierci. Będzie to trwało dla różnych organizacji, nie jest takie samo i może trwać od kilku umysłów przez setki lat (żółw). Ontogeneza jest ściśle związana z filogenezą, ale nie jest absolutnym powtórzeniem filogenezy. Pod wpływem środowiska ulega zmianom i może wystąpić zmienność mutacyjna. okresy ontogenezy:
1 preembrionalny
2. zarodkowy (embrionalny)
3. postembrionalny
W okresie 1 rozwój komórek płciowych i przygotowanie ich do dalszego rozwoju po zapłodnieniu (spermatogeneza i owogeneza) spermatogeneza z 4 okresów: 1. rozmnażanie; 2. wzrost; 3. dojrzewanie; 4. kształtowanie
Spermatogeneza: Spermatogonia 2n „spermatocyty pierwszego rzędu 2n” spermatocyty 2 pory 2n „plemniki n.
Owogeneza: O vogonia 2n "oocyty 1 por2n" oocyty 2 por2n "1 jajo + 3 ciałka kierunkowe (zredukowane)
W II okresie rozwój zarodka następował in utero (w łonie matki) lub w środowisku zewnętrznym. Har-Xia szybkie procesy kształtowania, do-rycia krótkoterminowe doprowadzić do powstania wielu org-ma. Ma 3 okresy: 1. inicjał- 1 tydzień rozwój zarodkowy rozpoczyna się od momentu zapłodnienia, a kończy wraz z zagnieżdżeniem się zarodka w błonie śluzowej matki. 2 zarodkowe. Czas od startu 2 do 8 tygodni od implantacji do zakończenia organogenezy. 3 płody- rozwój płodu od 9 tygodnia do porodu. Zwiększony wzrost, kształtowanie, odżywianie hematotroficzne.
3. okres post-ebr rozpoczyna się od momentu narodzin, tj. uwolnienie organizmu z zarodka muszli. W tym okresie rozwoju wykształciła się specyficzna cecha anatomiczna i funkcjonalna, tzw wiek. Periodyzacja:
1-10 dni - noworodek
11d - 1 rok niemowlęcia
1g-11 lat dzieci (dziewczęta i chłopcy)
11-15 lat (d) i 12-16 (m) - nastolatki
16-20 (d) i 17-21 (m) - młodzież
21-55 (w) i 22-60 (m) - dojrzałe
56, 61-74 - osoby starsze
75-90 starców, ponad 90 stulatków
Rozwój bezpośredni (nielarwalny), związany z > liczbą żółtek w jaju lub rozwojem org. W organizmie matki jest charakterystyczny dla ryb, ssaków, ptaków, gadów.
Rozwój pośredni - (larwa) z metamorfozą - charakter. d/ niekonsekwencja, skorupiaki, owady, robaki, charakter. Krótki okres embrionalny u kota. Powstaje larwa. Siebie, ale istnieć. Niepełna przemiana - jajo-larwa-imago. Całkowita przemiana - jajo - larwa - poczwarka - imago. WCZESNA EMBRIOGENEZA: plemnik jest wprowadzany do komórki jajowej, cytoplazma rozpuszcza się - stado syncarionu (jądra łączą się z „diploidalnym jądrem” zygotą 2n-div. Na blastomerach-4, 8,16,32,64,128 ..-arr. blastula (zarodek z jamą wewnątrz) - przez inwaginację arr. - dwuwarstwowy - gastrula, ect.ent) - (blastocoel, blastopor) - rozwinięty. 3 kiełki. Listok. endoderma, mezoderma, ektoderma
Kruszenie może być równomierne i nierówne (tarczowe)
Gastrulacja występuje na 4 sposoby:
1 przez wgłobienie (wgłobienie)
2 epibolia (zanieczyszczenie)
3 rozwarstwienie (rozwarstwienie)
4 imigracja
Neurulacja anlage cewy nerwowej, strun, fałdów owodniowych i tułowia
ektoderma tworzy płytkę nerwową-st. neurula (ułożenie rurki nerwowej, struny), słuch, węch, powłoka zewnętrzna.Endoderma jest początkiem jelit, wątroby, płuc, trzustki. Forma mezodermy. Mięśnie (dermotom, miotom), tkanka mięśniowa, układ krwionośny, wszystkie elementy chrzęstno-kostne szkieletu, prążki, układ płciowy.
16.Główne ścieżki filogenezy. Rozbieżność, zbieżność, równoległość.(52)
Rozwój ewolucyjny może iść różne sposoby w zależności od obiektu i jego relacji z otoczeniem. Istnieje kilka form filogenezy.
Ewolucja fitetyczna to stopniowe ewolucyjne przegrupowanie pewnej grupy taksonomicznej, które prowadzi do powstania nowej grupy, różniącej się od pierwotnej. Przykładem ewolucji filetycznej może być szereg przodków koni: eogippus - epigippus - meryk-hippus - gashogippus - koń współczesny.
Rozbieżność - główna ścieżka ewolucji, który był osadzony w podstawach teorii darwinowskiej. Rozbieżna ewolucja jest determinowana przez rozbieżność taksonu na kilka grup pod wpływem elementarnych czynników ewolucji i adaptacji do różne warunki istnienie. W przypadku dywergencji podobieństwo jest określane przez wspólne pochodzenie, a różnice są określane przez adaptację do innego środowiska. Początek dywergencji w obrębie gatunku podczas formowania się ekotypów, ras ekologicznych, podgatunków geograficznych. Gdy cechy się rozchodzą, odmiany stają się gatunkami, gatunki przechodzą dalsze różnicowanie i tworzą rodzaje, rodzaje stają się rodzinami i tak dalej.
Rozbieżność może przybierać różne formy: dychotomiczne rozgałęzienie (rozpad gatunku na dwa gatunki potomne), promieniowanie (raj upadek gatunku na kilka gatunków potomnych), rozszczepienie (istnienie przodka istnieje jednocześnie z podmiotami zależnymi, utworzonymi z izolowanych w inny czas populacje). Dywergencja rozwija się na podstawie narządów homologicznych. Klasyczny przykład rozbieżność można nazwać rozbieżnością cech u zięb archipelagu Galapagos.
Biologiczne znaczenie dywergencji polega na tym, że prowadzi ona do różnorodności organizmów żywych, do równomiernego rozmieszczenia gatunków w niszach ekologicznych i łagodzi napięcie walki o byt.
Konwergencja jest przeciwieństwem procesu dywergencji. Polega ona na samodzielnym nabyciu, w tych samych warunkach, podobnych cech w formach zasadniczo niezwiązanych ze sobą na podstawie podobnych narządów lub struktur. Na przykład, kopiące nory kończyny niedźwiedzia-kreta są podobne; walenie i ryby mają ten sam kształt ciała. Konwergencja określa ogólne podobieństwo między ptakami brodzącymi „kostkowymi” (żurawie i czaple). Zjawiska konwergencji obejmują „geograficzne” (klimatyczne) „paralele”, które wyrażają się w większej liczbie stałocieplnych zwierząt krajów zimnych w porównaniu z najbliższymi przedstawicielami regionów umiarkowanych, w większej liczbie lęgów u ptaków i odchodów u ssaków Arktyki, w zwiększonej pigmentacji ptaków tropikalnych.
Ta konwergencja w odległych grupach zwierząt jest spowodowana warunki zewnętrzne krewni muszą również brać pod uwagę pokrewieństwo. Pod tym względem obserwuje się zbieżność w obrębie klasy.
Równoległość.Jeśli rozbieżne wcześniej grupy wpadną w różne warunki egzystencji, mogą niezależnie, równolegle, wykształcić podobne cechy adaptacyjne. Na przykład u ryb głębinowych narządy luminescencyjne powstały równolegle i niezależnie; rozwój szablozębności u różnych kotów Czasami paralelizmy obejmują całe układy narządów lub część całej organizacji. Na przykład równoległy rozwój ryb oddychających płazów. Jest prawdopodobne, że podobieństwa między dwoma oddychającymi i płazami są określone przez pokrewieństwo tych grup. Szczególnie znaczące są paralelizmy w budowie serca i układ krążenia. Im więcej powiązań między organizmami, tym częściej równoległy rozwój obejmuje znaczne części całej organizacji. Świadczą o tym również dane paleontologiczne, na przykład rozwój trąby. Im bliżej siebie znajdują się organizmy, tym bardziej podobne są ich reakcje na tę samą zmianę w środowisku, tym bardziej podobne (równoległe) są zmiany w ich strukturze. Dla rozwoju równoległego ważne jest zarówno środowisko zewnętrzne, jak i czynniki wewnętrzne (tj. Historycznie ustalone dziedziczne struktury organizmu). Stosunek czynniki wewnętrzne z zewnętrznym determinuje ewolucję organizmów. Na przykład kopytne żyjące w różnych strefach klimatycznych uciekają przed drapieżnikami w locie. Dlatego u zwierząt kopytnych obserwuje się rozwój skrajnych stopni palców, a następnie stopniowe zmniejszanie się palców brzeżnych.
Genetyczne podstawy ewolucji równoległej. Przy pewnej zmianie czynników środowiskowych pokrewne organizmy zmieniają się podobnie. W walce o byt poprzez dobór naturalny zmiany te przybierają formę „dziedziczną”. Materiałem do selekcji są indywidualne odchylenia w budowie, czyli mutacje. Pokrewne lub bliskie formy z reguły dają te same mutacje (na przykład różne rodzaje Drosophila). Nie oznacza to jednak, że mutacje równoległe opierają się na tych samych zmianach w tych samych genach.
Wiadomo, że różne geny tego samego organizmu powodują podobne mutacje. Zwłaszcza jeśli te zmiany są niewielkie. A ponieważ w toku ewolucji wybierane są tylko kombinacje małych mutacji, zmiany równoległe mogą być budowane na innej podstawie genetycznej.
PRACA KURSU
w biologii
„Pochodzenie eukariota
Ukończone przez: Severova Vera
10 klasa, szkoła N55
Sprawdzone przez: Strukova N.A.
Ryazan 1999
1. Teoria symbiotyczna:
1.1 Podstawowe rozróżnienie między prokariontami a eukariontami;
1.2 Organizmy prokariotyczne;
1.3 Komórki eukariotyczne;
1.4 Mitochondria i plastydy;
1.5 Zdolność eukariontów do fotosyntezy;
2. Prekursorzy życia;
3. Organizmy fermentujące:
3.1 Źródła energii wczesnych komórek;
3.2 Metanogeneza jako źródło energii;
3.3 Cyjanobakterie i chlorooksybakterie;
4. Fotosynteza i powietrze;
5. Asocjacje i eukarionty:
5.1 Teorie kolejnych symbioz;
5.2 Symbiotyczne nabywanie mobilności;
5.3 Eukarionty - fotosynteza;
6. Spojrzenie na ewolucję komórki:
6.1 Hipoteza bezpośredniego synostwa;
6.2 Hipoteza: od sinic do alg;
6.3 Zgodność 2 hipotez: bezpośredniego synostwa i
endosymbioza;
7. Wnioski;
8. Literatura.
TEORIA ENDOSYMBIOZY
Wszystkie żywe organizmy na Ziemi są zwykle podzielone na prokariota i eukariota. Główna cecha prokariontów, w przeciwieństwie do eukariotów, jest ich brak kompletności Jądro komórkowe pokryty membraną. Komórki prokariotyczne są bardzo małe, rzędu 1 µm. Tom komórki eukariotyczne zawierające pełnoprawne jądro o objętości 800 - 1000 razy większej niż komórki prokariotyczne. Pod tym względem komórki - eukarionty potrzebują znacznie większego aparatu błonowego do wykonywania funkcji życiowych. Rzeczywiście, w komórce eukariotycznej zidentyfikowano struktury subkomórkowe otoczone błonami niezależnymi od plazmalemmy i od siebie nawzajem. Struktury te nazywane są organellami. Każda organella: mitochondria, chloroplast, jądro, lizosomy, aparat Golgiego itp. ma swoje specyficzne funkcje w życiu komórki.
Za pomocą podejść molekularno-biologicznych uzyskano przekonujące dowody na to, że wszystkie żywe istoty należy podzielić na 3 królestwa: bakterie, archeony i eukarionty. Bakterie i archeony powstały na Ziemi od wspólnego przodka – tzw. progenota – około 4 mld lat temu, a według paleontologów eukarionty pojawiły się na Ziemi 500 mln lat po prokariotach. Najbardziej popularną i wystarczająco uzasadnioną koncepcją pochodzenia komórek eukariotycznych jest obecnie idea wielokrotnej endosymbiozy różnych prokariotów. Według znanych mikrobiologów A. Kluyvera i K. Van Niela, to właśnie od starożytnych prokariotów miała miejsce selekcja komórek o najbardziej ekonomicznych i często wielofunkcyjnych reakcjach biochemicznych, które stały się podstawą najważniejsze sposoby metabolizm.
Symbiotyczna teoria pochodzenia i ewolucji komórek opiera się na dwóch nowych dla biologii koncepcjach. Zgodnie z pierwszą z nich najbardziej fundamentalnym rozróżnieniem w żywej przyrodzie jest rozróżnienie między prokariotami i eukariontami, tj. między bakteriami a organizmami składającymi się z komórek z prawdziwym jądrem – protistami, zwierzętami, grzybami i roślinami. Druga koncepcja mówi, że niektóre części komórek eukariotycznych powstały w wyniku ewolucji symbioz, tj. tworzenie trwałych związków między organizmami różne rodzaje. Przyjmuje się, że trzy klasy organelli – mitochondria, rzęski i fotosyntetyzujące plastydy – wywodzą się od wolno żyjących bakterii, które w wyniku symbiozy zostały włączone w określonej kolejności do komórek gospodarzy prokariotycznych. Teoria ta w dużej mierze opiera się na neodarwinowskich ideach opracowanych przez genetyków, ekologów, cytologów i innych naukowców, którzy powiązali genetykę Mendla z ideą darwinowską. naturalna selekcja Czerpie również z zupełnie nowych lub niedawno odrodzonych dziedzin nauki: biologii molekularnej, zwłaszcza budowy białek i sekwencji aminokwasów, mikropaleontologii, badającej najstarsze ślady życia na Ziemi, a nawet fizyki i chemii atmosfera, ponieważ nauki te są związane z gazami pochodzenia biologicznego. Wszystkie organizmy komórkowe można podzielić na pięć królestw: królestwo prokariotyczne (Monera, do którego należą bakterie) i cztery królestwa eukariotyczne (Protoctista, Animalia, Fungi, Plantae). Protoktyści to organizmy eukariotyczne, które nie są zwierzętami, grzybami ani roślinami. Królestwo protoktystów obejmuje algi, pierwotniaki, śluzowce i inne organizmy eukariotyczne niejasnego pochodzenia. Protisty definiuje się bardziej restrykcyjnie jako jednokomórkowe eukarionty. Tak więc królestwo Protoctista obejmuje nie tylko protisty - jednokomórkowe eukarionty, ale także ich najbliższych wielokomórkowych potomków, takich jak czerwony i algi brunatne i także wiele mikroorganizmów podobnych do grzybów, takich jak chytrids. Zgodnie z tradycyjnym poglądem na bezpośrednie filiowanie, organelle komórkowe, takie jak mitochondria i plastydy, powstały w wyniku kompartmentalizacji samej komórki.
KILKA FORM PROKARIOTÓW
JEDEN EUKARIOT
Komórki są układami związanymi z błonami, najmniejszymi jednostkami biologicznymi zdolnymi do samoreprodukcji. Nawet najmniejsze komórki zawierają geny w postaci cząsteczek DNA (jednej lub więcej) oraz aparatu do syntezy białek, składającego się z kilku rodzajów RNA i wielu białek. Wszystkie komórki zawierają rybosomy, ciała o średnicy około 0,02 µm, zbudowane z co najmniej trzech rodzajów RNA i około pięćdziesięciu różnych białek. Organizmy prokariotyczne to najmniejsze jednostki biologiczne, które spełniają tę definicję komórki. Nie mają jąder. Z drugiej strony pojedyncze komórki eukariotyczne (posiadające jądro) w pewnym sensie wcale nie są jednostkami. Są to kompleksy różnych jednostek syntetyzujących białka (nukleocytoplazma, mitochondria i plastydy), których typ i liczba różni się w zależności od gatunku. Tak więc, zgodnie z tą teorią, komórki eukariotyczne powstały w wyniku współpracy początkowo niezależnych elementów, połączonych w pewien porządek. Taylor nazwał tę ideę odrębnego pochodzenia i rozwoju części komórki eukariotycznej oraz ich późniejszego zjednoczenia teorią kolejnych endosymbioz. Warunki wysoka temperatura a kwasowość doprowadziła do wytworzenia w nukleocytoplazmie specjalnej klasy białek - histonów - w celu ochrony jej DNA. Mitochondria były w stanie rozłożyć związki trójwęglowe na dwutlenek węgla i wodę. Symbiotyczny związek nukleocytoplazmy i mitochondriów zaczął się zmieniać w kierunku wzmocnienia ich wzajemnej zależności. W skład tego kompleksu wchodziły organelle ruchowe - rzęski i wici. Po tej transformacji nastąpiła transformacja morfogenetyczna, która polegała na rozwoju wyraźnych asymetrii komórkowych. Wraz z integracją współzależnej nukleocytoplazmy, mitochondriów, undulipodiów zakończono tworzenie struktury poligenomicznej komórki eukariotycznej. Pierwsza komórka eukariotyczna była heterotroficzna. Jadała gotowe związki organiczne. U tych eukariontów powstały procesy mitozy, a następnie mejozy. I wreszcie nabycie przez eukarionty zdolności do fotosyntezy nastąpiło w procesie stawania się główną organizacją eukariotyczną, a po jego zakończeniu – ostatnim wydarzeniem w tej epopei. Symbiozy stawały się coraz bardziej zintegrowane. Obecnie zależność każdej organelli od produktów metabolicznych innych organelli stała się tak kompletna, że tylko nowoczesne metody analiza pozwala prześledzić szlaki metaboliczne pierwotnych partnerów. Eukarionty różnią się budową, ale metabolicznie pozostają jednolite. Opracowano metaboliczne sztuczki eukariontów różne bakterie jeszcze przed czasem, kiedy się zjednoczyły, zamieniając się w żywiciela i jego organelle. Prokarionty osiągnęły wielką różnorodność w sposobie pozyskiwania energii i zawiłościach biosyntezy. Pozostawiły ślad swojego istnienia na długo przed tym, zanim z komórki eukariotycznej powstały tak duże organizmy jak zwierzęta ze szkieletem.
Schemat pro I konwergencja komórek eukariotycznych przez symbiozę
PREKURSORZY ŻYCIA
Przodkowie dzisiejszych organizmów powstali ponad trzy miliardy lat temu ze związków organicznych powstałych na powierzchni Ziemi. Atrakcyjność tej koncepcji wynika częściowo z jej testowalności.
S. Miller i H. Urey wykazali, że w warunkach atmosferycznych, jakie miały miejsce na młodej Ziemi, mogą powstawać aminokwasy i inne cząsteczki organiczne. Różne rodzaje cząsteczki organiczne powstały spontanicznie podczas pierwszego miliarda lat istnienia Ziemi. W przestrzeni międzygwiezdnej iw meteorytach znaleziono wiele cząsteczek organicznych. Oznacza, związki organiczne powstają podczas interakcji lekkich elementów, w obecności źródeł energii. Wszechstronność mechanizm chemiczny bioreprodukcja oznacza, że całe życie na Ziemi pochodzi od wspólnych przodków: całe obecne życie wywodzi się z komórek z systemami informacyjnymi opartymi na replikacji DNA i syntezie białek kierowanej przez informacyjny RNA.
FERMENTUJĄCE ORGANIZMY
Możliwe, że kwasy nukleinowe od samego początku były oddzielane od środowiska przez membrany lipoproteinowe. Informacje zaczęły być kodowane przez sekwencję nukleotydów w cząsteczkach DNA. Najstarsze komórki wykorzystywały ATP i aminokwasy, które są częścią białek, jako bezpośrednie źródła energii i jako Elementy konstrukcyjne. Tak więc - najstarsze życie istniało w postaci bakterii heterotroficznych, które otrzymywały żywność i energię z materiału organicznego pochodzenia abiotycznego. Neodarwinowska ewolucja organiczna wymaga trzech zjawisk: reprodukcji, mutacji i selektywnej presji środowiskowej. Rozmnażanie jest niemożliwym do zastosowania warunkiem ewolucji biologicznej. Przy wystarczająco dokładnej reprodukcji mutacje i dobór naturalny są nieuniknione. Wyjaśnienie pochodzenia takiej reprodukcji oznacza zrozumienie pochodzenia samego życia. Mutacja, główne źródło zmienności w ewolucji, prowadzi do dziedzicznych zmian w samoreprodukującym się systemie. Po opracowaniu niezawodnej reprodukcji rozpoczęła się ewolucja neodarwinowska. Organizmy, które wyewoluowały szlaki syntetyczne potrzebna klatka komponentów, miał przewagę nad innymi. Najstarsze autotrofy, czyli organizmy zdolne do zaspokojenia swoich potrzeb energetycznych i materia organiczna ah, ze względu na źródła nieorganiczne, istniały bakterie beztlenowe wytwarzające metan. Zdolność do fotosyntezy została po raz pierwszy rozwinięta u bakterii wrażliwych na tlen. Absorbujący światło słoneczne za pomocą bakteryjnego chlorofilu przekształcili atmosferyczny dwutlenek węgla w substancje organiczne.Pojawienie się fotosyntezy beztlenowej nieodwracalnie zmieniło powierzchnię i atmosferę Ziemi.
Dwutlenek węgla został usunięty z atmosfery i przekształcony w materię organiczną zgodnie z ogólnym równaniem fotosyntezy: 2H2X+CO2=CH2O+2X+H2O. Bakterie fotosyntetyczne, które wykorzystywały siarkowodór do redukcji CO2 osadzonej siarki elementarnej. Dały początek bakteriom zdolnym do fotosyntezy z uwalnianiem tlenu. Ten rodzaj fotosyntezy powstał prawdopodobnie w wyniku nagromadzenia mutacji w fotosyntetycznych bakteriach siarkowych. Droga ta doprowadziła do pojawienia się sinic lub sinic. Niedawno odkryte i zbadane nowa grupa fotosyntetyzujące prokarionty produkujące O2 - chloroksybakterie. Można przypuszczać, że przodkami plastydów występujących w komórkach alg i roślin były cyjano- i chlorooksybakterie. Gdy w wyniku mikrobiologicznej fotosyntezy gromadził się O2, atmosfera ziemska zaczęła się utleniać, co spowodowało kryzys wszelkiego życia – do tego czasu świat zamieszkiwały bakterie beztlenowe. Aby przetrwać, drobnoustroje musiały unikać tlenu, ukrywając się na bagnach, lub musiały rozwinąć metabolizm zdolny do ochrony przed rosnącą ilością tlenu.
FOTOSYNTEZA I POWIETRZE
Około 2 miliardów lat temu, jeden po drugim, miały miejsce różne procesy ewolucyjne, które stworzyły różnorodność metaboliczną. Zdolność do wytwarzania określonych enzymów może być przenoszona w obrębie tego samego gatunku za pomocą cząsteczek wirusopodobnych zwanych plazmidami. Zdolności biosyntetyczne prokariontów są uderzające: z dwutlenku węgla, kilku soli i zredukowanych związków nieorganicznych, które służą jako źródła energii, mikroorganizmy chemoautotroficzne mogą syntetyzować w nieoświetlonej wodzie zawierającej tlen wszystkie najbardziej złożone makrocząsteczki niezbędne do wzrostu i rozmnażania. Te procesy biochemiczne doprowadziły do powstania układów krążenia pierwiastków nawet wtedy, gdy nie było roślin, grzybów, zwierząt.
asocjacje i eukarionty.
Spojrzenie na ewolucję komórki.
Oddział bezpośredni.
Tradycyjny pogląd na pochodzenie i ewolucję komórek eukariotycznych jest taki, że wszystkie żywe organizmy pochodzą bezpośrednio od jednej populacji przodków w wyniku nagromadzenia mutacji pod wpływem doboru naturalnego.
Mutacje punktowe, duplikacje, delecje i inne zmiany dziedziczne, o których wiadomo, że odegrały rolę w ewolucji zwierząt i roślin, są również odpowiedzialne za różnicowanie komórek eukariotycznych od prokariotycznych - teoria bezpośredniego filiowania. tj. pojęcie niesymbiotycznego pochodzenia komórek. Być może zielone algi są potomkami przodków wszystkich roślin. Wiele cech tych glonów sugeruje, że one i rośliny mają wspólnego przodka. Filogeneza zwierząt jest bardziej złożona, ale koncepcja bezpośredniego pokrewieństwa prawdopodobnie odnosi się również do ich ścieżek ewolucyjnych. Gąbki najwyraźniej wyewoluowały z protistów, niezależnie od innych grup zwierząt. Tak więc, przynajmniej w przypadku zwierząt i roślin, hipoteza, że złożone organizmy eukariotyczne wyewoluowały z prostszych eukariotów poprzez akumulację indywidualnych mutacji pod wpływem doboru naturalnego, jest całkiem uzasadniona. Innowacje ewolucyjne polegają na zachowaniu wysoce doskonałych korzystnych genów. Tak więc nowe organizmy o zwiększonej zdolności do kontrolowania swojego środowiska ewoluowały bardziej dzięki utrzymywaniu duplikacji i rekombinacji wysoce przystosowanych genów niż nabywaniu nowych właściwości poprzez proste, losowe mutacje punktowe. Hipoteza bezpośredniego synostwa nie wyjaśnia pochodzenia eukariontów od prokariotów. Na przykład przerwa między sinicami niezdolnymi do mitozy a takimi formami z w pełni rozwiniętą mitozą, jak krasnorosty, jest trudna do zrozumienia z punktu widzenia bezpośredniego filiowania, ale naturalnie wynika z teorii symbiozy.
Zgodnie z teorią bezpośredniego potomstwa i teorią symbiozy wszystkie organizmy na Ziemi wywodzą się od biochemicznie podobnych przodków bakterii, a drobnoustroje eukariotyczne dały początek roślinom i zwierzętom. Bakterie fotosyntetyzujące dały początek glonom, a ostatecznie roślinom, a niektóre glony utraciły plastydy i stały się przodkami grzybów i zwierząt. Teoretycy bezpośredniego filiowania zgadzają się, że plastydy i inne organelle eukariotyczne, w tym jądro, powstają w wyniku różnicowania w samych komórkach i nazywają to mitem botanicznym.
Filogeneza typów roślin
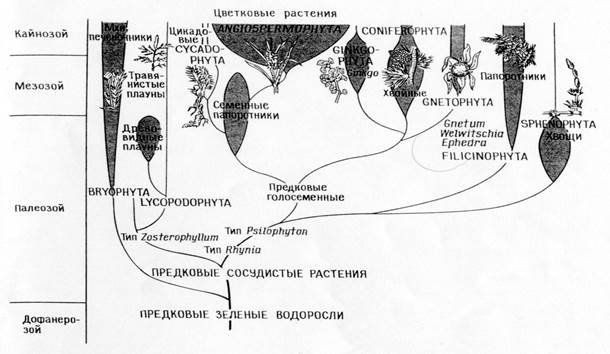
Porównanie filogenez odpowiadających teorii bezpośredniego synostwa (A)
i teoria symbiozy (B)
Różnice między teorią bezpośredniego synostwa a teorią symbiozy:
Teoria bezpośredniego synostwa.
Główna dychotomia: zwierzęta - rośliny
Fotosyntetyczne eukarionty (algi i rośliny zielone) wyewoluowały z fotosyntetycznych prokariontów (niebiesko-zielone algi)
Rośliny i ich systemy fotosyntetyczne ewoluowały monofiletycznie w prekambrze.
Zwierzęta i grzyby wyewoluowały z autotrofów, które utraciły plastydy.
Mitochondria różnicowały się wewnątrzkomórkowo w algach fotosyntetyzujących.
W chwastach prawoskrętnych unlipodia, aparat mitotyczny i wszystkie inne organelle eukariotyczne różnicowały się wewnątrzkomórkowo.
Wszystkie organizmy wywodzą się od swoich przodków – prokariontów poprzez nagromadzenie pojedynczych mutacji.
KONSEKWENCJE: Zgodnie z teorią muszą istnieć prawe chwasty; trudno jest powiązać tę teorię z zapisem kopalnym i skonstruować odpowiednią filogenezę.
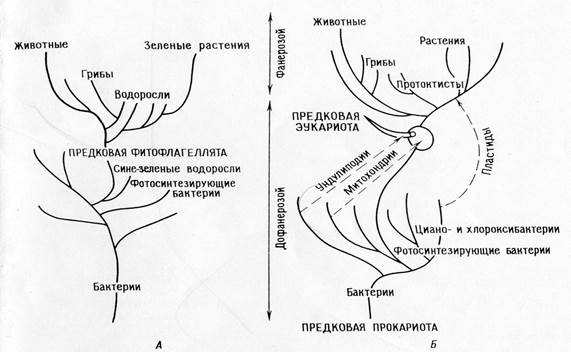
Teoria kolejnych endosymbioz:
Główna dychotomia: prokarioty - eukarionty.
Fotosyntetyzujące eukarionty (algi jądrowe i rośliny) oraz niefotosyntetyzujące eukarionty (zwierzęta, grzyby, pierwotniaki) wyewoluowały ze wspólnych heterotroficznych przodków (ameboflagellaty).
Ewolucja fotosyntezy u Archaean była monofiletyczna u bakterii (w tym cyjano- i chlorooksybakterii), protisty fotosyntetyzujące powstały nagle, gdy ich heterotroficzni protisty-przodkowie weszli w symbiozę z fotosyntetycznymi prokariotami (protoplastidami).
Zwierzęta oraz większość glonów i grzybów pochodzi bezpośrednio od heterotroficznych protistów.
Mitochondria były już obecne u protistów heterotroficznych, kiedy te ostatnie nabyły plastydy w wyniku symbiozy. Promitochondria były wcześniej nabywane w sposób symbiotyczny.
Mitoza rozwinęła się u heterotroficznych protistów eukariotycznych poprzez różnicowanie układu unlipodialnego.
Prokarioty ewoluowały od wczesnych prokariotycznych przodków poprzez serię pojedynczych zdarzeń mutacyjnych, eukarioty również ewoluowały od eukariotycznych przodków poprzez pojedyncze zdarzenia mutacyjne, a eukariotyczni przodkowie ewoluowali od prokariotów poprzez serię symbioz.
Konsekwencje: Teoria wyjaśnia szlaki biochemiczne i ich redundancję w organellach. Pozwala to na zbudowanie spójnej filogenezy, która wyjaśnia „lukę biologiczną” między proterozoikiem a fanerozoikiem. Teoria przewiduje, że organelle muszą zachowywać pozostałości systemów replikacyjnych.
mit botaniczny
Od czasów Jussieu niższe rośliny zwane talofitami, a wyższe - mszakami i tracheofitami. Według mitu botanicznego, wspólni przodkowie bakterii fotosyntetyzujących i sinic, poprzez bezpośrednie filiowanie, dali początek glonom eukariotycznym, w tym algom morskim. Do dziedziny botaniki trafiły grupy organizmów, na których filogenezę najbardziej oddziałuje koncepcja bezpośredniej filiacji – są to bakterie, algi, grzyby i rośliny.
Hipotetyczne organizmy, które mogłyby być łącznikiem między sinicami a zielonymi algami, nazwano fitomonadami przodków (fitomonadami przodków) lub prawicowcami. Niektórzy autorzy argumentowali, że eukarionty Cyanidium i Cyanophora, które są podobne w swoich układach barwnikowych do sinic, mogą być odpowiednimi kandydatami do roli prawicowców. U talofitów nie stwierdzono form pośrednich między niemitotycznym stanem cyjanobakterii a w pełni rozwiniętą typową mitozą zielonych alg i roślin. Chociaż grzyby i rośliny kwitnące całkowicie pozbawione undulipodiów, charakteryzują się mitozą i mejozą, które wymagają wrzecion składających się z mikrotubul. Istnieje powszechna zgoda co do tego, że wszystkie organizmy wielokomórkowe z mikrotubulami pochodzą od mikroorganizmów eukariotycznych z undolipodiami, ale nikt nie wie, z jakiej grupy protistów. Wzmocnieniem mitu botanicznego była starożytność i złożoność fotosyntezy, a także fakt, że wiele organizmów (bakterie, euglenoidy, rośliny pasożytnicze), zarówno w naturze, jak iw laboratorium, miało utratę fotosyntezy w wyniku mutacji. Ponieważ fotosynteza jest procesem beztlenowym, który ostatecznie zapewnia istnienie wszystkich organizmów, musiała rozwinąć się bardzo wcześnie w historii życia na naszej planecie, jeszcze zanim pojawiła się mitoza, która wymaga tlenu. w fotosyntezie bierze udział wiele pigmentów, lipidów i enzymów, uporządkowanych w bardzo złożonych błonach. Jednocześnie szczegóły mechanizmów fotosyntezy u alg i roślin z jednej strony oraz u cyjanobakterii z drugiej są uderzająco podobne. Wszystkie te organizmy wydzielają gazowy tlen z wody, który służy jako źródło atomów wodoru wykorzystywanych do redukcji CO2 do materii organicznej komórki. Wiele glonów i pierwotniaków nie ma ani mitozy, ani systemu genetycznego podobnego do zwierząt i roślin. Orzęski mają specjalny system genetyczny z 2 typami jąder, a komórki są często wielojądrowe. Jeden rodzaj jąder to makrojądra, a drugi to znacznie mniejsze mikrojądra. Mikrojądra są fizjologicznie nieistotne. przechowują jednak kopie genów, a podczas procesu płciowego przechodzą mejozę. Fizjologicznie niezbędne makrojądra służą jako miejsce transkrypcji RNA, a tym samym rządzą fizjologią komórki. Jedną z trudności w wyjaśnieniu związku między sinicami a algami eukariotycznymi jest brak danych na temat pochodzenia undolipodiów: nie znaleziono pośrednich powiązań między sinicami, które nigdy nie mają undolipodiów, a wieloma algami, które je mają. Innym problemem jest kompletna nieobecność proces płciowy u cyjanobakterii: między sinicami bezpłciowymi a algami, z których większość charakteryzuje się procesem płciowym, nie było form pośrednich. Stwierdzono, że większość glonów ma na pewnym etapie undolipodia koło życia. Ruchome struktury glonów, pierwotniaków, zwierząt i roślin okazały się takie same: wszystkie składają się z mikrotubul o średnicy 0,024 mikrona. Okazało się, że wrzeciono mitotyczne, nawet u grzybów nieposiadających undolipdiów, składa się z białkowych mikrotubul o takiej samej średnicy jak u undolipdiów. Przez pół wieku wydawało się niezwykle prawdopodobne, że glony eukariotyczne wyewoluowały z sinic. Ten botaniczny mit doprowadził do przypuszczeń, które przez wiele lat służyły zarówno jako bodziec do badań, jak i układ współrzędnych, w której dokonano oceny uzyskanych wyników. Twierdzenie, że powiązania między sinicami a eukariotycznymi algami wymarły, nie pozostawiając żadnych śladów skamielin i żywych reliktów, stawało się coraz mniej przekonujące w miarę pojawiania się nowych danych na temat żywych drobnoustrojów i skamieniałości. Główne ogniwa pośrednie między fotosyntetyzującymi prokariotami a eukariontami nie wymarły bez śladu, jak tego wymaga mit botaniczny – nigdy nie istniały. Zgodnie z teorią kolejnych endosymbioz, sinice są wprawdzie przodkami, ale tylko plastydami, a nie resztą komórki eukariotycznej, a już na pewno nie jej jądrem. A jednak w kwestii pochodzenia jądra hipoteza bezpośredniego synostwa i teoria symbiozy są zgodne. Możliwe do pomyślenia modyfikacje skrajnej wersji teorii symbiozy obejmują na przykład ideę symbiotycznego pochodzenia plastydów, ale nie mitochondriów, lub symbiotycznego pochodzenia plastydów i mitochondriów, ale nie unlipodiów. Zwolennicy bezpośredniego synostwa wysunęli również założenie o pochodzeniu jądra poprzez symbiozę.
Krytyczne dowody na endosymbiotyczne pochodzenie chloroplastów i mitochondriów.
Najbardziej znaczący dowód pochodzenia najważniejszych organelli komórek eukariotycznych – chloroplastów i mitochondriów z prokariotów uzyskano w ostatnie lata podczas badania struktury rybosomalnego RNA. Biolog molekularny
Już w latach 80. K. Woz wykazał, że budowa RNA z rybosomów chloroplastów roślinnych jest niemal identyczna z budową RNA niektórych sinic. Jednocześnie grupa naukowców odkryła, że RNA rybosomów pochodzących z mitochondriów różnych eukariontów jest podobne do RNA niektórych bakterii, w szczególności bakterii z rodzaju Paracoccus. Zestaw enzymów oddechowych tej bakterii jest bardzo podobny do
„zespół oddechowy” zwierzęcych mitochondriów.
Dane te wyraźnie wspierają bardziej prawdopodobne endosymbiotyczne pochodzenie komórek eukariotycznych. Zjawisko endosymbiozy jest nadal niezwykle rozpowszechnione w przyrodzie. Wiadomo, że na ciele niektórych ryba morska często osiedlają się świetliste bakterie, które nie tylko odnoszą ogromne korzyści dla własnego życia, ale są również bardzo przydatne dla organizmów żywiciela. Wszystko to w przekonujący sposób świadczy o endosymbiotycznym pochodzeniu komórek eukariotycznych ze znacznie mniejszych komórek prokariotycznych. Jednak głównym pytaniem jest ustalenie charakteru komórki gospodarza, ponieważ komórki eukariotyczne mają dobrze uformowane jądro z otoczką, podczas gdy prokarioty takiego jądra nie mają. L. Margelis uważa na przykład, że rolę komórek gospodarza mogły pełnić prokarioty – poprzednicy współczesne bakterie spokrewniony z mykoplazmami - bardzo prymitywnie zbudowanymi prokariontami, praktycznie pozbawionymi ścian komórkowych. W Ostatnio pojawiły się prace japońskiego naukowca i są szeroko dyskutowane
T. Oshima. Opierając się na dokładnym badaniu struktury rybosomalnego RNA wielu pro- i eukariontów, w tym wielu przedstawicieli mało zbadanego dotąd superkrólestwa archeonów, autor ten dochodzi do wniosku, że komórka gospodarza była jednym z przodków współczesnych archeonów. Trzeba powiedzieć, że przedstawiciele tego superkrólestwa, w przeciwieństwie do bakterii, mają wiele cech biochemicznych i biologii molekularnej wspólnych z eukariotami. W najnowszych pracach T. Oshimy podaje się argumenty przemawiające za tym, że komórka gospodarza w momencie pojawienia się eukariontów była przodkiem archeonów, podobnie jak jeden ze współczesnych przedstawicieli tych prokariotów - termoplazma. Jakie będzie ostateczne rozwiązanie problemu ustalenia natury komórki gospodarza, która dała początek komórce eukariotycznej w procesie wielokrotnej endosymbiozy, pokażą dalsze badania.
Przedstawiono obecnie najbardziej popularną i potwierdzoną hipotezę pochodzenia komórek eukariotycznych
endosymbioza różnych prokariontów należących do bakterii i archeonów, która wielokrotnie występowała w procesie ewolucji w ciągu pierwszych 1,5–2 miliardów lat istnienia żywych komórek na Ziemi. Decydujące dowody na endosymbiotyczne pochodzenie chloroplastów i mitochondriów uzyskali czołowi światowi biolodzy badający strukturę rybosomalnego RNA. Dane te wyraźnie wspierają wysokie prawdopodobieństwo endosymbiotyczne pochodzenie komórek eukariotycznych od znacznie mniejszych (o kilka rzędów wielkości) komórek prokariotycznych.
LITERATURA
1. Czasopismo edukacyjne Sorosa. Biologia, Chemia, Nauki o Ziemi, Fizyka, Matematyka. N 5 1998
2 Margelis L. Rola symbiozy w ewolucji komórek. Za. z angielskiego, M. MIR 1983
3 Podstawy mikrobiologii. M. "Medycyna" 1992.
4 Chakhina L.N. Koncepcja A.S. Famincyn o znaczeniu symbiozy w ewolucji L.; Nauka, 1981, s. 165 - 181 .
5 Alberts B. Biologia molekularna komórki; Za. z angielskiego. M. Mir 1986 t.1
Termin Prokaryotae (przedjądrowy) odnosi się do mikroorganizmów o prymitywnej organizacji struktur jądrowych. Nazwę tę zaproponował kanadyjski naukowiec Murray w 1968 roku.
W drugiej połowie XX wieku. stało się jasne, że wszystkie żywe organizmy z struktura komórkowa, można przypisać do jednej z dwóch grup – prokariotów lub eukariontów. Komórki prokariotyczne są prostsze - pozbawione są jądra otoczonego otoczką. Przez pewien czas uważano, że terminy „prokarionty” i „bakterie” są synonimami i że bakterie reprezentują niezależną i zunifikowaną gałąź ewolucji organizmów żywych. Jednak stopniowo gromadzono dowody na to, że różnice między niektórymi bakteriami mogą być znaczące i wskazują, że nie mają one bliskiego związku ewolucyjnego. Zwłaszcza bardzo ważne dołączone do danych dotyczących struktury rybosomów i transportu kwasów rybonukleinowych (RNA). Obecnie na podstawie wyników porównania sekwencji zasad w cząsteczkach rybosomalnego RNA ocenia się pokrewieństwo organizmów. Tego rodzaju dane, jak również inne cechy prokariontów, pokazały, że w rzeczywistości prokarioty należy podzielić na co najmniej dwie grupy o różnym pochodzeniu i ścieżce ewolucyjnej. Ponadto komórki eukariotyczne są nie tylko bardziej złożone, ale także posiadają elementy, których nie mogły uzyskać od przodków współczesnych prokariotów. Są to przede wszystkim rybosomy cytoplazmatyczne, które różnią się od rybosomów prokariotycznych. Podstawowe podobieństwo kod genetyczny, organizacja makrocząsteczek i biochemiczny aparat syntezy białek wskazuje na jedność pochodzenia wszystkich żywych organizmów. Przyjmuje się, że istniał pewien wspólny przodek - „progenot”, ale nie wiadomo, kim był. To potomstwo mogło dać początek trzem niezależnym gałęziom drzewa ewolucyjnego. Z punktu widzenia systematyki hierarchicznej gałęzie te proponuje się traktować jako domeny o randze wyższej niż tradycyjne królestwa. Te domeny (imperia) to Eubacteria, Archaebacteria, Eukaryota (eubakteria, archebakterie i eukarionty). Ogólne i różnicujące cechy archebakterii, eubakterii i eukariontów podano w tabeli.
Główną cechą komórki prokariotycznej jest brak jądra, oddzielone od cytoplazmy podwójną błoną. Materiał dziedziczny jest skoncentrowany w chromosomie bakteryjnym, zwykle reprezentowanym jako kolista dwuniciowa cząsteczka DNA. Niektóre krętki mają chromosomy liniowe, podczas gdy promieniowce mają chromosomy „pseudo-pierścieniowe”. Nitkowata cząsteczka DNA działa jak jądro i znajduje się w centralnej strefie komórki, zwanej „nukleoidem”. W mikroskopie elektronowym na skrawku komórki bakteryjnej ta strefa wygląda na jaśniejszą niż reszta cytoplazmy.
Komórka prokariotyczna posiada szereg podstawowych cech związanych zarówno z jej ultrastrukturalnością jak i organizacja chemiczna. Przede wszystkim w komórce bakteryjnej nie ma sieci membranowej - retikulum endoplazmatycznego. W komórkach prokariotycznych błona cytoplazmatyczna leżąca pod ścianą komórkową i otaczająca protoplast od zewnątrz jest jedyną błoną w komórce.
Błona cytoplazmatyczna otaczająca każdą komórkę określa jej wielkość i zapewnia utrzymanie znaczących różnic między zawartością komórki a środowiskiem. Ale membrana to nie tylko mechaniczna przegroda. Służy również jako wysoce selektywny filtr, który utrzymuje różnicę stężeń jonów po obu stronach membrany i umożliwia przedostawanie się składników odżywczych do komórki i usuwanie produktów przemiany materii. Błona cytoplazmatyczna jest zespołem cząsteczek lipidów i białek utrzymywanych razem przez oddziaływania niekowalencyjne. W mikroskop elektronowy membrany wyglądają jak arkusze o grubości około 7 nm z wyraźną trójwarstwową strukturą. Podwójna warstwa cząsteczek lipidów określa cechy strukturalne błony, podczas gdy białka odpowiadają za większość funkcji błony.
Główne cechy archebakterii, eubakterii i eukariontów
| oznaki | archebakterie | eubakterie | eukarionty |
| Typowe organizmy | metanogeny, ekstremalne termofile, halofile | główne grupy bakterii | protisty, grzyby, rośliny, zwierzęta |
| Typowe wymiary, mikrony | 0,5-4 | 0,5-4 | powyżej 5 |
| Genom | chromosom pierścieniowy | chromosom pierścieniowy | jądro z wieloma złożonymi chromosomami |
| Struktury błony: retikulum endoplazmatyczne lizosomy kompleksu Golgiego | nie nie nie | nie nie nie | tak tak tak |
| mitochondria | NIE | NIE | Jest |
| Chloroplasty | brak (czasami tylakoidy) | NIE | nie (u grzybów i zwierząt), tak (u roślin i alg) |
| Ściany komórkowe | białko, pseudomureina | mureina, lipopolisacharyd | różny |
| Rybosomy | 70S* | 70S | 80S (w cytoplazmie) + 705 rybosomów organelli, jak u bakterii |
| polimeraza RNA | złożony | prosty | złożony |
| wici | prosty | proste, złożone z jednego lub więcej włókienek | złożona struktura (2х9)+2 |
| Wiązanie azotu | Jest | Jest | NIE |
| Oddychanie beztlenowe | Jest | Jest | NIE |
| Chemosynteza | Jest | Jest | NIE |
S* - jednostka sedymentacji Svedberga charakteryzująca wielkość rybosomu
W błonie cytoplazmatycznej prokariotów występuje wiele inwazji - inwazji pełniących różne funkcje. Mogą zawierać enzymy i wyglądać jak pęcherzyki lub być mezosomami i biorą udział w podziale komórek. Błona cytoplazmatyczna prokariotów jest miejscem lokalizacji enzymów metabolizmu energetycznego, a zatem pełni funkcję mitochondriów, których nie ma w komórkach prokariotycznych. Dlatego bakterie z wysoki poziom metabolizm oddechowy, błona cytoplazmatyczna jest szczególnie rozwinięta, tj. ma wiele wgnieceń, co zwiększa jego powierzchnię. Błona cytoplazmatyczna bierze również udział w podziale nukleoidu. Okrągły DNA jest utrwalony na błonie i po replikacji (podwojeniu) rozdziela się na dwie nowe komórki w wyniku wzrostu błony. Pokrywy przetrwalników powstają z błony cytoplazmatycznej. Stanowi również główną barierę dla przenikania substancji do wnętrza komórki. Błona cytoplazmatyczna nie przepuszcza dużych cząsteczek, nie jest zdolna do pinocytozy i egzocytozy.
Struktury znajdujące się poza błoną cytoplazmatyczną - ściana komórkowa, torebka, błona śluzowa, wici, fimbrie, pilusy, kosmki - są strukturami powierzchniowymi.
Ściana komórkowa jest ważnym i obowiązkowym elementem strukturalnym większości komórek prokariotycznych (prawdziwych bakterii, nie licząc archebakterii i mykoplazm), zlokalizowanym pod torebką lub błoną śluzową lub w przypadku jej braku w kontakcie z otoczeniem. Ściana komórkowa służy jako mechaniczna bariera między protoplastem a otoczenie zewnętrzne i nadaje komórce pewien właściwy jej kształt.
Pod względem struktury i składu chemicznego ściana komórkowa prokariotów znacznie różni się od ściany organizmów eukariotycznych. Ściana komórkowa prokariotów składa się z kilku warstw, z których główna – szkieletowa – warstwa odpowiedzialna za wytrzymałość, jest utworzona ze specjalnej substancji – peptydoglikanu mureiny, który jest syntetyzowany tylko przez komórkę prokariotyczną i nie występuje w innych organizmach. W grupie prawdziwych prokariotów wyróżnia się dwa główne typy budowy ściany komórkowej. Te dwa typy odpowiadają bakteriom Gram-dodatnim i Gram-ujemnym. Nazwy tych grup pochodzą od zdolności lub niemożności różne bakterie plama wg Grama. W 1884 roku duński lekarz K. Gram, który zajmował się barwieniem tkanek, zaproponował metodę barwienia, którą później zastosowano w przypadku bakterii. Jeśli utrwalone komórki eubakterii traktuje się najpierw roztworem fioletu krystalicznego, a następnie jodem, powstaje barwny kompleks. Podczas późniejszego traktowania alkoholem, w zależności od budowy ściany komórkowej, losy kompleksu są różne: u tzw. gatunków, kolorowy kompleks jest wypłukiwany z komórki i ulegają one odbarwieniu. Stwierdzono, że barwny kompleks tworzy się na protoplastach, ale jego retencja przez komórkę lub wypłukiwanie z niej podczas późniejszego traktowania alkoholem determinowane jest cechami strukturalnymi ściany komórkowej. Znaleziono metodę barwienia metodą Grama szerokie zastosowanie w praktyce mikrobiologicznej do różnicowania bakterii.
Ściany komórkowe eubakterii Gram-dodatnich i Gram-ujemnych różnią się znacznie zarówno pod względem składu chemicznego, jak i ultrastruktury.
Większość ściany komórkowej bakterii Gram-dodatnich stanowi specyficzny heteropolimer - mureina peptydoglikanu. Szkielet polisacharydowy cząsteczki zbudowany jest z naprzemiennych reszt N-acetyloglukozaminy i kwasu N-acetylomuraminowego, połączonych ze sobą wiązaniami glikozydowymi. Oprócz peptydoglikanu ściany komórkowe eubakterii Gram-dodatnich zawierają kwasy teichojowe i polisacharydy. Kwasy teichojowe to polimery na bazie alkohole wielowodorotlenowe(rybitol i glicerol), połączone wiązaniami fosfodiestrowymi.
Eubakterie Gram-ujemne mają znacznie bardziej złożoną strukturę ściany komórkowej niż bakterie Gram-dodatnie. Zawiera dużo więcej makrocząsteczki różnych typ chemiczny. Peptydoglikan tworzy tylko wewnętrzną warstwę ściany komórkowej, luźno przylegając do błony cytoplazmatycznej. Na zewnątrz peptydoglikanu znajduje się dodatkowa warstwa ściany komórkowej - zewnętrzna męmbrana, który składa się z fosfolipidów (typowych dla błon elementarnych), białek, lipoprotein i lipopolisacharydów.
Stwierdzono empirycznie, że różnice między dwiema grupami bakterii w odniesieniu do barwienia metodą Grama są skorelowane z wieloma innymi ważnymi cechami. Na przykład bakterie Gram-dodatnie rozmnażają się przez rozszczepienie binarne z powodu tworzenia poprzecznej przegrody w komórce, Gram-ujemne - przez tworzenie zwężenia. Te pierwsze są wrażliwe na penicylinę, te drugie nie. Bakterie Gram-ujemne nie tworzą przetrwalników, podczas gdy niektóre Gram-dodatnie tak.
Ściana komórkowa prokariontów spełnia różnorodne funkcje: mechanicznie chroni komórkę przed wpływami środowiska, utrzymuje jej zewnętrzny kształt i umożliwia komórce istnienie w roztworach hipotonicznych. Przede wszystkim jest to „zasługa” peptydoglikanu. Przepuszczalność i transport substancji do wnętrza komórki jest związany ze ścianą komórkową.
Pod błoną cytoplazmatyczną bakterii znajduje się cytoplazma. Jest to układ koloidalny składający się z wody, białek, tłuszczów, węglowodanów, związków mineralnych i innych substancji, których stosunek zmienia się w zależności od rodzaju bakterii i ich wieku. Cytoplazma komórki prokariotycznej nie podlega cyklicznym ruchom, jak u roślin, ani zjawiskom skurczu, jak u ameby. komórki bakteryjne wykonują ruch dzięki innym mechanizmom: albo za pomocą po prostu ułożonych wici, które różnią się od złożonej wici eukariontów, aktywnie pływają w ciekłym ośrodku lub ślizgają się, wydzielając śluz i poruszając się po powierzchni gęstego podłoża.
Ruchliwe komórki bakterii aktywnie poruszają się w kierunku określonym przez określone czynniki. Takie ukierunkowane ruchy bakterii nazywane są „taksówkami”. Istnieje chemotaksja, fototaksja, magnetotaksja, termotaksja, wiskozytaksja (reakcja na zmianę lepkości roztworu).
Spoczynkowe komórki bakteryjne są reprezentowane w bakteriach Gram-dodatnich przez endospory, w bakteriach Gram-ujemnych - przez cysty. Zarodniki bakterii mają wyjątkową odporność na działania niepożądane. Szczególnie uderzająca jest odporność zarodników bakterii na ciepło: niektóre zarodniki tolerują gotowanie przez 1 godzinę lub dłużej i nie tracą żywotności. Bardzo odporny na zarodniki i wysychanie. Stabilność zarodników związana jest z obecnością specjalnej warstwy w ich ścianie komórkowej - kory.
Podczas tworzenia zarodników komórki bakteryjne czasami przybierają nietypowy kształt wrzeciona, cytryny lub pałeczka do gry na bębnie. Przetrwalnik jest zwykle jeden na komórkę, ale istnieją bakterie z więcej niż czterema przetrwalnikami. Niektóre bakterie beztlenowe na zarodnikach tworzą się czapeczki komórkowe, zbudowane z wakuoli gazowych lub różnego rodzaju narośli
Zarodniki bakterii nie są jedynymi formami spoczynku u prokariotów. Bakterie odczytujące Grama, które nie są zdolne do tworzenia przetrwalników, w niektórych przypadkach zamieniają się w cysty - specjalne komórki z grubymi powłokami zewnętrznymi (np. , Azotobacter chroococcum)
Na zewnątrz ściany komórkowe prokariotów są często otoczone substancją śluzową. Takie formacje, w zależności od cech strukturalnych, nazywane są kapsułkami, warstwami śluzowymi, osłonkami lub osłonkami (u bakterii nitkowatych). Wszystkie są wynikiem biosyntezy polimerów organicznych przez prokarioty i ich osadzania się wokół komórek.
Kapsułki, formacje śluzowe i otoczki mogą zawierać składniki, które są takie same jak ściana komórkowa, ale skład chemiczny nieidentyczny. Z reguły skład chemiczny kapsułek tworzonych przez bakterie jest specyficzny dla rodzaju i gatunku. Głównymi składnikami chemicznymi większości kapsułek prokariotycznych są polisacharydy. Wyjątkiem są otoczki niektórych rodzajów prątków, zbudowane z polipeptydu, który jest polimerem kwasu glutaminowego. Sprawy - więcej złożone struktury, mają bardziej złożony skład chemiczny i zawierają cukry, białka, lipidy.
Kapsułki mają różne grubości i można je łatwo zidentyfikować
pod mikroskopem z negatywnym kontrastem preparatu czarnym tuszem.
Kapsułki i otoczki chronią komórkę przed uszkodzeniem i wysychaniem, tworzą dodatkową barierę osmotyczną, stanowią przeszkodę w przenikaniu fagów, tworzą wokół komórek specjalne środowisko, które zapewnia pewną reżim wodny. Kapsułki mają określone funkcje. Na przykład w bakteriach brodawkowych decydują o selektywnym przyczepianiu się do włośników roślin różnych gatunków. Czasami kapsułki mogą być używane jako rezerwowe składniki odżywcze. Za pomocą śluzu odbywa się komunikacja między komórkami w kolonii, przyczepianie komórek do różnych powierzchni
Polisacharydy otoczkowe oddziałują z kationami roztworów glebowych, mogą niszczyć minerały glebowe, uczestniczyć w procesach agregacji i tworzenia wodoodpornej struktury gleby. Być może włączenie mikrobiologicznego śluzu zewnątrzkomórkowego w proces powstawania substancji humusowych.
Kształt komórek prokariotów jest bardzo zróżnicowany. Znane prokarioty są kuliste, w kształcie pręta, spiralnie skręcone, w formie przecinka, w kształcie pierścienia - zamkniętego lub otwartego, z wypustkami - proste. Komórki mogą być kwadratowe lub w kształcie gwiazdy. Wielokomórkowe prokarioty mają kształt włókien lub robaków.
Wszystkie prokarionty należą do grupy organizmów mikroskopijnych, wielkość ich komórek mierzona jest w mikronach (mikrometrach) (1 μm = 10 3 mm), a ich szczegóły mierzone są w nanometrach (1 nm = 10 mm). Prawie 40% bakterii w glebie jest poza możliwościami mikroskopu świetlnego, widać je tylko w mikroskopie elektronowym. Przy tak małych rozmiarach stosunek powierzchni do objętości jest bardzo wysoki, co skutkuje wysokim potencjalnym tempem metabolizmu.
Wszystkie prokarionty dzielą się na cztery grupy w zależności od budowy zewnętrznej powłoki komórki: Gram-ujemne (Gracilicutes) eubakterie, Gram-dodatnie (Firmicutes) eubakterie, mykoplazmy (Mollicutes), pozbawione ścian komórkowych oraz archebakterie, czyli archeony (Archaebakterie). Pierwsze trzy grupy są zjednoczone jako eubakterie (Eubacteria), archebakterie to odrębna grupa organizmów (Archaebacteria), które różnią się od innych strukturą rybosomalnych i transferowych cząsteczek RNA, polimeraz RNA, lipidów i strukturą ściany komórkowej.
Bakterie Gram-ujemne posiadają ściany komórkowe, w których oprócz głównej warstwy szkieletowej mureiny znajduje się również błona zewnętrzna oraz warstwa białka i/lub glikoproteiny nachodząca na nią od góry. Pomiędzy błoną cytoplazmatyczną a warstwą mureiny znajduje się przestrzeń peryplazmatyczna. Ta grupa bakterii jest najbardziej rozbudowana i zróżnicowana pod względem liczby gatunków i funkcje fizjologiczne. Grupa prokariotów obejmuje bakterie fototroficzne i chemotroficzne. Należą do nich pospolite saprotrofy, kopiotrofy, oligotrofy, wewnątrzkomórkowe pasożyty innych bakterii, formy chorobotwórcze dla ludzi, zwierząt i roślin o złożonych potrzebach pokarmowych. Pod względem morfologicznym są to różnorodne bakterie, od prostych pseudomonad w kształcie pałeczek po myksobakterie o złożonym cyklu rozwojowym.
Mykoplazmy to bakterie, które nie mają ściany komórkowej, której funkcję pełni błona cytoplazmatyczna.
Archebakterie zrzeszają prokarionty o odmiennej budowie zewnętrznej powłoki, w której brakuje charakterystycznej dla prawdziwych bakterii mureiny. Niektórzy przedstawiciele archebakterii mają pseudomureinę w ścianie komórkowej, podczas gdy inni mają ścianę komórkową złożoną z warstwy białek kulistych lub glikoprotein.
Podział bakterii na grupy na podstawie cech fenotypowych (morfologia, fizjologia) jest wygodny ze względów praktycznych do szybkiej identyfikacji organizmów. Organizmy są pogrupowane w sztuczne grupy lub „sekcje”, które nie mają statusu taksonomicznego i często są określane na podstawie ich najbardziej charakterystyczny rodzaj lub tytuł. Zgodnie z tą zasadą budowany jest wyznacznik bakterii Burgeya.
Badając różnorodność organizmów, które zamieszkiwały i zamieszkują Ziemię, taksonomowie dążą do stworzenia systemu filogenetycznego, który odzwierciedlałby pokrewieństwo genealogiczne i wspólne pochodzenie organizmów zjednoczonych w jednej lub drugiej grupie - taksonie. Temu celowi służy systematyka molekularna, oparta na podobieństwie budowy biopolimerów - DNA, RNA, białek. Ze względu na możliwość zidentyfikowania podobieństwa genów często nazywa się to systematyką genów. Jednym z najważniejszych osiągnięć systematyki genów jest odkrycie nowej grupy mikroorganizmów wyizolowanych w odrębnym królestwie – archebakterii. Stopień podobieństwa genomów bakteryjnych określa się porównując sekwencje nukleotydowe w cząsteczce 16S rRNA. Zastosowanie tej metody w taksonomii bakterii posłużyło za podstawę do skonstruowania drzewa filogenetycznego bakterii przez K. Wese.
Bardzo trudno jest porównać systematykę filogenetyczną i fenotypową. Te same gałęzie filogenetyczne zawierają organizmy niezwykle zróżnicowane pod względem fizjologii. Przechodząc z dużych gałęzi bakterii Gram-dodatnich i Gram-ujemnych na niższe poziomy hierarchii, takich niedopasowań jest coraz więcej.
PRACA KURSU
w biologii
„Pochodzenie eukariota
Ukończone przez: Severova Vera
10 klasa, szkoła N55
Sprawdzone przez: Strukova N.A.
Ryazan 1999
1. Teoria symbiotyczna:
1.1 Podstawowe rozróżnienie między prokariontami a eukariontami;
organizmy prokariotyczne;
komórki eukariotyczne;
Mitochondria i plastydy;
Zdolność eukariontów do fotosyntezy;
2. Prekursorzy życia;
3. Organizmy fermentujące:
Źródła energii wczesnych komórek;
Metanogeneza jako źródło energii;
Cyjanobakterie i chlorooksybakterie;
Fotosynteza i powietrze;
Skojarzenia i eukarionty:
Teorie kolejnych symbioz;
Symbiotyczne nabywanie mobilności;
Eukarionty są fotosyntetyzujące;
Spojrzenie na ewolucję komórki:
6.1 Hipoteza bezpośredniego synostwa;
Hipoteza: od sinic do alg;
Zgodność 2 hipotez: bezpośredniego synostwa i
endosymbioza;
Literatura.
TEORIA ENDOSYMBIOZY Wszystkie żywe organizmy na Ziemi dzielą się zwykle na prokarioty i eukarionty. Główną cechą prokariontów, w przeciwieństwie do eukariontów, jest brak pełnoprawnego jądra komórkowego pokrytego błoną. Komórki prokariotyczne są bardzo małe, rzędu 1 µm. Objętość komórek eukariotycznych zawierających pełnoprawne jądro jest 800-1000 razy większa niż objętość komórek prokariotycznych. Pod tym względem komórki - eukarionty potrzebują znacznie większego aparatu błonowego do wykonywania funkcji życiowych. Rzeczywiście, w komórce eukariotycznej zidentyfikowano struktury subkomórkowe otoczone błonami niezależnymi od plazmalemmy i od siebie nawzajem. Struktury te nazywane są organellami. Każda organella: mitochondria, chloroplast, jądro, lizosomy, aparat Golgiego itp. ma swoje specyficzne funkcje w życiu komórki. Za pomocą podejść molekularno-biologicznych uzyskano przekonujące dowody na to, że wszystkie żywe istoty należy podzielić na 3 królestwa: bakterie, archeony i eukarionty. Bakterie i archeony powstały na Ziemi od wspólnego przodka – tzw. progenota – około 4 mld lat temu, a według paleontologów eukarionty pojawiły się na Ziemi 500 mln lat po prokariotach. Najbardziej popularną i wystarczająco uzasadnioną koncepcją pochodzenia komórek eukariotycznych jest obecnie idea wielokrotnej endosymbiozy różnych prokariotów. Według znanych mikrobiologów A. Kluyvera i K. Van Niela to właśnie u starożytnych prokariotów następowała selekcja komórek o najbardziej ekonomicznych i często wielofunkcyjnych reakcjach biochemicznych, które stały się podstawą najważniejszych szlaków metabolicznych.
Symbiotyczna teoria pochodzenia i ewolucji komórek opiera się na dwóch nowych dla biologii koncepcjach. Zgodnie z pierwszą z nich najbardziej fundamentalnym rozróżnieniem w żywej przyrodzie jest rozróżnienie między prokariotami i eukariontami, tj. między bakteriami a organizmami składającymi się z komórek z prawdziwym jądrem – protistami, zwierzętami, grzybami i roślinami. Druga koncepcja mówi, że niektóre części komórek eukariotycznych powstały w wyniku ewolucji symbioz, tj. tworzenie trwałych związków między organizmami różnych gatunków. Przyjmuje się, że trzy klasy organelli – mitochondria, rzęski i fotosyntetyzujące plastydy – wywodzą się od wolno żyjących bakterii, które w wyniku symbiozy zostały włączone w określonej kolejności do komórek gospodarzy prokariotycznych. Teoria ta w dużej mierze opiera się na neodarwinowskich ideach opracowanych przez genetyków, ekologów, cytologów i innych naukowców, którzy powiązali genetykę mendlowską z darwinowską ideą doboru naturalnego.Opiera się również na zupełnie nowych lub niedawno odrodzonych kierunkach naukowych: na biologii molekularnej , zwłaszcza na danych dotyczących struktury białek i sekwencji aminokwasów, na mikropaleontologii, która bada najstarsze ślady życia na Ziemi, a nawet na fizyce i chemii atmosfery, ponieważ te nauki są związane z gazami pochodzenia biologicznego. Wszystkie organizmy komórkowe można podzielić na pięć królestw: królestwo prokariotyczne (Monera, do którego należą bakterie) i cztery królestwa eukariotyczne (Protoctista, Animalia, Fungi, Plantae). Protoktyści to organizmy eukariotyczne, które nie są zwierzętami, grzybami ani roślinami. Królestwo protoktystów obejmuje algi, pierwotniaki, śluzowce i inne organizmy eukariotyczne niejasnego pochodzenia. Protisty definiuje się bardziej restrykcyjnie jako jednokomórkowe eukarionty. Tak więc królestwo Protoctista obejmuje nie tylko protisty - jednokomórkowe eukarionty, ale także ich najbliższych wielokomórkowych potomków, takich jak algi czerwone i brunatne, a także wiele mikroorganizmów podobnych do grzybów, takich jak chytridy. Zgodnie z tradycyjnym poglądem na bezpośrednie filiowanie, organelle komórkowe, takie jak mitochondria i plastydy, powstały w wyniku kompartmentalizacji samej komórki.
KILKA PROKARIONTÓW TWORZY JEDNEGO EUKARIONTA
Komórki są układami związanymi z błonami, najmniejszymi jednostkami biologicznymi zdolnymi do samoreprodukcji. Nawet najmniejsze komórki zawierają geny w postaci cząsteczek DNA (jednej lub więcej) oraz aparatu do syntezy białek, składającego się z kilku rodzajów RNA i wielu białek. Wszystkie komórki zawierają rybosomy, ciała o średnicy około 0,02 µm, zbudowane z co najmniej trzech rodzajów RNA i około pięćdziesięciu różnych białek. Organizmy prokariotyczne to najmniejsze jednostki biologiczne, które spełniają tę definicję komórki. Nie mają jąder. Z drugiej strony pojedyncze komórki eukariotyczne (posiadające jądro) w pewnym sensie wcale nie są jednostkami. Są to kompleksy różnych jednostek syntetyzujących białka (nukleocytoplazma, mitochondria i plastydy), których typ i liczba różni się w zależności od gatunku. Tak więc, zgodnie z tą teorią, komórki eukariotyczne powstały w wyniku współpracy początkowo niezależnych elementów, połączonych w pewien porządek. Taylor nazwał tę ideę odrębnego pochodzenia i rozwoju części komórki eukariotycznej oraz ich późniejszego zjednoczenia teorią kolejnych endosymbioz. Warunki wysokiej temperatury i kwasowości doprowadziły do wytworzenia w nukleocytoplazmie specjalnej klasy białek - histonów - w celu ochrony jej DNA. Mitochondria były w stanie rozłożyć związki trójwęglowe na dwutlenek węgla i wodę. Symbiotyczny związek nukleocytoplazmy i mitochondriów zaczął się zmieniać w kierunku wzmocnienia ich wzajemnej zależności. W skład tego kompleksu wchodziły organelle ruchowe - rzęski i wici. Po tej transformacji nastąpiła transformacja morfogenetyczna, która polegała na rozwoju wyraźnych asymetrii komórkowych. Wraz z integracją współzależnej nukleocytoplazmy, mitochondriów, undulipodiów zakończono tworzenie struktury poligenomicznej komórki eukariotycznej. Pierwsza komórka eukariotyczna była heterotroficzna. Jadała gotowe związki organiczne. U tych eukariontów powstały procesy mitozy, a następnie mejozy. I wreszcie nabycie przez eukarionty zdolności do fotosyntezy nastąpiło w procesie stawania się główną organizacją eukariotyczną, a po jego zakończeniu – ostatnim wydarzeniem w tej epopei. Symbiozy stawały się coraz bardziej zintegrowane. Obecnie zależność każdej organelli od produktów metabolicznych innych organelli stała się tak całkowita, że tylko nowoczesne metody analizy umożliwiają prześledzenie szlaków metabolicznych pierwotnych partnerów. Eukarionty różnią się budową, ale metabolicznie pozostają jednolite. Metaboliczne sztuczki eukariontów zostały opracowane przez różne bakterie jeszcze przed czasem, kiedy połączyły się, by stać się żywicielem i jego organellami. Prokarionty osiągnęły wielką różnorodność w sposobie pozyskiwania energii i zawiłościach biosyntezy. Pozostawiły ślad swojego istnienia na długo przed tym, zanim z komórki eukariotycznej powstały tak duże organizmy jak zwierzęta ze szkieletem.
Schemat pochodzenia komórek eukariotycznych przez symbiozę
PREKURSORZY ŻYCIA
Przodkowie dzisiejszych organizmów powstali ponad trzy miliardy lat temu ze związków organicznych powstałych na powierzchni Ziemi. Atrakcyjność tej koncepcji wynika częściowo z jej testowalności.
S. Miller i H. Urey wykazali, że w warunkach atmosferycznych, jakie miały miejsce na młodej Ziemi, mogą powstawać aminokwasy i inne cząsteczki organiczne. Różne typy cząsteczek organicznych powstały spontanicznie w ciągu pierwszego miliarda lat istnienia Ziemi. W przestrzeni międzygwiezdnej iw meteorytach znaleziono wiele cząsteczek organicznych. Oznacza to, że związki organiczne powstają podczas interakcji lekkich pierwiastków, w obecności źródeł energii. Uniwersalność chemicznego mechanizmu bioreprodukcji oznacza, że całe życie na Ziemi pochodzi od wspólnych przodków: całe obecne życie wywodzi się z komórek z systemami informacyjnymi opartymi na replikacji DNA i syntezie białek kierowanej przez informacyjne RNA.
FERMENTUJĄCE ORGANIZMY
Możliwe, że kwasy nukleinowe od samego początku były oddzielane od środowiska przez membrany lipoproteinowe. Informacje zaczęły być kodowane przez sekwencję nukleotydów w cząsteczkach DNA. Najstarsze komórki wykorzystywały ATP i aminokwasy, które są częścią białek, jako bezpośrednie źródła energii i jako składniki strukturalne. Tak więc - najstarsze życie istniało w postaci bakterii heterotroficznych, które otrzymywały żywność i energię z materiału organicznego pochodzenia abiotycznego. Neodarwinowska ewolucja organiczna wymaga trzech zjawisk: reprodukcji, mutacji i selektywnej presji środowiskowej. Rozmnażanie jest niemożliwym do zastosowania warunkiem ewolucji biologicznej. Przy wystarczająco dokładnej reprodukcji mutacje i dobór naturalny są nieuniknione. Wyjaśnienie pochodzenia takiej reprodukcji oznacza zrozumienie pochodzenia samego życia. Mutacja, główne źródło zmienności w ewolucji, prowadzi do dziedzicznych zmian w samoreprodukującym się systemie. Po opracowaniu niezawodnej reprodukcji rozpoczęła się ewolucja neodarwinowska. Organizmy, które rozwinęły szlaki syntezy składników potrzebnych komórce, miały przewagę nad innymi. Bakterie beztlenowe wytwarzające metan były najstarszymi autotrofami, czyli organizmami zdolnymi do zaspokajania swoich potrzeb w zakresie energii i substancji organicznych ze źródeł nieorganicznych. Zdolność do fotosyntezy została po raz pierwszy rozwinięta u bakterii wrażliwych na tlen. Pochłaniając światło słoneczne za pomocą bakteryjnego chlorofilu, przekształcili dwutlenek węgla z atmosfery w substancje organiczne.Pojawienie się fotosyntezy beztlenowej nieodwracalnie zmieniło powierzchnię i atmosferę Ziemi.
Dwutlenek węgla został usunięty z atmosfery i przekształcony w materię organiczną zgodnie z ogólnym równaniem fotosyntezy: 2H2X+CO2=CH2O+2X+H2O. Bakterie fotosyntetyczne, które wykorzystywały siarkowodór do redukcji CO2 osadzonej siarki elementarnej. Dały początek bakteriom zdolnym do fotosyntezy z uwalnianiem tlenu. Ten rodzaj fotosyntezy powstał prawdopodobnie w wyniku nagromadzenia mutacji w fotosyntetycznych bakteriach siarkowych. Droga ta doprowadziła do pojawienia się sinic lub sinic. Niedawno odkryto i zbadano nową grupę fotosyntetycznych prokariontów wytwarzających O2, chlorooksybakterie. Można przypuszczać, że przodkami plastydów występujących w komórkach alg i roślin były cyjano- i chlorooksybakterie. Gdy w wyniku mikrobiologicznej fotosyntezy gromadził się O2, atmosfera ziemska zaczęła się utleniać, co spowodowało kryzys wszelkiego życia – do tego czasu świat zamieszkiwały bakterie beztlenowe. Aby przetrwać, drobnoustroje musiały unikać tlenu, ukrywając się na bagnach, lub musiały rozwinąć metabolizm zdolny do ochrony przed rosnącą ilością tlenu.
FOTOSYNTEZA I POWIETRZE
Około 2 miliardów lat temu, jeden po drugim, miały miejsce różne procesy ewolucyjne, które stworzyły różnorodność metaboliczną. Zdolność do wytwarzania określonych enzymów może być przenoszona w obrębie tego samego gatunku za pomocą cząsteczek wirusopodobnych zwanych plazmidami. Zdolności biosyntetyczne prokariontów są uderzające: z dwutlenku węgla, kilku soli i zredukowanych związków nieorganicznych, które służą jako źródła energii, mikroorganizmy chemoautotroficzne mogą syntetyzować w nieoświetlonej wodzie zawierającej tlen wszystkie najbardziej złożone makrocząsteczki niezbędne do wzrostu i rozmnażania. Te procesy biochemiczne doprowadziły do powstania układów krążenia pierwiastków nawet wtedy, gdy nie było roślin, grzybów, zwierząt.
asocjacje i eukarionty.
Spojrzenie na ewolucję komórki.
Oddział bezpośredni.
Tradycyjny pogląd na pochodzenie i ewolucję komórek eukariotycznych jest taki, że wszystkie żywe organizmy pochodzą bezpośrednio od jednej populacji przodków w wyniku nagromadzenia mutacji pod wpływem doboru naturalnego.
Mutacje punktowe, duplikacje, delecje i inne zmiany dziedziczne, o których wiadomo, że odegrały rolę w ewolucji zwierząt i roślin, są również odpowiedzialne za różnicowanie komórek eukariotycznych od prokariotycznych - teoria bezpośredniego filiowania. tj. pojęcie niesymbiotycznego pochodzenia komórek. Być może zielone algi są potomkami przodków wszystkich roślin. Wiele cech tych glonów sugeruje, że one i rośliny mają wspólnego przodka. Filogeneza zwierząt jest bardziej złożona, ale koncepcja bezpośredniego pokrewieństwa prawdopodobnie odnosi się również do ich ścieżek ewolucyjnych. Gąbki najwyraźniej wyewoluowały z protistów, niezależnie od innych grup zwierząt. Tak więc, przynajmniej w przypadku zwierząt i roślin, hipoteza, że złożone organizmy eukariotyczne wyewoluowały z prostszych eukariotów poprzez akumulację indywidualnych mutacji pod wpływem doboru naturalnego, jest całkiem uzasadniona. Innowacje ewolucyjne polegają na zachowaniu wysoce doskonałych korzystnych genów. Tak więc nowe organizmy o zwiększonej zdolności do kontrolowania swojego środowiska ewoluowały bardziej dzięki utrzymywaniu duplikacji i rekombinacji wysoce przystosowanych genów niż nabywaniu nowych właściwości poprzez proste, losowe mutacje punktowe. Hipoteza bezpośredniego synostwa nie wyjaśnia pochodzenia eukariontów od prokariotów. Na przykład przerwa między sinicami niezdolnymi do mitozy a takimi formami z w pełni rozwiniętą mitozą, jak krasnorosty, jest trudna do zrozumienia z punktu widzenia bezpośredniego filiowania, ale naturalnie wynika z teorii symbiozy.
Zgodnie z teorią bezpośredniego potomstwa i teorią symbiozy wszystkie organizmy na Ziemi wywodzą się od biochemicznie podobnych przodków bakterii, a drobnoustroje eukariotyczne dały początek roślinom i zwierzętom. Bakterie fotosyntetyzujące dały początek glonom, a ostatecznie roślinom, a niektóre glony utraciły plastydy i stały się przodkami grzybów i zwierząt. Teoretycy bezpośredniego filiowania zgadzają się, że plastydy i inne organelle eukariotyczne, w tym jądro, powstają w wyniku różnicowania w samych komórkach i nazywają to mitem botanicznym.
Filogeneza typów roślin
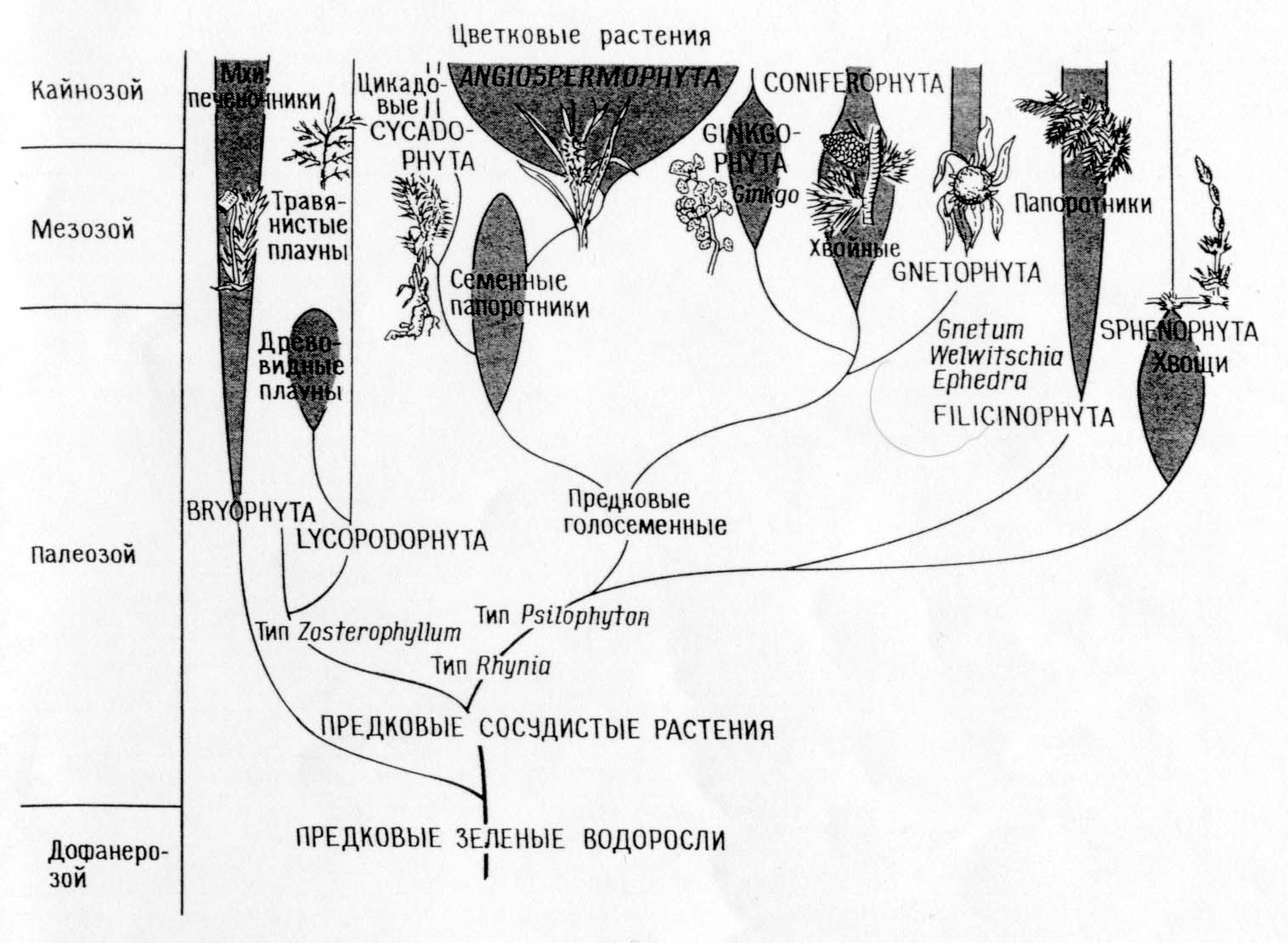
Porównanie filogenez odpowiadających teorii bezpośredniego synostwa (A)
i teoria symbiozy (B)
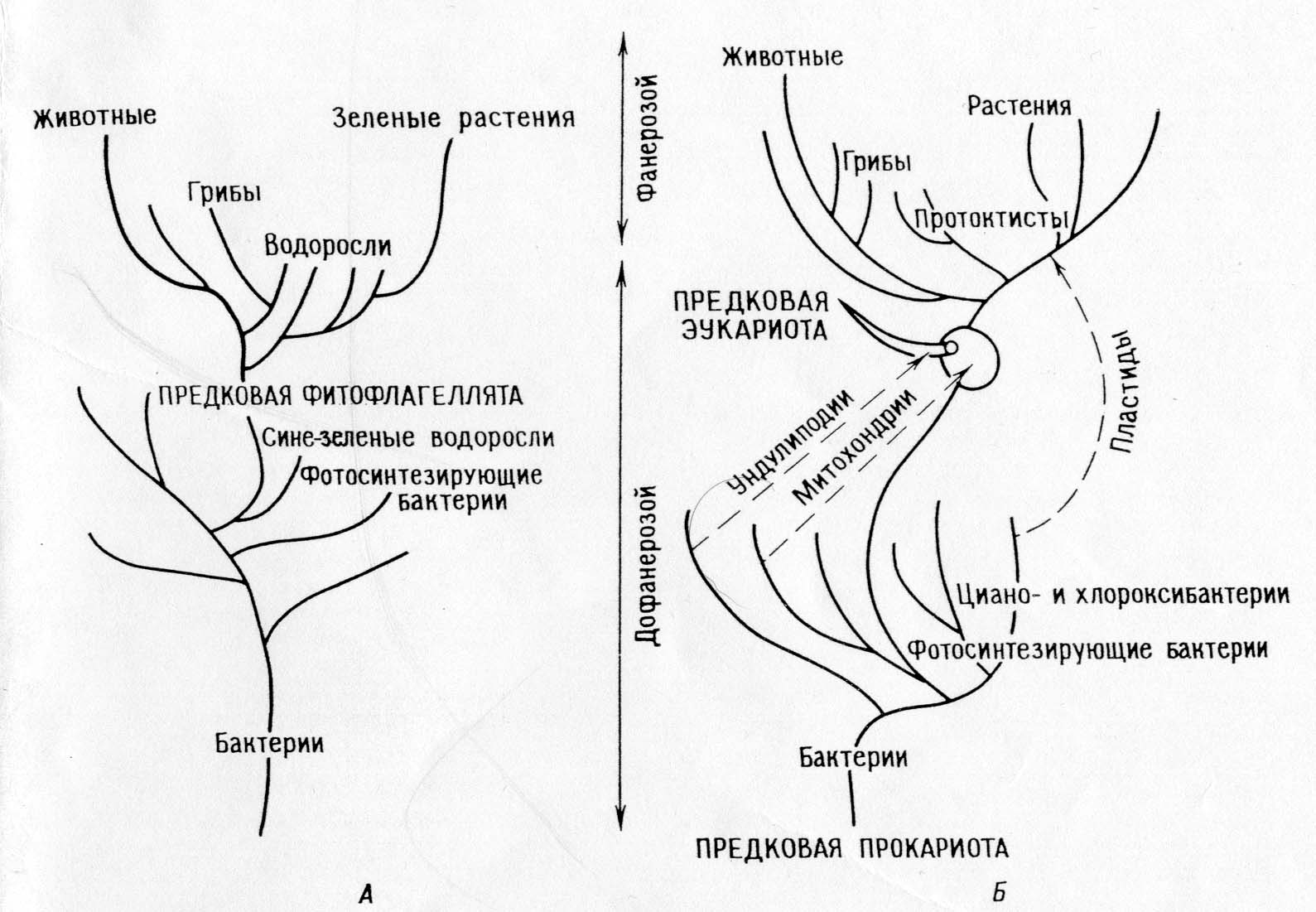
Różnice między teorią bezpośredniego synostwa a teorią symbiozy:
Teoria bezpośredniego synostwa.
Główna dychotomia: zwierzęta - rośliny
Fotosyntetyczne eukarionty (algi i rośliny zielone) wyewoluowały z fotosyntetycznych prokariontów (niebiesko-zielone algi)
Rośliny i ich systemy fotosyntetyczne ewoluowały monofiletycznie w prekambrze.
Zwierzęta i grzyby wyewoluowały z autotrofów, które utraciły plastydy.
Mitochondria różnicowały się wewnątrzkomórkowo w algach fotosyntetyzujących.
W chwastach prawoskrętnych unlipodia, aparat mitotyczny i wszystkie inne organelle eukariotyczne różnicowały się wewnątrzkomórkowo.
Wszystkie organizmy wywodzą się od swoich przodków – prokariontów poprzez nagromadzenie pojedynczych mutacji.
KONSEKWENCJE: Zgodnie z teorią muszą istnieć prawe chwasty; trudno jest powiązać tę teorię z zapisem kopalnym i skonstruować odpowiednią filogenezę.
Teoria kolejnych endosymbioz:
Główna dychotomia: prokarioty - eukarionty.
Fotosyntetyzujące eukarionty (algi jądrowe i rośliny) oraz niefotosyntetyzujące eukarionty (zwierzęta, grzyby, pierwotniaki) wyewoluowały ze wspólnych heterotroficznych przodków (ameboflagellaty).
Ewolucja fotosyntezy u Archaean była monofiletyczna u bakterii (w tym cyjano- i chlorooksybakterii), protisty fotosyntetyzujące powstały nagle, gdy ich heterotroficzni protisty-przodkowie weszli w symbiozę z fotosyntetycznymi prokariotami (protoplastidami).
Zwierzęta oraz większość glonów i grzybów pochodzi bezpośrednio od heterotroficznych protistów.
Mitochondria były już obecne u protistów heterotroficznych, kiedy te ostatnie nabyły plastydy w wyniku symbiozy. Promitochondria były wcześniej nabywane w sposób symbiotyczny.
Mitoza rozwinęła się u heterotroficznych protistów eukariotycznych poprzez różnicowanie układu unlipodialnego.
Prokarioty ewoluowały od wczesnych prokariotycznych przodków poprzez serię pojedynczych zdarzeń mutacyjnych, eukarioty również ewoluowały od eukariotycznych przodków poprzez pojedyncze zdarzenia mutacyjne, a eukariotyczni przodkowie ewoluowali od prokariotów poprzez serię symbioz.
Konsekwencje: Teoria wyjaśnia szlaki biochemiczne i ich redundancję w organellach. Pozwala to na zbudowanie spójnej filogenezy, która wyjaśnia „lukę biologiczną” między proterozoikiem a fanerozoikiem. Teoria przewiduje, że organelle muszą zachowywać pozostałości systemów replikacyjnych.
mit botaniczny
Od czasów Jussieu niższe rośliny nazywane są talofitami, a wyższe mszakami i tracheofitami. Według mitu botanicznego, wspólni przodkowie bakterii fotosyntetyzujących i sinic, poprzez bezpośrednie filiowanie, dali początek glonom eukariotycznym, w tym algom morskim. Do dziedziny botaniki trafiły grupy organizmów, na których filogenezę najbardziej oddziałuje koncepcja bezpośredniej filiacji – są to bakterie, algi, grzyby i rośliny.
Hipotetyczne organizmy, które mogłyby być łącznikiem między sinicami a zielonymi algami, nazwano fitomonadami przodków (fitomonadami przodków) lub prawicowcami. Niektórzy autorzy argumentowali, że eukarionty Cyanidium i Cyanophora, które są podobne w swoich układach barwnikowych do sinic, mogą być odpowiednimi kandydatami do roli prawicowców. U talofitów nie stwierdzono form pośrednich między niemitotycznym stanem cyjanobakterii a w pełni rozwiniętą typową mitozą zielonych alg i roślin. Chociaż grzyby i rośliny kwitnące są całkowicie pozbawione undulipodiów, charakteryzują się mitozą i mejozą, które wymagają wrzecion mikrotubul. Istnieje powszechna zgoda co do tego, że wszystkie organizmy wielokomórkowe z mikrotubulami pochodzą od mikroorganizmów eukariotycznych z undolipodiami, ale nikt nie wie, z jakiej grupy protistów. Wzmocnieniem mitu botanicznego była starożytność i złożoność fotosyntezy, a także fakt, że wiele organizmów (bakterie, euglenoidy, rośliny pasożytnicze), zarówno w naturze, jak iw laboratorium, miało utratę fotosyntezy w wyniku mutacji. Ponieważ fotosynteza jest procesem beztlenowym, który ostatecznie zapewnia istnienie wszystkich organizmów, musiała rozwinąć się bardzo wcześnie w historii życia na naszej planecie, jeszcze zanim pojawiła się mitoza, która wymaga tlenu. w fotosyntezie bierze udział wiele pigmentów, lipidów i enzymów, uporządkowanych w bardzo złożonych błonach. Jednocześnie szczegóły mechanizmów fotosyntezy u alg i roślin z jednej strony oraz u cyjanobakterii z drugiej są uderzająco podobne. Wszystkie te organizmy wydzielają gazowy tlen z wody, który służy jako źródło atomów wodoru wykorzystywanych do redukcji CO2 do materii organicznej komórki. Wiele glonów i pierwotniaków nie ma ani mitozy, ani systemu genetycznego podobnego do zwierząt i roślin. Orzęski mają specjalny system genetyczny z 2 typami jąder, a komórki są często wielojądrowe. Jeden rodzaj jąder to makrojądra, a drugi to znacznie mniejsze mikrojądra. Mikrojądra są fizjologicznie nieistotne. przechowują jednak kopie genów, a podczas procesu płciowego przechodzą mejozę. Fizjologicznie niezbędne makrojądra służą jako miejsce transkrypcji RNA, a tym samym rządzą fizjologią komórki. Jedną z trudności w wyjaśnieniu związku między sinicami a algami eukariotycznymi jest brak danych na temat pochodzenia undolipodiów: nie znaleziono pośrednich powiązań między sinicami, które nigdy nie mają undolipodiów, a wieloma algami, które je mają. Innym problemem jest całkowity brak procesu płciowego u cyjanobakterii: między sinicami bezpłciowymi a algami, z których większość charakteryzuje się procesem płciowym, nie było form pośrednich. Stwierdzono, że większość alg ma unlipodia na pewnym etapie ich cyklu życiowego. Ruchome struktury glonów, pierwotniaków, zwierząt i roślin okazały się takie same: wszystkie składają się z mikrotubul o średnicy 0,024 mikrona. Okazało się, że wrzeciono mitotyczne, nawet u grzybów nieposiadających undolipdiów, składa się z białkowych mikrotubul o takiej samej średnicy jak u undolipdiów. Przez pół wieku wydawało się niezwykle prawdopodobne, że glony eukariotyczne wyewoluowały z sinic. Ten botaniczny mit doprowadził do założeń, które przez wiele lat służyły zarówno jako bodziec do badań, jak i punkt odniesienia, w którym oceniano wyniki. Twierdzenie, że powiązania między sinicami a eukariotycznymi algami wymarły, nie pozostawiając żadnych śladów skamielin i żywych reliktów, stawało się coraz mniej przekonujące w miarę pojawiania się nowych danych na temat żywych drobnoustrojów i skamieniałości. Główne ogniwa pośrednie między fotosyntetyzującymi prokariotami a eukariontami nie wymarły bez śladu, jak tego wymaga mit botaniczny – nigdy nie istniały. Zgodnie z teorią kolejnych endosymbioz, sinice są wprawdzie przodkami, ale tylko plastydami, a nie resztą komórki eukariotycznej, a już na pewno nie jej jądrem. A jednak w kwestii pochodzenia jądra hipoteza bezpośredniego synostwa i teoria symbiozy są zgodne. Możliwe do pomyślenia modyfikacje skrajnej wersji teorii symbiozy obejmują na przykład ideę symbiotycznego pochodzenia plastydów, ale nie mitochondriów, lub symbiotycznego pochodzenia plastydów i mitochondriów, ale nie unlipodiów. Zwolennicy bezpośredniego synostwa wysunęli również założenie o pochodzeniu jądra poprzez symbiozę.
Krytyczne dowody na endosymbiotyczne pochodzenie chloroplastów i mitochondriów.
Najbardziej znaczący dowód pochodzenia najważniejszych organelli komórek eukariotycznych - chloroplastów i mitochondriów z prokariotów uzyskano w ostatnich latach, badając strukturę rybosomalnego RNA. Biolog molekularny
Już w latach 80. K. Woz wykazał, że budowa RNA z rybosomów chloroplastów roślinnych jest niemal identyczna z budową RNA niektórych sinic. Jednocześnie grupa naukowców odkryła, że RNA rybosomów pochodzących z mitochondriów różnych eukariontów jest podobne do RNA niektórych bakterii, w szczególności bakterii z rodzaju Paracoccus. Zestaw enzymów oddechowych tej bakterii jest bardzo podobny do
„zespół oddechowy” zwierzęcych mitochondriów.
Dane te dość jednoznacznie świadczą o większym prawdopodobieństwie endosymbiotycznego pochodzenia komórek eukariotycznych. Zjawisko endosymbiozy jest nadal niezwykle rozpowszechnione w przyrodzie. Wiadomo, że bakterie świecące często osadzają się na ciele niektórych ryb morskich, które nie tylko odnoszą ogromne korzyści dla własnego życia, ale są również bardzo przydatne dla organizmów żywicielskich. Wszystko to w przekonujący sposób świadczy o endosymbiotycznym pochodzeniu komórek eukariotycznych ze znacznie mniejszych komórek prokariotycznych. Jednak głównym pytaniem jest ustalenie charakteru komórki gospodarza, ponieważ komórki eukariotyczne mają dobrze uformowane jądro z otoczką, podczas gdy prokarioty takiego jądra nie mają. L. Margelis uważa na przykład, że rolę komórek gospodarza mogły pełnić prokarioty - poprzednicy współczesnych bakterii, należące do mykoplazm - prokarioty bardzo prymitywnie zbudowane, praktycznie pozbawione ścian komórkowych. Niedawno pojawiły się prace japońskiego naukowca, które są szeroko dyskutowane.
T. Oshima. Opierając się na dokładnym badaniu struktury rybosomalnego RNA wielu pro- i eukariontów, w tym wielu przedstawicieli mało zbadanego dotąd superkrólestwa archeonów, autor ten dochodzi do wniosku, że komórka gospodarza była jednym z przodków współczesnych archeonów. Trzeba powiedzieć, że przedstawiciele tego superkrólestwa, w przeciwieństwie do bakterii, mają wiele cech biochemicznych i biologii molekularnej wspólnych z eukariotami. W najnowszych pracach T. Oshimy podaje się argumenty przemawiające za tym, że komórka gospodarza w momencie pojawienia się eukariontów była przodkiem archeonów, podobnie jak jeden ze współczesnych przedstawicieli tych prokariotów - termoplazma. Jakie będzie ostateczne rozwiązanie problemu ustalenia natury komórki gospodarza, która dała początek komórce eukariotycznej w procesie wielokrotnej endosymbiozy, pokażą dalsze badania.
Przedstawiono obecnie najbardziej popularną i potwierdzoną hipotezę pochodzenia komórek eukariotycznych
endosymbioza różnych prokariontów należących do bakterii i archeonów, która wielokrotnie występowała w procesie ewolucji w ciągu pierwszych 1,5–2 miliardów lat istnienia żywych komórek na Ziemi. Decydujące dowody na endosymbiotyczne pochodzenie chloroplastów i mitochondriów uzyskali czołowi światowi biolodzy badający strukturę rybosomalnego RNA. Dane te dość jednoznacznie świadczą o dużym prawdopodobieństwie endosymbiotycznego pochodzenia komórek eukariotycznych ze znacznie mniejszych (o kilka rzędów wielkości) komórek prokariotycznych.
LITERATURA
Dziennik edukacyjny Sorosa. Biologia, Chemia, Nauki o Ziemi, Fizyka, Matematyka. N 5 1998
Margelis L. Rola symbiozy w ewolucji komórek. Za. z angielskiego, M. MIR 1983
Z głównych działów kinetyki studiuje kinetykę reakcji biologicznych; Ta sekcja nazywa się biokinetyka. Biokinetyka jest nauką pionierską, która powstała na przecięciu biochemii i kinetyki chemicznej (patrz ryc. 1). Wybór biokinetyki jako odrębnej dyscypliny nie jest przypadkowy, jest logicznie uzasadniony i wiąże się z wyjątkowym znaczeniem procesów kinetycznych dla wszystkich żywych organizmów. Biokinetyka...
Geny, ale także stworzyły warunki do kumulacji w nich zmian, dywergencji genów i wzrostu różnorodności kontrolowanych przez nie produktów. 2. Ruchome elementy genetyczne Pewną rolę w ewolucji genomów zarówno komórek pro- jak i eukariotycznych odgrywają tzw. ruchome elementy genetyczne - transpozony. Są to autonomiczne jednostki, które niosą nukleotyd...
Kilka możliwe podejścia do rozwoju przyszłości Teoria ogólna Systemy dynamiczne. W rozdziale 10, Okrągłe procesy życia, rozważamy charakterystyczną cykliczną organizację samoorganizujących się systemy rozpraszające. Uogólniony schemat przedstawia charakterystykę generowania cykli transformacyjnych i cykli reakcji katalitycznych, a także hipercykli katalitycznych zgodnie z...