نسالة ، نسالة - التطور التاريخيعالم الكائنات الحية ، في كل من المجموعات التصنيفية العامة والفردية: الممالك ، والأنواع (الأقسام) ، والطبقات ، والأوامر (الأوامر) ، والعائلات ، والأجناس ، والأنواع. تم تقديم مصطلح نسالة من قبل E. Haeckel.
يمكن تمثيل النظام الحديث للحيوانات ، المبني على مبادئ القرابة ، كشجرة أنساب (أو شجرة سلالة) ، مما يعكس المراحل الرئيسية لتطور الحيوان. التكوُّن مجموعات مختلفةدراسة الكائنات الحية بشكل غير متساو. تمت دراسة نسالة الفقاريات (خاصة المجموعات العليا) بشكل أفضل ؛ من اللافقاريات ، تمت دراسة نسالة الرخويات وشوكيات الجلد والمفصليات بشكل أفضل. إن نسالة بدائيات النوى غير مفهومة بشكل جيد.
في قاعدة الشجرة التطورية من الحيوانات أحادية الخلية. في الكائنات أحادية الخلية ، يتم تنفيذ جميع وظائف الجسم بواسطة خلية واحدة. هناك عدة فرضيات حول الأصل حقيقيات النوى أحادية الخليةق: 1) حقيقيات النوى تطورت من بدائيات النوى بالتكافل. تدعم هذه الفرضية حقيقة أن للميتوكوندريا حقيقية النواة والبلاستيدات الخضراء الحمض النووي الخاص بها ؛ 2) ظهرت حقيقيات النوى بسبب تمايز الخلايا بدائية النواة إلى السيتوبلازم والعضيات. تنحدر جميع أنواع البروتوزوا الحديثة من طيور الساركوفلاجيلات القديمة.
الحيوانات متعددة الخلايا ، أكثر تنظيماً ، وفيها وظائف فرديةيتم تنفيذ الكائنات الحية بواسطة خلايا أو أعضاء فردية. تبدأ جميع الكائنات متعددة الخلايا في التطور الفردي بخلية واحدة - الزيجوت ، و المراحل الأوليةيشبه الجنين (بلاستولا) مستعمرة وحيدة الخلية.
هناك عدد من الفرضيات حول أصل الحيوانات متعددة الخلايا. الأكثر شهرة هي فرضيات E. متشنيكوف. وفقًا لنظرية هيكل ، فإن جميع الحيوانات متعددة الخلايا تنحدر من أحد أشكال الأجداد - المعدة. أنا. افترض متشنيكوف أن أسلاف الحيوانات متعددة الخلايا كانت مستعمرة من السوط. أولاً متعددة الخلايالها شكل كروي ، ومغطاة بطبقة من الخلايا مع سوط ، ولديها خلايا أميبية هضمية داخل الجسم ، تطفو بحرية في الماء. أنا. أطلق عليه متشنيكوف اسم البلعمة. استمر التطور الإضافي للكائنات متعددة الخلايا في أربعة اتجاهات. أدى أحد اتجاهات التطور إلى تكوين النوع الرقائقي ، الذي يتكون من طبقة واحدة من الخلايا ويؤدي إلى نمط حياة زاحف. أدى اتجاه آخر للتطور إلى تكوين نوع الإسفنج بطبقتين من الخلايا غير متشكلة في الأنسجة. ليس لديهم جهاز عصبي بعد. هذه حيوانات قاعية غير متحركة. أدى الاتجاه الثالث للتطور إلى تكوين النوع Coelenterates. تجاويف الأمعاء هي بالفعل حيوانات ذات طبقتين (الأديم الظاهر والأديم الباطن) مع أمعاء أولية وفم وتناظر شعاعي وجهاز عصبي منتشر. من بينها حيوانات حرة السباحة وتقود أسلوب حياة متصل. أدى الاتجاه الرابع للتطور إلى تكوين حيوانات ثلاثية الطبقات أكثر تنظيماً. تشكلت أنسجة وأعضاء هذه الحيوانات من ثلاث طبقات من الخلايا (طبقات جرثومية) من الأديم الظاهر والأديم الباطن والأديم المتوسط.
كانت الطبقات الثلاث الأولى حيوانات تشبه مستورقات الأمعاء. ذهب تطور الحيوانات ثلاثية الطبقات في ثلاثة اتجاهات. اتجاه واحد أدى إلى التشكيل الديدان المفلطحة، حيث لا يوجد تجويف للجسم ، ويقع الحمة بين الأعضاء. الديدان المفلطحة هي أول الحيوانات الثنائية (جميع الأنواع اللاحقة لها هذا التناظر). تظهر لأول مرة الجهاز العضلي(سلس)، العقد العصبية. ومع ذلك ، فإن الأمعاء المتكونة تكون مغلقة بشكل أعمى ، ولا تحتوي على فتحة الشرج.
أدى اتجاه آخر إلى تكوين حيوانات تجويف أولية الديدان. تصبح أمعائهم طرفًا إلى طرف ، وتتكون من ثلاثة أقسام بها فتحة الشرج. هناك مزيد من التركيز الخلايا العصبية، وتشكيل حلقة عصبية حول البلعوم. من الشائع في الديدان المفلطحة والديدان الحلقية وجود كيس عضلي جلدي وجهاز هضمي وإخراجي وجهاز تناسلي.
الاتجاه الثالث لتطور الطبقات الثلاث يؤدي إلى تكوين حيوانات فاسدة. وفر تكوين تجويف ثانوي للجسم قدرًا أكبر من الحركة والاستقرار البيئة الداخليةالكائن الحي. تكون التجاويف الثانوية أكثر تنظيماً: لها جهاز دوري. وتشمل هذه جميع اللافقاريات والحبليات الأخرى.
أدى تطور deuterostomes إلى تكوين البروتوستومات و deuterostomes. تطور البروتستومات. نشأت الرخويات والحلقيات الأولية من سلف البروتستومات. في المقابل ، نشأت المفصليات والحلقيات الحديثة من الحلقات الأولية.
تتميز الحلقيات بسمات تنظيمية تقدمية: تجويف جسم ثانوي ، وجسم تشريح نقعي ، ونظام دوري مغلق (غير مغلق في اللافقاريات الأخرى) ، وحلقة حول البلعوم ، وسلسلة عصبية.
تحقق مفصليات الأرجل قدرًا أكبر من الكمال: ينشأ القلب في الدورة الدموية ؛ في الجهاز العصبي ، يزداد تركيز العقد. شكلت الهيكل الخارجي(غطاء الكيتين) ؛ مجموعة متنوعة من أعضاء الجهاز التنفسي (الخياشيم والقصبة الهوائية والرئتين) مميزة ؛ تتشكل الأطراف والأجنحة المفصلية. سمح لها هيكل أكثر كمالًا للمفصليات بتعبئة الأرض على نطاق واسع.
الرخويات لها تجويف جسم ثانوي ومعقد اعضاء داخلية(تتشكل الكلى ، قلب من ثلاث غرف ، خياشيم ، رئتان) ، معظمها لها قشرة كلسية تحمي الجسم. ومع ذلك ، في الرخويات ، لا يكون الجهاز الدوري مغلقًا ، والجهاز العصبي من النوع العقدي المتناثر.
تطور Deuterostome. تنحدر شوكيات الجلد والحبليات من أسلاف deuterostomes.
شوكيات الجلد. وهي تتميز بـ: تجويف ثانوي للجسم ، نظام الدورة الدموية يشكل فجوات ، نوع من نظام المحرك المائي (ambulacral) ، التناظر الشعاعي ، الجهاز العصبي على شكل حلقة مع أشعة نصف قطرية.
من المحتمل أن تكون الحبال الأولية قد تطورت من أسلاف تشبه الدودة. يرتبط عزل الحبليات بزيادة في التنقل. يشهد ما يلي على العلاقات التطورية بين الحبليات واللافقاريات: 1) تتكون الأنسجة والأعضاء من ثلاث طبقات جرثومية (تتشكل لأول مرة في الديدان المفلطحة) ؛ 2) التناظر الثنائي (يحدث في الديدان المفلطحة) ؛ 3) تجويف الجسم الثانوي (يحدث في الحلقات) ؛ 4) الفم الثانوي (متعلق بشوكيات الجلد) ؛ 5) الدورة الدموية المغلقة (الحلقات) ؛
من الحبليات الأولية ، نشأت الحيوانات الحديثة غير القحفية (lancelet) والفقاريات. الحبليات اليرقية هي فرع جانبي من الحبليات ، والتي تنتقل في معظمها إلى دورة حياة مستقرة ، وبالتالي لديها علامات الحبليات فقط في مرحلة اليرقات.
السيفالوثوردات هي حبليات بدائية. لديهم nephridia ، على غرار metanephridia of annelids ، وهو نظام دوري مغلق بدون قلب ، مثل annelids.
ظهر أسلاف الفقاريات منذ حوالي 500 مليون سنة. ذهب تطورهم في اتجاهين: الفك (cyclostomes) والفك (جميع الفقاريات الأخرى).
تطورت الأسماك المدرعة ذات الفك الأول من فكوك مدرعة. لديهم بالفعل أقوياء الهيكل العظمي الداخلي، الجمجمة الدماغية والحشوية ، الفكين مع الأسنان ، الزعانف المزدوجة. انقرضت هذه الأسماك. تم استبدالهم بالغضروف و الأسماك العظميةالذي كان جسمه مغطى بالحراشف.
كانت الأسماك الغضروفية شائعة في البحار ، وبحسب نوع الطعام كانت من الحيوانات المفترسة.
ظهرت الأسماك العظمية في الأصل في المياه العذبة. لقد طوروا غطاء حماية (يحمي الخياشيم من الانسداد) ومثانة سباحة (يسهل السباحة في المياه العذبة). في تجفيف المسطحات المائية (في حقب الحياة القديمة) ، ظهرت الأسماك ذات الزعانف (سمك الرئة وشحمة الزعانف) ، والتي ، بالإضافة إلى الخياشيم ، تطورت الأكياس الرئوية لتنفس الهواء ، وجعلت العضلات المتكونة على الزعانف من الممكن الاتكاء على ركيزة صلبة. بعد ذلك ، سكنت الأسماك العظمية المسطحات المائية العذبة والبحرية. في الوقت الحاضر ، تعد الأسماك العظمية واحدة من أكثر مجموعات الفقاريات ازدهارًا (20 ألف نوع).
ظهرت البرمائيات في العصر الديفوني. تنحدر البرمائيات من الأسماك القديمة ذات الزعانف. كانت البرمائيات الأولى هي Ichthyostegidae (سمكة بأربعة أقدام). لديهم ، مثل الأسماك ، زعنفة الذيل ، وغطاء الخياشيم ، والمقاييس. ولكن على عكس الأسماك ، كان لديهم زوجان من الأطراف ذات الخمسة أصابع والرئتين. كانوا يعيشون في الغالب في الماء.
من ichthyosteg جاءت البرمائيات الحقيقية - stegocephals (رأس قذيفة). كان رأس الرسغ مغطى بقشرة من عظام الجلد. عاش Stegocephalians حتى بداية العصر الطباشيري. من المحتمل أنها أدت إلى ظهور البرمائيات الحديثة الخالية من الدروع والزواحف الأولى.
كان الهبوط مصحوبًا بالرائحة: تغير التنفس الخيشومي إلى التنفس الرئوي ، والزعانف إلى أطراف بخمس أصابع ، وأصبح القلب مكونًا من ثلاث غرف ، ونشأت دائرة ثانية من الدورة الدموية ، وتغيرت أعضاء الإحساس ، وأصبح الجهاز العصبي أكثر كمالا.
ظهرت الزواحف الأولى في العصر الكربوني. كانت البقايا الأحفورية في سيموريا تحتوي على سمات البرمائيات والزواحف. من المحتمل أن تكون الزواحف الأولى ، وهي cotylosaurs ، قد نشأت من هذه الأشكال. كان لديهم أيضًا جمجمة على شكل صندوق عظمي (يشبه الدماغ) مع ثقوب للعينين والأنف والعضو الجداري. كانت ذروة الزواحف في الدهر الوسيط.
يحدث: تقرن البشرة وتكوين قشور ، أو حراشف (تحمي الجسم من الجفاف) ، مضاعفات بنية الرئتين ، القلب ، تحسين الهيكل العظمي ، تكوين الأغشية الجرثومية للمصل. و الذي يحيط بالجنين (يوفر القدرة على التكاثر على الأرض).
أدت الزواحف إلى ظهور فئتين جديدتين من الفقاريات: الثدييات والطيور.
انحدرت الطيور من الطيور الأولى من مجموعة الديناصورات السحلية التي كانت تجري عليها رجليه الخلفيتين. من الطيور القديمة الأولى ، تم العثور على بقايا الأركيوبتركس طويل الذيل. كان للأركيوبتركس علامات على الزواحف (ذيل طويل ، أسنان على الفكين ، مخالب على الأجنحة) وطيور (ريش ، أجنحة ، مظهر). ظهرت الطيور الحقيقية من الطيور الأولى في نهاية حقبة الدهر الوسيط. لديهم أيضا أسنان صغيرة.
السمات التقدمية للطيور: دم دافئ ، قلب بأربع غرف ، قوس أبهر واحد (يخلق فصلًا كاملاً للدورة الدموية الرئوية والجهازية).
ظهرت الثدييات قبل الطيور. يُفترض أنها نشأت من السحالي ذات الأسنان الحيوانية ، والتي كان لها عدد من الميزات التقدمية: توجد الأسنان في الحويصلات الهوائية ، والحنك العظمي الثانوي ، والأطراف تقع تحت الجسم ، وليس على الجانبين ، وكانت هناك تغييرات في الهيكل العظمي. كانت Cynodonts هي الأقرب إلى الثدييات بين الزواحف ذات الأسنان الثديية. يرتبط التطور الإضافي للثدييات بتغيير في نظام الأسنان. أدت هذه التغييرات إلى تكوين مجموعتين من الحيوانات. أحفاد مجموعة واحدة - المتدربون حيوانات صغيرة ، لديهم العديد من الدرنات على الأضراس ، قواطع متطورة بقوة وليس لديهم أنياب. من الممكن أن تكون هذه المجموعة قد أدت إلى ظهور أحاديات المسامير.
تطورت المجموعة الأخرى بشكل تدريجي. كانت هذه حيوانات صغيرة تأكل الحشرات والحيوانات الصغيرة. في المستقبل ، تنقسم هذه المجموعة إلى فرعين مستقلين - السفلي والجرابي والعليا والمشيمة.
نشأت الجرابيات خلال العصر الطباشيري. أقدم مجموعة من الجرابيات هي عائلة البوسوم.
ظهرت الثدييات المشيمية أيضًا في العصر الطباشيري. تعتبر أقدم مجموعة من المشيمة هي فرقة من الحشرات. من المفترض أنها أدت إلى ظهور معظم مجموعات المشيمة اللاحقة. من الممكن أن الرئيسيات انحدرت مباشرة من الحشرات.
التطور التدريجي للثدييات Stlan مع اكتساب أهم التكيفات: الدم الحار ، ارتفاع معدل التمثيل الغذائي ، تكوين المشيمة ، تغذية الصغار بالحليب ، رعاية النسل. تم تسهيل ذلك من خلال التغييرات التدريجية في الجهاز التنفسي والدورة الدموية الجهاز العصبي. للثدييات قلب مكون من أربع غرف وقوس أبهر وشرياني و الدم الوريدي، رئتي بنية سنخية ، تم تطوير القبو العصبي الثانوي - نيوباليوم جيدًا.
13 ... هيكل خلية بدائية النواة. نمو البكتيريا وتطورها وتكاثرها. أنواع البكتيريا الغذائية. التمثيل الضوئي ، التركيب الكيميائي ، الامتصاص الضوئي. قيمة الكائنات الدقيقة في الطبيعة والاقتصاد الوطني خلايا بدائية النواةتشمل البكتيريا والطحالب الخضراء المزرقة. في بدائيات النوىلا تحتوي المادة النووية على غشاء نووي ؛ يحتوي غشاء البلازما على الكربوهيدرات والأحماض الأمينية. تحتوي بدائيات النوى على كروموسوم واحد فقط ، والذي يتكون فقط من الحمض النووي. هذا الكروموسوم له شكل حلقي ولا يحتوي على بروتينات قلوية - هيستونات. إن مجموعة الكروموسومات في بدائيات النوى هي أحادية الصيغة الصبغية ، وتفتقر هذه الكائنات أحادية الخلية إلى الانقسام الفتيلي ولا تحتوي على جهاز انقسام. لا تحتوي بدائيات النوى على الانقسام الاختزالي ، وخلال العملية الجنسية يتم نقل جزء فقط من المعلومات الوراثية ، والكائن نفسه هو الأمشاج. يتم ترجمة الحمض النووي السيتوبلازمي في بدائيات النوى في البلازميدات ، والأغشية السيتوبلازمية غائبة. يحتوي جدار الخلية بدائيات النوى على ببتيدوغليكان. لا يتم ملاحظة الميتوكوندريا في بدائيات النوى ، ويتم وضع إنزيمات الجهاز التنفسي في الميزوسومات الموجودة على السطح الداخلي لغشاء البلازما ، ويتم إجراء عمليات التمثيل الضوئي باستخدام صبغة الكلوروفيل الجرثومي ، وتعمل مركبات الكبريت كعوامل اختزال. الريبوسومات بدائية النواة أصغر من الريبوسومات حقيقية النواة. لا توجد حركة في السيتوبلازم ؛ الفجوات نادرة جدًا فيه. سوط بدائيات النوى هو حجم تحت المجهر ، السوط يحتوي على واحد أو أكثر من الأنابيب الدقيقة (ليفية). بدائيات النوى حساسة للمضادات الحيوية ومقاومة للأشعة السينية و حرارة عالية، يتحمل التسخين حتى + 75 ... + 90 درجة مئوية. السمة المميزةبدائيات النوى هي أنها غير قادرة على التعايش مع أنواع أخرى من الخلايا. بكتيريا- الكائنات الحية وحيدة الخلية؛ لا يمكن رؤيتها إلا تحت المجهر. لذلك ، يطلق عليهم الميكروبات أو الكائنات الحية الدقيقة. لقد أتقنت البكتيريا مجموعة متنوعة من الموائل: فهي تعيش في التربة والغبار والماء والهواء وعلى الأغطية الخارجية للحيوانات والنباتات وداخل الجسم. يمكن العثور عليها حتى في الينابيع الساخنة ، حيث تعيش في درجات حرارة تبلغ حوالي 60 درجة مئوية أو أعلى. نشاط الكائنات الحية الدقيقة أهميةلجميع الكائنات الحية الأخرى ، لأن البكتيريا والفطريات تدمر المواد العضوية وتشارك في دورة المواد في الطبيعة. بالإضافة إلى ذلك ، تكتسب البكتيريا كل شيء قيمة أكبرفي حياة الناس ، وليس لأنها تسبب أمراضًا مختلفة ، بل لأنها يمكن استخدامها للحصول على العديد من المنتجات الضرورية.
هيكل الخلية. بدائية النواة: لا نواة مشكلة ، ميتوكوندريا ، بلاستيدات. يوجد في وسط السيتوبلازم نواة تحتوي على "كروموسوم" واحد مع جزيء دنا دائري. لا يفصله غشاء عن السيتوبلازم. داخل الخلية ، تؤدي نواتج الغشاء وظائف EPS ، جهاز جولجي ؛ يشارك الميزوسوم في التنفس ؛ هناك ريبوسومات. السيتوبلازم مغطى بغشاء خارجي وجدار خلوي كثيف. في بعض الأحيان تكون الخلية مغمورة في كبسولة غروانية شبه سائلة. وفقًا لتركيب جدار الخلية ، يمكن تقسيم البكتيريا إلى مجموعتين ، بعضهما ملطخة بالجرام ، لذلك يطلق عليها إيجابية الجرام ، بينما يصبح البعض الآخر عديم اللون عند غسل الصبغة وبالتالي يطلق عليها سالبة الجرام. النمو والتكاثر.النمو الفردي والتكاثر اللاجنسي للخلايا.نسبة السطح / الحجم للخلايا البكتيرية عالية جدًا. يساهم استحواذ سريع العناصر الغذائيةمن بيئةمن خلال الانتشار والنقل النشط. في الظروف المواتيةتنمو البكتيريا بسرعة كبيرة. النمو يعتمد في المقام الأولعلى درجة الحرارة ودرجة الحموضة في الوسط ، وتوافر العناصر الغذائية وتركيز الأيونات. تحتاج الأكياس الهوائية الملزمة أيضًا إلى الأكسجين ، وتحتاج اللاهوائية ، على العكس من ذلك ، إلى أن تكون غائبة تمامًا. بعد أن وصلت البكتيريا إلى حجم معين ، تنتقل إليها التكاثر اللاجنسي(الانشطار الثنائي) ، أي أنها تبدأ في الانقسام بتكوين خليتين ابنتيتين. يتم الانتقال إلى الانقسام من خلال نسبة حجم النواة إلى حجم السيتوبلازم. قبل انقسام الخلية ، يحدث تكاثر الحمض النووي ، حيث تحتفظ الميزوسومات بالجينوم في وضع معين ، ويمكن أن ترتبط الميزوسومات أيضًا بأقسام جديدة بين الخلايا الوليدة وتشارك بطريقة ما في تركيب مواد جدار الخلية. تنقسم البكتيريا الأسرع نموًا كل 20 دقيقة.
4 أنواع من التغذية البكتيرية وبعض خصائصها.
ثلاث مجموعات رئيسية معروفة البناء الضوئي(البكتيريا التي تحصل على الطاقة من الشمس): 1. بكتيريا الكبريت الأخضر . البكتيريا اللاهوائية التي تستخدم كبريتيد الهيدروجين (H2S) ومركبات الكبريت المختزلة الأخرى كمانحات للهيدروجين والإلكترون.
مثال: ضوء
2H2S + CO2 و rarr. و rarr. و .0.
جرثومي كلوروفيل
في هذه الحالة ، يتم ترسيب الكبريت.
2. بكتيريا الكبريت الأرجواني. تحتوي الخلايا على لون أحمر غامق ، نظرًا لوجود جرثومة أقل بكثير من الأصباغ الحمراء والبنية. هذه هي في الغالب اللاهوائية. يحدث التمثيل الضوئي فيها بنفس الطريقة كما في المجموعة السابقة.
3. البكتيريا غير الكبريتية الأرجواني . بالنسبة لهذه البكتيريا ، تعمل المركبات العضوية المختلفة كمصدر للهيدروجين ، والذي يستخدم لتقليل ثاني أكسيد الكربون (في التغذية الضوئية) أو بعض المركبات العضوية (في التغذية الضوئية).
التخليق الكيميائيالكائنات الحية (chemoautotrophs) هي بكتيريا تستخدم ثاني أكسيد الكربون كمصدر للكربون ، ولكنها تتلقى الطاقة ليس من الشمس ، ولكن بمساعدة تفاعلات كيميائية. يمكن إطلاق الطاقة ، على سبيل المثال ، أثناء أكسدة الهيدروجين وكبريتيد الهيدروجين والكبريت والحديد (II) والأمونيا والنتريت وغيرها من المركبات غير العضوية. في بكتيريا الحديد : الأكسجين
Fe2 + → و rarr. و rarr. Fe3 + + الطاقة ؛
في بكتيريا الكبريت عديمة اللون : الأكسجين أو
S → و rarr. و rarr. و SO42-+ الطاقة.
كبريتات الكبريت
في ظل الظروف اللاهوائية ، تستخدم بعض الأنواع النترات كمستقبل للهيدروجين وبالتالي تقوم بنزع النتروجين.
في البكتيريا الآزوتية: الأكسجين
2NH4 + → و rarr. و rarr. NO-2 + الطاقة
نتريت الأمونيا
جميع البكتيريا التي تم اعتبارها للتو هوائية ، لأن الأكسجين يعمل كمتقبل للإلكترونات (والهيدروجين).
تلعب البكتيريا التخليقية الكيميائية دورًا مهمًا للغاية في المحيط الحيوي ؛ يشاركون بشكل رئيسي في دورة النيتروجين وبالتالي يحافظون على خصوبة التربة.
التخفيض الضوئي - للبكتيريا المصلية الخضراء والبنفسجية ، والتي لا تستخدم H2O ، ولكن H2 S كمصدر
أهمية الطبيعة والأنشطة الاقتصادية البشرية.
بكتيريا التخمير اللبني(السابروتروف غير المتجانسة) التي تكسر الكربوهيدرات: إيجابي:تحضير منتجات حمض اللاكتيك (الجبن ، اللبن الرائب ، الزبدة ، القشدة الحامضة) ، الأعلاف ، الملفوف المخمر ، الخيار المخلل والطماطم. سلبي:تلف المنتجات.
بكتيريا التخمير الخليك(السابروتروف غير المتجانسة) التي تكسر الكربوهيدرات: إيجابي:أكسدة الكحول ل حمض الاسيتيك، والذي يستخدم لتخليل وتعليب الفواكه والخضروات. سلبي:تلف المنتجات.
البكتيريا المتعفنة(السابروتروف غيرية التغذية) التي تكسر البروتينات: إيجابي:الدور الصحي - تمعدن المخلفات العضوية. سلبي:تلف المواد الغذائية؛ لتجنب التعفن ، يتم استخدام التجفيف والتمليح والتخليل والتعقيم والبسترة والحلاوة.
بكتيريا العقيدات(المتعايشات غير المتجانسة): تخترق بكتيريا الجذور العقدية جذور النباتات البقولية (البرسيم ، الترمس ، البرسيم ، إلخ) وتدخل في التعايش معها. نتيجة لذلك ، تتشكل الأورام على الجذور - عقيدات مليئة بالبكتيريا ، والتي تصنع المركبات النيتروجينية من النيتروجين في الغلاف الجوي ، وهي متاحة لكل من النبات المضيف والنباتات الأخرى. يؤخذ هذا الإثراء الطبيعي للتربة بالأسمدة النيتروجينية في الاعتبار عند تجميع دورات المحاصيل الحقلية ، والتي تشمل بالضرورة البقوليات.
14. هيكل خلية حقيقية النواة. غشاء ثلاث طبقاتالريبوسوماتمجمع جولجيالميتوكوندرياالجسيمات المحللةالبلاستيدات الخضراءكروموبلاستسLeucoplastsمركز الخلية أو الجسيم المركزيالادراجفجواتالهيكل الخلويكاريوبلازمنوية1. 2 3 4 5
15.تطور الجنين وفتراته. التطور الجنيني المبكر. التنمية المباشرة وغير المباشرة. (51)
التكوُّن- التطور الفردي للكائن الحي منذ الولادة حتى الموت. سيستمر لمنظمات مختلفة ليست هي نفسها ويمكن أن تستمر من عدة عقول لمئات السنين (السلحفاة). يرتبط علم الوجود ارتباطًا وثيقًا بعلم التطور ولكنه ليس تكرارًا مطلقًا لعلم التطور. تحت تأثير البيئة ، يتغير ويمكن أن يحدث تقلب طفرة. فترات تطور الجنين:
1 قبل الجنين
2. جرثومية (جنينية)
3. ما بعد الجنين
في الفترة الأولى ، يتم تطوير الخلايا التناسلية وإعدادها لمزيد من التطور بعد الإخصاب (تكوين الحيوانات المنوية وتكوين البويضات) تكوين الحيوانات المنوية من 4 فترات: 1. التكاثر. 2. النمو. 3. النضج. 4. تشكيل
تكوين الحيوانات المنوية: Spermatogonia 2n "الحيوانات المنوية من الدرجة الأولى 2n" الخلايا المنوية 2 المسام 2n "الحيوانات المنوية n.
تولد البويضات: ا vogonia 2n "البويضات 1 pore2n" البويضات 2 pore2n "1 بيضة + 3 أجسام اتجاهية (منخفضة)
في الفترة الثانية ، حدث تطور الجنين في الرحم (في رحم الأم) أو في البيئة الخارجية. عمليات هار شيا السريعة للتشكيل ، والتنفيذ المدى القصيريؤدي إلى ظهور الأورغ-أماه المتعددة. لها 3 فترات: 1. الأولي- أسبوع 1 التطور الجنينييبدأ من لحظة الإخصاب وينتهي بزرع الجنين في الغشاء المخاطي للأم. 2 جرثومية.الوقت من البداية 2 ما يصل إلى 8 أسابيع من الزرع حتى اكتمال تكوين الأعضاء. 3 جنيني- نمو الجنين من 9 أسابيع حتى الولادة. تعزيز النمو ، تشكيل ، التغذية الدموية.
تبدأ الفترة الثالثة بعد إبريل من لحظة الولادة ، أي إطلاق الكائن الحي من جرثومة الأصداف. خلال هذه الفترة من التطور ، نشأت ميزة تشريحية ووظيفية محددة تسمى عمر.الفترة الزمنية:
1-10 أيام - حديثي الولادة
11 يوم - 1 سنة رضيع
1 جم - 11 ص - أطفال (بنات وفتيان)
11-15 سنة (د) و 12-16 (م) - في سن المراهقة
16-20 (د) و 17-21 (م) - شباب
21-55 (ث) و22-60 (م) - ناضجة
56 ، 61-74 - كبار السن
75-90 شيخوخة ، وأكثر من 90 من المعمرين
التطور المباشر (النوع غير اليرقي) ، المرتبط بعدد الصفار في البويضة ، أو تطور العضو. في كائن الأم ، هي سمة من سمات الأسماك والثدييات والطيور والزواحف.
التطور غير المباشر - (اليرقات) مع التحول - الشخصية. د / التناقض ، القشريات ، الحشرات ، الديدان ، الشخصية. فترة جنينية قصيرة أثناء القط. تتكون اليرقة. نفسه ولكن في الوجود. لم يتم التحول الكامل - بيضة يرقة إيماجو. تحول كامل - بيضة - يرقة - خادرة - إيماجو. التهاب الجنين المبكر:يتم إدخال الحيوانات المنوية في خلية البويضة ، ويذوب السيتوبلازم - سرب من syncarion (النوى تدمج "النواة ثنائية الصبغية" zygote 2n-div. على blastomeres-4 ، 8،16،32،64،128 ..- arr. blastula (جنين مع تجويف بداخله) - عن طريق الانغماس. - طبقتان - معدة ، إلخ. 3 جرثومة. ليستوك. الأديم الباطن ، الأديم المتوسط ، الأديم الظاهر
يمكن أن يكون التكسير متجانسًا وغير متساوٍ (قرصي)
تحدث المعدة بأربع طرق:
1 عن طريق الانغلاف (الانغلاف)
2 epiboly (قاذورات)
3 التفريغ (التقسيم الطبقي)
4 الهجرة
فتحة عصبية للأنبوب العصبي ، الوتر ، الطيات التي يحيط بالجنين والجذع
تشكل الأديم الظاهر لوحة العصب. العصب (زرع الأنبوب العصبي ، الحبال) ، السمع ، الشم ، الأغشية الخارجية ، الأديم الباطن هو بداية الأمعاء ، الكبد ، الرئتين ، غدة البنكرياس. شكل الأديم المتوسط. الجهاز العضلي (dermotome ، myotome) ، والأنسجة العضلية ، والجهاز الدوري ، وجميع العناصر الغضروفية والعظام للهيكل العظمي ، والإبراز ، والجهاز الجنسي.
16.المسارات الرئيسية لتطور نسالة. الاختلاف والتقارب والتوازي. (52)
يمكن أن يستمر التطور التطوري طرق مختلفةاعتمادًا على الكائن وعلاقته بالبيئة. هناك عدة أشكال لتكوين السلالات.
التطور Phyletic هو إعادة الترتيب التطوري التدريجي لمجموعة تصنيفية معينة ، مما يؤدي إلى ظهور مجموعة جديدة مختلفة عن المجموعة الأصلية.يمكن أن يكون أحد الأمثلة على التطور phyletic عددًا من أسلاف الخيول: eogippus - epigippus - merik-hippus - gashogippus - الحصان الحديث.
تشعب - الطريق الرئيسي للتطور ،التي كانت جزءا لا يتجزأ من أساس النظرية الداروينية. يتم تحديد التطور المتباين من خلال اختلاف الأصنف إلى عدة مجموعات تحت تأثير العوامل الأولية للتطور والتكيف مع ظروف مختلفةوجود.مع الاختلاف ، يتم تحديد التشابه من خلال أصل مشترك ، ويتم تحديد الاختلافات من خلال التكيف مع بيئة مختلفة. بداية الاختلاف داخل الأنواع أثناء تكوين الأنماط البيئية ، والأجناس البيئية ، والأنواع الفرعية الجغرافية. مع تباعد الشخصيات ، تصبح الأنواع أنواعًا ، وتخضع الأنواع لمزيد من التمايز وتشكل الأجناس ، وتصبح الأجناس عائلات ، وما إلى ذلك.
يمكن أن يتخذ الاختلاف عدة أشكال: التفرع ثنائي التفرع (تقسيم النوع إلى نوعين من الأطفال) ، الإشعاع (سقوط الجنة ، من الأنواع إلى عدة أنواع من الأطفال) ، الانقسام (يوجد كيان سلفي في وقت واحد مع الفروع ، يتشكل من معزول في وقت مختلفالسكان).يتطور الاختلاف على أساس الأعضاء المتجانسة. مثال كلاسيكييمكن أن يسمى الاختلاف اختلاف الشخصيات في عصافير أرخبيل غالاباغوس.
تكمن الأهمية البيولوجية للاختلاف في حقيقة أنه يؤدي إلى تنوع الكائنات الحية ، وإلى توزيع موحد للأنواع على منافذ بيئية ، ويخفف من حدة التوتر في الصراع من أجل الوجود.
التقارب هو العملية المعاكسة للتباعد. وهو يتألف من الاستحواذ المستقل ، في ظل نفس الظروف ، على شخصيات متشابهة في أشكال لا علاقة لها أساسًا على أساس أعضاء أو هياكل متشابهة.على سبيل المثال ، الأطراف المختبئة لدب الخلد متشابهة ؛ الحوتيات والأسماك لها نفس شكل الجسم. يحدد التقارب التشابه العام بين الطيور الخواضة "ذات القدمين الكاحلين" (الرافعات والمالك الحزين). تشمل ظواهر التقارب "المتوازيات" (المناخية) "الجغرافية" ، والتي يتم التعبير عنها في عدد أكبر من الحيوانات ذوات الدم الحار في البلدان الباردة مقارنة بأقرب ممثلي المناطق المعتدلة ، في عدد أكبر من براثن الطيور والفضلات في ثدييات القطب الشمالي ، في زيادة تصبغ الطيور الاستوائية.
هذا التقارب في مجموعات الحيوانات البعيدة يرجع إلى الظروف الخارجيةيجب على الأقارب أيضًا مراعاة القرابة. في هذا الصدد ، لوحظ التقارب داخل الفصل.
تماثل.إذا وقعت المجموعات المتباينة سابقًا في ظروف مختلفة من الوجود ، فيمكنها بشكل مستقل ، بالتوازي ، تكوين سمات تكيفية مماثلة.على سبيل المثال ، في أسماك أعماق البحار ، نشأت أجهزة التلألؤ بشكل متوازي ومستقل ؛ تطوير أسنان السيف في الماكرات المختلفة في بعض الأحيان ، تغطي التوازي أنظمة أعضاء كاملة أو جزءًا من المنظمة بأكملها. على سبيل المثال ، التطور الموازي للأسماك البرمائية التي تتنفس. من المحتمل أن يتم تحديد أوجه التشابه بين النفسين والبرمائيات من خلال العلاقة بين هذه المجموعات. أهمية خاصة هي التوازي في بنية القلب و نظام الدورة الدموية. كلما زادت الروابط بين الكائنات الحية ، كلما كان التطور الموازي في كثير من الأحيان يغطي أجزاء مهمة من المنظمة بأكملها. يتضح هذا أيضًا من خلال بيانات الحفريات ، على سبيل المثال ، تطور خرطوم التنظير. كلما اقتربت الكائنات الحية من بعضها البعض ، كلما كانت ردود أفعالها أكثر تشابهًا مع نفس التغيير في البيئة ، كلما كانت التغييرات في بنيتها أكثر تشابهًا (موازية). من أجل التنمية المتوازية ، تعتبر كل من البيئة الخارجية والعوامل الداخلية (أي الهياكل الوراثية المنشأة تاريخياً للكائن الحي) مهمة. نسبة العوامل الداخليةمع خارجي يحدد تطور الكائنات الحية. على سبيل المثال ، تفر ذوات الحوافر التي تعيش في مناطق مناخية مختلفة من الحيوانات المفترسة عن طريق الطيران. لذلك ، في ذوات الحوافر ، لوحظ تطور درجات متطرفة من أصابع القدم ثم انخفاض تدريجي في أصابع القدم الهامشية.
الأسس الجينية للتطور الموازي.مع تغيير معين في العوامل البيئية ، تتغير الكائنات ذات الصلة بالمثل. في النضال من أجل الوجود من خلال الانتقاء الطبيعي ، تتخذ هذه التغييرات شكلاً "وراثيًا". مادة الاختيار هي الانحرافات الفردية في الهيكل ، أي الطفرات. الأشكال ذات الصلة أو القريبة ، كقاعدة عامة ، تعطي نفس الطفرات (على سبيل المثال ، أنواع مختلفة من ذبابة الفاكهة). ومع ذلك ، هذا لا يعني أن الطفرات المتوازية تستند إلى نفس التغييرات في نفس الجينات.
من المعروف أن الجينات المختلفة لنفس الكائن الحي تنتج طفرات مماثلة. خاصة إذا كانت هذه التغييرات طفيفة. ونظرًا لأنه يتم اختيار مجموعات من الطفرات الصغيرة فقط أثناء التطور ، يمكن بناء التغييرات المتوازية على أساس جيني مختلف.
عمل الدورة
في علم الأحياء
"أصل حقيقيات النوى
أنجزه: سيفيروفا فيرا
الصف العاشر ، المدرسة N55
فحص بواسطة: Strukova N.A.
ريازان 1999
1. النظرية التكافلية:
1.1 التمييز الأساسي بين بدائيات النوى وحقيقيات النوى ؛
1.2 الكائنات بدائية النواة.
1.3 الخلايا حقيقية النواة.
1.4 الميتوكوندريا والبلاستيدات.
1.5 قدرة حقيقيات النوى على التمثيل الضوئي ؛
2. أسلاف الحياة.
3. الكائنات الحية المخمرة:
3.1 مصادر الطاقة للخلايا المبكرة ؛
3.2 توليد الميثان كمصدر للطاقة ؛
3.3 البكتيريا الزرقاء وبكتيريا الكلوروكسي ؛
4. التمثيل الضوئي والهواء.
5. الجمعيات وحقيقيات النوى:
5.1 نظريات التكافل المتتالي ؛
5.2 الاكتساب التكافلي للتنقل ؛
5.3 حقيقيات النوى - مواد البناء الضوئي ؛
6. نظرة على تطور الخلية:
6.1 فرضية البنوة المباشرة ؛
6.2 الفرضية: من البكتيريا الزرقاء إلى الطحالب ؛
6.3 توافق الفرضيتين: البنوة المباشرة و
التعايش الداخلي.
7. استنتاجات؛
8. الأدب.
نظرية endosyMBIOSIS
تنقسم جميع الكائنات الحية على الأرض عادةً إلى بدائيات النوى وحقيقيات النوى. الميزة الأساسيةبدائيات النوى ، على عكس حقيقيات النوى ، هي افتقارها إلى كامل نواة الخليةمغطاة بغشاء. الخلايا بدائية النواة صغيرة جدًا في الحجم ، في حدود 1 ميكرومتر. مقدار الخلايا حقيقية النواةتحتوي على نواة كاملة ، 800 - 1000 ضعف حجم الخلايا بدائية النواة. في هذا الصدد ، تحتاج الخلايا - حقيقيات النوى إلى جهاز غشاء أكبر بكثير لأداء الوظائف الحيوية. في الواقع ، في خلية حقيقية النواة ، تم تحديد الهياكل تحت الخلوية المحاطة بأغشية مستقلة عن غشاء البلازما وبعضها البعض. تسمى هذه الهياكل العضيات. كل عضية: الميتوكوندريا ، البلاستيدات الخضراء ، النواة ، الجسيمات الحالة ، جهاز جولجي ، إلخ ، لها وظائفها الخاصة في حياة الخلية.
بمساعدة المناهج الجزيئية البيولوجية ، تم الحصول على أدلة مقنعة مفادها أن جميع الكائنات الحية يجب تقسيمها إلى ثلاث ممالك: البكتيريا والعتائق وحقيقيات النوى. نشأت البكتيريا والعتائق على الأرض من سلف مشترك - ما يسمى بروجينوت - منذ حوالي 4 مليارات سنة ، ووفقًا لعلماء الأحافير ، ظهرت حقيقيات النوى على الأرض بعد 500 مليون سنة من بدائيات النوى. الفكرة الأكثر شيوعًا والمثبتة بما فيه الكفاية عن أصل الخلايا حقيقية النواة هي حاليًا فكرة التعايش الداخلي المتعدد بدائيات النوى المختلفة. وفقًا لعلماء الأحياء الدقيقة المعروفين A. أهم الطرقالاسْتِقْلاب.
تستند النظرية التكافلية لأصل الخلايا وتطورها على مفهومين جديدين في علم الأحياء.وفقًا لأول هذين المفهومين ، فإن التمييز الأساسي في الطبيعة الحية هو التمييز بين بدائيات النوى وحقيقيات النوى ، أي بين البكتيريا والكائنات الحية التي تتكون من خلايا ذات نوى حقيقية - الطلائعيات والحيوانات والفطريات والنباتات. المفهوم الثاني هو أن بعض أجزاء الخلايا حقيقية النواة نشأت من تطور التكافل ، أي تكوين روابط دائمة بين الكائنات الحية أنواع مختلفة. من المفترض أن ثلاث فئات من العضيات - الميتوكوندريا والأهداب والبلاستيدات الضوئية - نشأت من بكتيريا تعيش بحرية ، والتي ، نتيجة للتكافل ، تم تضمينها في تسلسل معين في خلايا المضيفات بدائية النواة. تعتمد هذه النظرية بشكل كبير على الأفكار الداروينية الجديدة التي طورها علماء الوراثة وعلماء البيئة وعلماء الخلايا وغيرهم من العلماء الذين ربطوا علم الوراثة المندلية بالفكرة الداروينية. الانتقاء الطبيعيكما أنه يعتمد على مجالات علمية جديدة تمامًا أو تم إحياؤها مؤخرًا: في البيولوجيا الجزيئية ، لا سيما في بنية البروتينات وتسلسل الأحماض الأمينية ، وعلم الأحياء الدقيقة ، الذي يدرس أقدم آثار الحياة على الأرض ، وحتى على فيزياء وكيمياء الغلاف الجوي ، لأن هذه العلوم مرتبطة بالغازات ذات الأصل البيولوجي. يمكن تصنيف جميع الكائنات الخلوية في خمس ممالك: مملكة بدائية النواة (مونيرا ، التي تضم البكتيريا) وممالك حقيقيات النوى الأربع (Protoctista و Animalia و Fungi و Plantae). البدائيون هم كائنات حقيقية النواة ليست حيوانات أو فطريات أو نباتات. تضم المملكة البدائية الطحالب والأوليات وقوالب الوحل والكائنات الحية حقيقية النواة الأخرى ذات الأصل غير الواضح. يتم تعريف الطلائعيات بشكل أكثر تقييدًا على أنها حقيقيات النوى أحادية الخلية. وهكذا ، فإن مملكة Protoctista لا تشمل فقط الطلائعيات - حقيقيات النوى أحادية الخلية ، ولكن أيضًا أقرب أحفادها متعددة الخلايا ، مثل الأحمر و الطحالب البنية وأيضا العديد من الكائنات الحية الدقيقة المشابهة للفطريات ، مثل الكيتريد. وفقًا لوجهة النظر التقليدية للبنوة المباشرة ، نشأت عضيات الخلية مثل الميتوكوندريا والبلاستيدات من خلال تجزئة الخلية نفسها.
العديد من أشكال بدائيات النوى
يوكاريوت واحد
الخلايا هي أنظمة مرتبطة بالغشاء ، وهي أصغر الوحدات البيولوجية القادرة على التكاثر الذاتي. حتى أصغر الخلايا تحتوي على جينات على شكل جزيئات DNA (واحد أو أكثر) وجهاز تخليق البروتين ، الذي يتكون من عدة أنواع من الحمض النووي الريبي والعديد من البروتينات. تحتوي جميع الخلايا على ريبوسومات ، يبلغ قطرها حوالي 0.02 ميكرومتر ، وتتكون من ثلاثة أنواع على الأقل من الحمض النووي الريبي وحوالي خمسين بروتينًا مختلفًا. الكائنات بدائية النواة هي أصغر الوحدات البيولوجية التي تلبي هذا التعريف للخلية. ليس لديهم نوى. من ناحية أخرى ، فإن الخلايا حقيقية النواة (التي تحتوي على نواة) هي ، بمعنى ما ، ليست وحدات على الإطلاق. هذه مجمعات من وحدات تخليق البروتين المختلفة (نيوكليوسيتوبلازم ، ميتوكوندريا ، بلاستيدات) ، يختلف نوعها وعددها اعتمادًا على الأنواع. وهكذا ، وفقًا لهذه النظرية ، نشأت الخلايا حقيقية النواة نتيجة لتعاون عناصر مستقلة في البداية ، متحدة في ترتيب معين. أطلق تايلور على فكرة الأصل المنفصل وتطور أجزاء من خلية حقيقية النواة وتوحيدها اللاحق نظرية التعايش الداخلي المتتالي. شروط درجة حرارة عاليةوأدت الحموضة إلى إنتاج صنف خاص من البروتينات - الهيستونات - في الخلية النووية لحماية الحمض النووي. كانت الميتوكوندريا قادرة على تحطيم مركبات الكربون الثلاثة إلى ثاني أكسيد الكربون والماء. بدأ الارتباط التكافلي للنيوكليوسيتوبلازم والميتوكوندريا يتغير في اتجاه تعزيز اعتمادهما المتبادل. تضمن هذا المجمع عضيات حركية - أهداب وسوط. أعقب هذا التحول تحول مورفوجيني ، والذي يتكون من تطوير عدم تناسق خلوي واضح. مع تكامل nucleocytoplasm المترابطة ، الميتوكوندريا ، undulipodia ، تم الانتهاء من إنشاء بنية متعددة الجينوم لخلية حقيقية النواة. كانت أول خلية حقيقية النواة غيرية التغذية. أكلت مركبات عضوية جاهزة. في هذه حقيقيات النوى ، نشأت عمليات الانقسام ، ثم الانقسام الاختزالي. وأخيرًا ، حدث اكتساب حقيقيات النوى للقدرة على التمثيل الضوئي في عملية أن تصبح المنظمة الرئيسية حقيقية النواة وبعد اكتمالها - الحدث الأخير في هذه الملحمة. أصبحت التكافل أكثر تكاملاً. في الوقت الحاضر ، أصبح اعتماد كل عضية على المنتجات الأيضية للعضيات الأخرى مكتملاً للغاية الأساليب الحديثةيسمح لك التحليل بتتبع المسارات الأيضية للشركاء الأصليين. تتنوع حقيقيات النوى في بنيتها ، لكنها تظل موحدة من الناحية الأيضية. تم تطوير الحيل الأيضية لحقيقيات النوى بكتيريا مختلفةحتى قبل الوقت الذي اتحدوا فيه ، تحولوا إلى مضيف وعضياته. حققت بدائيات النوى تنوعًا كبيرًا في طريقة حصولها على الطاقة وفي تعقيدات التخليق الحيوي. لقد تركوا أثرًا لوجودهم قبل فترة طويلة من تشكل الكائنات الحية الكبيرة مثل الحيوانات ذات الهيكل العظمي من الخلية حقيقية النواة.
مخطط للمحترفينو تقارب الخلايا حقيقية النواة عن طريق التكافل
مقدمات الحياة
نشأت أسلاف الكائنات الحية اليوم منذ أكثر من ثلاثة مليارات سنة من المركبات العضوية التي تشكلت على سطح الأرض. جاذبية هذا المفهوم ترجع جزئيًا إلى قابليتها للاختبار.
أظهر S. Miller و H. Urey أنه في ظل الظروف الجوية التي حدثت على الأرض الفتية ، يمكن تكوين الأحماض الأمينية والجزيئات العضوية الأخرى. أنواع مختلفةتشكلت الجزيئات العضوية بشكل تلقائي خلال المليار سنة الأولى من وجود الأرض. تم العثور على العديد من الجزيئات العضوية في الفضاء بين النجوم والنيازك. وسائل، مركبات العضويةتنشأ أثناء تفاعل عناصر الضوء ، في وجود مصادر الطاقة. براعه آلية كيميائيةيعني التكاثر الحيوي أن كل أشكال الحياة على الأرض تأتي من أسلاف مشتركة: نشأت كل الحياة الحالية من خلايا ذات أنظمة معلومات تعتمد على تكرار الحمض النووي وعلى تخليق البروتين الموجه بواسطة الرنا المرسال.
الكائنات المخمرة
من الممكن أن تكون الأحماض النووية قد تم فصلها عن البيئة منذ البداية بواسطة أغشية البروتين الدهني. بدأت المعلومات في الترميز من خلال تسلسل النيوكليوتيدات في جزيئات الحمض النووي. استخدمت أقدم الخلايا ATP والأحماض الأمينية ، والتي تعد جزءًا من البروتينات ، كمصادر مباشرة للطاقة ، وكذلك مركبات اساسيه. لذلك - كانت أقدم أشكال الحياة موجودة في شكل بكتيريا غيرية التغذية التي تلقت الغذاء والطاقة من المواد العضوية ذات الأصل اللاأحيائي. يتطلب التطور العضوي الدارويني الجديد ثلاث ظواهر: التكاثر والطفرة والضغط البيئي الانتقائي. التكاثر هو شرط غير قابل للتطبيق للتطور البيولوجي. مع التكاثر الدقيق بما فيه الكفاية ، لا مفر من الطفرات والانتقاء الطبيعي. لشرح أصل هذا التكاثر هو فهم أصل الحياة نفسها. تؤدي الطفرة ، المصدر الأساسي للتنوع في التطور ، إلى تغييرات وراثية في نظام التكاثر الذاتي. بمجرد تطوير التكاثر الموثوق به ، بدأ التطور الدارويني الجديد. الكائنات الحية التي طورت مسارات اصطناعية القفص المطلوبالمكونات ، لديها ميزة على الآخرين. أقدم ذاتية التغذية ، أي الكائنات الحية القادرة على تلبية احتياجاتها من الطاقة و المواد العضويةآه بسبب مصادر غير عضوية ، كانت هناك بكتيريا لا هوائية منتجة للميثان. تم تطوير القدرة على التمثيل الضوئي لأول مرة في البكتيريا الحساسة للأكسجين. تمتص ضوء الشمسبمساعدة الكلوروفيل البكتيري ، قاموا بتحويل ثاني أكسيد الكربون في الغلاف الجوي إلى مواد عضوية ، وقد أدى ظهور التمثيل الضوئي اللاهوائي إلى تغيير سطح الأرض وغلافها بشكل لا رجعة فيه.
تمت إزالة ثاني أكسيد الكربون من الغلاف الجوي وتحويله إلى مادة عضوية وفقًا لمعادلة التمثيل الضوئي العامة: 2H2X + CO2 = CH2O + 2X + H2O. البكتيريا الضوئية التي تستخدم كبريتيد الهيدروجين لتقليل ثاني أكسيد الكربون المترسب من عنصر الكبريت. لقد أدت إلى ظهور بكتيريا قادرة على التمثيل الضوئي مع إطلاق الأكسجين. ربما نشأ هذا النوع من التمثيل الضوئي نتيجة لتراكم الطفرات في بكتيريا الكبريت الضوئية. أدى هذا المسار إلى ظهور الطحالب الخضراء المزرقة أو البكتيريا الزرقاء. تم اكتشافه ودراسته مؤخرًا مجموعة جديدةبدائيات النوى الضوئية التي تنتج O2 - بكتيريا كلوروكسي. يمكن افتراض أن بكتيريا cyano- و chlorooxybacteria هي أسلاف البلاستيدات الموجودة في خلايا الطحالب والنباتات. نظرًا لتراكم O2 نتيجة لعملية التمثيل الضوئي الميكروبي ، أصبح الغلاف الجوي للأرض مؤكسدًا ، مما تسبب في أزمة لجميع أشكال الحياة - حتى ذلك الوقت كانت البكتيريا اللاهوائية تسكن العالم. من أجل البقاء على قيد الحياة ، كان على الميكروبات تجنب الأكسجين عن طريق الاختباء في المستنقعات ، أو كان عليها تطوير عملية التمثيل الغذائي القادرة على حمايتها من كمية الأكسجين المتزايدة.
التصوير الضوئي والهواء
منذ ما يقرب من ملياري سنة ، حدثت عمليات تطورية مختلفة واحدة تلو الأخرى لخلق التنوع الأيضي. يمكن نقل القدرة على إنتاج إنزيمات معينة داخل نفس النوع باستخدام جزيئات تشبه الفيروسات تسمى البلازميدات. قدرات بدائيات النوى على التخليق الحيوي مدهشة: من ثاني أكسيد الكربون والعديد من الأملاح والمركبات غير العضوية المختزلة التي تعمل كمصادر للطاقة ، يمكن للكائنات الدقيقة ذات التغذية الكيميائية أن تصنع في الماء غير المشرق المحتوي على الأكسجين جميع الجزيئات الكبيرة الأكثر تعقيدًا اللازمة للنمو والتكاثر. أدت هذه العمليات البيوكيميائية إلى أنظمة دوران العناصر حتى في حالة عدم وجود نباتات أو فطريات أو حيوانات.
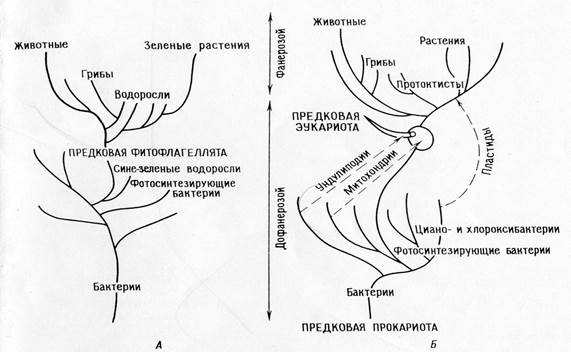
الجمعيات وحقيقيات النوى.
نظرة على تطور الخلية.
فرع مباشر.
النظرة التقليدية لأصل وتطور الخلايا حقيقية النواة هي أن جميع الكائنات الحية قد انحدرت مباشرة من مجموعة سلالة واحدة نتيجة لتراكم الطفرات تحت تأثير الانتقاء الطبيعي.
تعد الطفرات النقطية والازدواجية والحذف والتغيرات الوراثية الأخرى ، والتي من المعروف أنها لعبت دورًا في تطور الحيوانات والنباتات ، مسؤولة أيضًا عن تمايز الخلايا حقيقية النواة عن الخلايا بدائية النواة - نظرية البنوة المباشرة. أي مفهوم الأصل غير التكافلي للخلايا. ربما تكون الطحالب الخضراء من نسل أسلاف جميع النباتات. تشير العديد من سمات هذه الطحالب إلى أن لها وللنباتات سلف مشترك. تعتبر نسالة الحيوانات أكثر تعقيدًا ، ولكن ربما ينطبق مفهوم البنوة المباشرة على مساراتها التطورية أيضًا. يبدو أن الإسفنج نشأ من الطلائعيات ، بشكل مستقل عن مجموعات الحيوانات الأخرى. لذلك ، على الأقل بالنسبة للحيوانات والنباتات ، فإن الفرضية القائلة بأن الكائنات حقيقية النواة المعقدة قد تطورت من حقيقيات النوى المرتبة بشكل أكثر بساطة من خلال تراكم الطفرات الفردية تحت تأثير الانتقاء الطبيعي هي فرضية مشروعة تمامًا. تعتمد الابتكارات التطورية على الحفاظ على الجينات الملائمة المثالية للغاية. لذلك ، تطورت الكائنات الحية الجديدة ذات القدرة المتزايدة على التحكم في بيئتها عن طريق الحفاظ على تكرار الجينات شديدة التكيف وإعادة تركيبها عن طريق اكتساب خصائص جديدة من خلال طفرات نقطية بسيطة وعشوائية. لا تفسر فرضية التبويض المباشر أصل حقيقيات النوى من بدائيات النوى. على سبيل المثال ، يصعب فهم الفجوة بين البكتيريا الزرقاء غير القادرة على الانقسام ومثل هذه الأشكال ذات الانقسام الكامل مثل الطحالب الحمراء من وجهة نظر التبويض المباشر ، ولكنها تتبع بشكل طبيعي من النظرية التكافلية.
وفقًا لنظرية البنوة المباشرة ونظرية التكافل ، نشأت جميع الكائنات الحية على الأرض من أسلاف بكتيرية متشابهة كيميائيًا ، وأدت الميكروبات حقيقية النواة إلى ظهور النباتات والحيوانات. أدت بكتيريا التمثيل الضوئي إلى ظهور الطحالب ، وفي النهاية النباتات ، وفقدت بعض الطحالب بلاستيداتها وأصبحت أسلاف الفطريات والحيوانات. يتفق منظرو البنوة المباشرة على أن البلاستيدات والعضيات حقيقية النواة الأخرى ، بما في ذلك النواة ، تشكلت عن طريق التمايز داخل الخلايا نفسها ويسمي هذا أسطورة نباتية.
نسالة أنواع النباتات
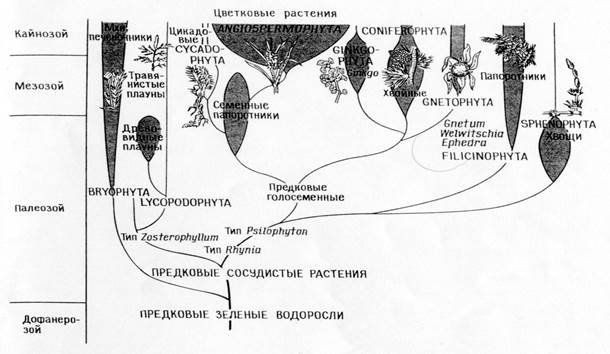
مقارنة بين السلالات المقابلة لنظرية البنوة المباشرة (أ)
ونظرية التعايش (ب)
الاختلافات بين نظرية البنوة المباشرة والنظرية التكافلية:
نظرية البنوة المباشرة.
الانقسام الرئيسي: الحيوانات - النباتات
تطورت حقيقيات النوى الضوئية (الطحالب والنباتات الخضراء) من بدائيات النوى الضوئية (الطحالب الخضراء المزرقة)
تطورت النباتات وأنظمتها الضوئية بشكل أحادي في عصر ما قبل الكمبري.
تطورت الحيوانات والفطريات من ذاتية التغذية فقدت البلاستيدات.
تتمايز الميتوكوندريا داخل الخلايا في طحالب التمثيل الضوئي.
في الطحالب اليمنى ، تمايزت أوندوليبوديا ، والجهاز الانقسام ، وجميع العضيات حقيقية النواة الأخرى داخل الخلايا.
تنحدر جميع الكائنات الحية من أسلافها - بدائيات النوى عن طريق تراكم الطفرات الفردية.
النتيجة: وفقًا للنظرية ، يجب أن يكون هناك أعشاب صحيحة ؛ من الصعب ربط هذه النظرية بسجل الحفريات وتكوين سلالة مناظرة.
نظرية التعايش الداخلي المتتالي:
الانقسام الرئيسي: بدائيات النوى - حقيقيات النوى.
تطورت حقيقيات النوى الضوئية (الطحالب والنباتات النووية) وحقيقيات النوى غير الضوئية (الحيوانات ، الفطريات ، البروتوزوا) من أسلاف مشتركة غيرية التغذية (أميبوفلاجيلات).
كان تطور التمثيل الضوئي في الأركيان أحادي الخلية في البكتيريا (بما في ذلك cyano- و chlorooxybacteria) ، نشأت الطلائعيات الضوئية فجأة عندما دخلت أسلافها غير المتجانسة في التعايش مع بدائيات النوى الضوئية (protoplastids).
تنحدر الحيوانات ومعظم الطحالب والفطريات مباشرة من الطلائعيات غيرية التغذية.
كانت الميتوكوندريا موجودة بالفعل في الطلائعيات غيرية التغذية عندما اكتسبت الأخيرة البلاستيدات نتيجة التعايش. تم الحصول على البروميتوكوندريا سابقًا بطريقة تكافلية.
تطور الانقسام المتساوي في الطلائعيات حقيقية النواة غيرية التغذية عن طريق التمايز بين النظام غير الشحمي.
تطورت بدائيات النوى من أسلاف بدائية النواة المبكرة من خلال سلسلة من الأحداث الطفرية الفردية ، كما تطورت حقيقيات النوى أيضًا من أسلاف حقيقية النواة من خلال أحداث طفرة واحدة ، وتطورت أسلاف حقيقية النواة من بدائيات النوى من خلال سلسلة من التعايش.
النتائج: تشرح النظرية المسارات البيوكيميائية وتكرارها في العضيات. إنه يسمح ببناء نسالة متسقة تشرح "الفجوة البيولوجية" بين العصر البدائي وعصر دهر الحياة. تتنبأ النظرية بأن العضيات يجب أن تحتفظ ببقايا أنظمة النسخ المتماثل.
أسطورة نباتية
من زمن جوسيو النباتات السفليةتسمى النباتات الطويلة. وفقًا للأسطورة النباتية ، فإن الأسلاف المشتركة للبكتيريا التي تعمل بالتمثيل الضوئي والطحالب الخضراء المزرقة ، من خلال التبويض المباشر ، أدت إلى ظهور طحالب حقيقية النواة ، بما في ذلك الطحالب البحرية. انتهى المطاف بمجموعات الكائنات الحية التي تأثرت نسختها بمفهوم البنوة المباشرة في مجال علم النبات - وهي البكتيريا والطحالب والفطريات والنباتات.
الكائنات الافتراضية التي يمكن أن تكون رابطًا بين البكتيريا الزرقاء والطحالب الخضراء كانت تسمى phytomonads الأسلاف (phytomonads الأجداد) أو الأعشاب الصحيحة. جادل بعض المؤلفين بأن Cyanidium و Cyanophora eukaryotes ، المتشابهة في أنظمة الصبغ الخاصة بهم مع البكتيريا الزرقاء ، قد تكون مرشحين مناسبين لدور الأعشاب الصحيحة. في النباتات الطويلة ، لم يتم العثور على أشكال وسيطة بين الحالة غير الانقسامية للبكتيريا الزرقاء والانقسام النموذجي المطور بالكامل للطحالب والنباتات الخضراء. على الرغم من الفطر و نباتات مزدهرةخالية تمامًا من undulipodia ، فهي تتميز بالانقسام والانقسام الاختزالي ، والتي تتطلب مغازل تتكون من الأنابيب الدقيقة. هناك اتفاق عام على أن جميع الكائنات متعددة الخلايا ذات الأنابيب الدقيقة تنحدر من الكائنات الحية الدقيقة حقيقية النواة ذات الأجنحة غير الشحمية ، ولكن لا أحد يعرف من أي مجموعة من الطلائعيات. كان تعزيز الأسطورة النباتية هو العصور القديمة وتعقيد عملية التمثيل الضوئي ، بالإضافة إلى حقيقة أن العديد من الكائنات الحية (البكتيريا ، الأوجلينيدات ، النباتات الطفيلية) ، سواء في الطبيعة أو في المختبر ، فقدت التمثيل الضوئي نتيجة للطفرات. نظرًا لأن عملية التمثيل الضوئي هي عملية لا هوائية تضمن في النهاية وجود جميع الكائنات الحية ، فيجب أن تكون قد تطورت في وقت مبكر جدًا من تاريخ الحياة على كوكبنا ، حتى قبل ظهور الانقسام الفتيلي ، الذي يتطلب الأكسجين. تشارك العديد من الأصباغ والدهون والإنزيمات في عملية التمثيل الضوئي ، مرتبة بطريقة منظمة في أغشية معقدة للغاية. في الوقت نفسه ، فإن تفاصيل آليات التمثيل الضوئي في الطحالب والنباتات ، من ناحية ، وفي البكتيريا الزرقاء من ناحية أخرى ، متشابهة بشكل لافت للنظر. تطلق كل هذه الكائنات غاز الأكسجين من الماء ، والذي يعمل كمصدر لذرات الهيدروجين المستخدمة لتقليل ثاني أكسيد الكربون إلى المادة العضوية للخلية. العديد من الطحالب والأوليات ليس لها انقسام ولا نظام وراثي مشابه لنظام الحيوانات والنباتات. تمتلك Ciliates نظامًا وراثيًا خاصًا به نوعان من النوى ، وغالبًا ما تكون الخلايا متعددة النوى. نوع واحد من النوى هو النوى الكبيرة ، والآخر أصغر بكثير من النوى. النوى الصغيرة غير مهمة من الناحية الفسيولوجية. ومع ذلك ، فإنهم يخزنون نسخًا من الجينات ، وأثناء العملية الجنسية ، يخضعون للانقسام الاختزالي. تعمل النوى الكبيرة من الناحية الفسيولوجية كموقع لنسخ الحمض النووي الريبي وبالتالي تتحكم في فسيولوجيا الخلية. تتمثل إحدى الصعوبات في توضيح العلاقة بين البكتيريا الزرقاء والطحالب حقيقية النواة في الافتقار إلى البيانات المتعلقة بأصل أوندوليبوديا: لم يتم العثور على روابط وسيطة بين البكتيريا الزرقاء ، التي لا تحتوي أبدًا على أوندوليبوديا ، والعديد من الطحالب التي تفعل ذلك. مشكلة أخرى هي الغياب التامالعملية الجنسية في البكتيريا الزرقاء: بين البكتيريا الزرقاء والطحالب اللاجنسية ، والتي يتميز معظمها بعملية جنسية ، ولم تكن هناك أشكال وسيطة. لقد وجد أن معظم الطحالب لديها undolipodia في مرحلة ما دورة الحياة. اتضح أن الهياكل المتنقلة في الطحالب والبروتوزوا والحيوانات والنباتات هي نفسها: كلها تتكون من أنابيب دقيقة يبلغ قطرها 0.024 ميكرون. اتضح أن المغزل الانقسامي ، حتى في الفطريات التي لا تحتوي على شحميات ، يتكون من أنابيب دقيقة بروتينية من نفس القطر كما هو الحال في undolipdia. لمدة نصف قرن ، بدا من المعقول للغاية أن الطحالب حقيقية النواة تطورت من البكتيريا الزرقاء. أدت هذه الأسطورة النباتية إلى افتراضات كانت لسنوات عديدة بمثابة حافز للبحث و نظام الإحداثيات ، فيالتي تم تقييم النتائج التي تم الحصول عليها. أصبح التأكيد على أن الروابط بين البكتيريا الزرقاء والطحالب حقيقية النواة قد انقرضت ، ولم يترك أي آثار أحفورية وآثار حية ، وأصبح أقل إقناعًا مع توفر بيانات جديدة عن الميكروبات الحية والحفريات. لم تنقرض الروابط الوسيطة الرئيسية بين بدائيات النوى الضوئية وحقيقيات النوى بدون أثر ، كما هو مطلوب في الأسطورة النباتية - لم تكن موجودة أبدًا. وفقًا لنظرية التعايش الداخلي المتتالي ، فإن البكتيريا الزرقاء هي بالفعل الأسلاف ، ولكنها فقط البلاستيدات ، وليست بقية الخلية حقيقية النواة ، وبالتأكيد ليست نواتها. ومع ذلك ، في مسألة أصل النواة ، تتوافق فرضية البنوة المباشرة ونظرية التعايش. تتضمن التعديلات التي يمكن تصورها على النسخة المتطرفة من نظرية التعايش ، على سبيل المثال ، فكرة الأصل التكافلي للبلاستيدات ، ولكن ليس الميتوكوندريا ، أو الأصل التكافلي للبلاستيدات والميتوكوندريا ، ولكن ليس من أصل شحمي. طرح أنصار البنوة المباشرة أيضًا افتراضًا حول أصل النواة من خلال التعايش.
أدلة حاسمة على أصل التكافل الداخلي للبلاستيدات الخضراء والميتوكوندريا.
تم الحصول على أهم دليل على أصل أهم عضيات الخلايا حقيقية النواة - البلاستيدات الخضراء والميتوكوندريا من بدائيات النوى في السنوات الاخيرةعند دراسة بنية RNA الريبوسوم. عالم الأحياء الجزيئي
أظهر K. Woz بالفعل في الثمانينيات أن بنية الحمض النووي الريبي من ريبوسومات البلاستيدات الخضراء النباتية متطابقة تقريبًا مع بنية الحمض النووي الريبي من بعض البكتيريا الزرقاء. في الوقت نفسه ، وجدت مجموعة من الباحثين أن الحمض النووي الريبي للريبوزومات المشتق من الميتوكوندريا لحقيقيات النوى المختلفة يشبه الحمض النووي الريبي لبعض البكتيريا ، ولا سيما بكتيريا جنس Paracoccus. مجموعة إنزيمات الجهاز التنفسي لهذه البكتيريا تشبه إلى حد بعيد
"المجموعة التنفسية" من الميتوكوندريا الحيوانية.
هذه البيانات تدعم بوضوح اكثر اعجاباأصل التكافل الداخلي للخلايا حقيقية النواة. لا تزال ظاهرة التعايش الداخلي منتشرة للغاية في الطبيعة. ومن المعروف أن على جسد البعض الأسماك البحريةغالبًا ما تستقر البكتيريا المضيئة ، والتي لا تتلقى فقط فوائد كبيرة لحياتها ، ولكنها أيضًا مفيدة جدًا للكائنات الحية المضيفة. كل ما سبق يشهد بشكل مقنع لصالح الأصل التعايش الداخلي للخلايا حقيقية النواة من خلايا بدائية النواة أصغر بكثير. ومع ذلك ، فإن السؤال الرئيسي هو تحديد طبيعة الخلية المضيفة ، لأن الخلايا حقيقية النواة لها نواة جيدة التكوين مع غلاف ، بينما لا تحتوي بدائيات النوى على مثل هذه النواة. يعتقد L.Margelis ، على سبيل المثال ، أن دور الخلايا المضيفة ربما لعبت بدائيات النوى - أسلافها البكتيريا الحديثةالمتعلقة بالميكوبلازما - بدائيات النوى المبنية بشكل بدائي للغاية ، وخالية عمليا من جدران الخلايا. في مؤخراظهرت أعمال العالم الياباني ونوقشت على نطاق واسع
تي أوشيما. استنادًا إلى دراسة شاملة لبنية الحمض النووي الريبي الريبوسومي للعديد من الكائنات الأولية وحقيقيات النوى ، بما في ذلك العديد من ممثلي مملكة العتائق التي لم تتم دراستها كثيرًا حتى الآن ، استنتج هذا المؤلف أن الخلية المضيفة كانت أحد أسلاف العتائق الحديثة. يجب أن يقال أن ممثلي هذه المملكة الفائقة ، على عكس البكتيريا ، لديهم الكثير من السمات البيولوجية البيوكيميائية والجزيئية المشتركة مع حقيقيات النوى. في أحدث أعمال T. Oshima ، تم تقديم الحجج لصالح حقيقة أن الخلية المضيفة في وقت ظهور حقيقيات النوى كانت سلف العتائق ، على غرار أحد الممثلين المعاصرين لهذه بدائيات النوى - البلازما الحرارية. ماذا سيكون الحل النهائي لمشكلة تحديد طبيعة الخلية المضيفة التي أدت إلى ظهور الخلية حقيقية النواة في عملية التعايش الداخلي المتعدد ستظهر من خلال مزيد من الدراسات.
يتم حاليًا تقديم الفرضية الأكثر شيوعًا والأكثر إثباتًا لأصل الخلايا حقيقية النواة
التعايش الداخلي للعديد من بدائيات النوى التي تنتمي إلى البكتيريا والعتائق ، والتي حدثت عدة مرات في عملية التطور خلال أول 1.5-2 مليار سنة من وجود الخلايا الحية على الأرض. تم الحصول على دليل حاسم على أصل التكافل الداخلي للبلاستيدات الخضراء والميتوكوندريا من قبل علماء الأحياء الرائدين في العالم في دراسة بنية الحمض النووي الريبي الريباسي. هذه البيانات تدعم بوضوح احتمال كبيرأصل التكافل الداخلي للخلايا حقيقية النواة من خلايا بدائية النواة أصغر بكثير (بعدة أوامر من حيث الحجم).
الأدب
1. مجلة سوروس التعليمية. علم الأحياء والكيمياء وعلوم الأرض والفيزياء والرياضيات. العدد 5 1998
2 مارجليس ل. دور التعايش في تطور الخلية. لكل. من الإنجليزية M. MIR 1983
3 أساسيات علم الأحياء الدقيقة. M. "الطب" 1992.
4 خاخينة ل. مفهوم A. Famintsyn على أهمية التعايش في تطور L. علم ، 1981 ، ص. 165 - 181.
5 ألبرتس ب. البيولوجيا الجزيئية للخلية ؛ لكل. من الانجليزية. مير 1986 v.1
يشير مصطلح بدائيات النوى (ما قبل النووي) إلى الكائنات الحية الدقيقة ذات التنظيم البدائي للهياكل النووية. تم اقتراح هذا الاسم من قبل العالم الكندي موراي في عام 1968.
في النصف الثاني من القرن العشرين. أصبح من الواضح أن جميع الكائنات الحية البنية الخلوية، يمكن تعيينها لإحدى مجموعتين - بدائيات النوى أو حقيقيات النوى. الخلايا بدائية النواة أبسط - فهي خالية من نواة محاطة بقشرة. لبعض الوقت ، ساد الاعتقاد بأن مصطلحي "بدائيات النوى" و "البكتيريا" مترادفان وأن البكتيريا تمثل فرعًا مستقلاً وموحدًا لتطور الكائنات الحية. ومع ذلك ، تراكمت الأدلة تدريجيًا على أن الاختلافات بين بعض البكتيريا يمكن أن تكون مهمة وتشير إلى أنه ليس لديهم علاقة تطورية وثيقة. خصوصاً أهمية عظيمةالمرفقة بالبيانات المتعلقة ببنية الريبوسوم ونقل الأحماض النووية الريبية (RNA). في الوقت الحاضر ، بناءً على نتائج مقارنة التسلسل الأساسي في جزيئات الحمض النووي الريبي الريبوزومي ، يتم الحكم على العلاقة بين الكائنات الحية. أظهر هذا النوع من البيانات ، بالإضافة إلى السمات الأخرى لدائيات النوى ، أنه في الواقع ، يجب تقسيم بدائيات النوى إلى مجموعتين على الأقل من أصول مختلفة ومسارات تطورية مختلفة. بالإضافة إلى ذلك ، فإن الخلايا حقيقية النواة ليست أكثر تعقيدًا من حيث التنظيم فحسب ، بل تمتلك أيضًا عناصر لم تستطع الحصول عليها من أسلاف بدائيات النوى الحديثة. هذه ، أولاً وقبل كل شيء ، الريبوسومات السيتوبلازمية ، والتي تختلف عن الريبوسومات بدائية النواة. التشابه الأساسي الكود الجينييشير تنظيم الجزيئات الكبيرة والجهاز البيوكيميائي لتخليق البروتين إلى وحدة أصل جميع الكائنات الحية. من المفترض أنه كان هناك سلف مشترك معين - "السلالة" ، لكن ما كان غير معروف. يمكن أن يؤدي هذا الأصل إلى ظهور ثلاثة فروع مستقلة للشجرة التطورية. من وجهة نظر علم اللاهوت النظامي الهرمي ، يُقترح اعتبار هذه الفروع مجالات لها مرتبة أعلى من الممالك التقليدية. هذه المجالات (الإمبراطوريات) هي Eubacteria ، Archaebacteria ، Eukaryota (eubacteria ، archaebacteria و eukaryotes). ويرد في الجدول السمات العامة والمميزة للبكتيريا البدائية والبكتيريا eubacteria وحقيقيات النوى.
السمة الرئيسية لخلية بدائية النواة هي عدم وجود نواة ، محدودة من السيتوبلازم بواسطة غشاء مزدوج. تتركز المادة الوراثية في الكروموسوم البكتيري ، وعادة ما يتم تمثيلها على شكل جزيء DNA دائري مزدوج الشريطة. تحتوي بعض اللولبيات على صبغيات خطية ، بينما تحتوي الفطريات الشعاعية على كروموسومات "الحلقة الزائفة". يعمل جزيء الحمض النووي الذي يشبه الخيط كنواة ويقع في المنطقة المركزية للخلية ، والتي تسمى "النواة النووية". في المجهر الإلكتروني ، في جزء من الخلية البكتيرية ، تبدو هذه المنطقة أفتح من بقية السيتوبلازم.
تحتوي الخلية بدائية النواة على عدد من الميزات الأساسية المتعلقة بكل من بنيتها التحتية و التنظيم الكيميائي. بادئ ذي بدء ، لا توجد شبكة غشاء في الخلية البكتيرية - الشبكة الإندوبلازمية. في الخلايا بدائية النواة ، يكون الغشاء السيتوبلازمي الموجود تحت جدار الخلية وإحاطة البروتوبلاست من الخارج هو الغشاء الوحيد في الخلية.
يحدد الغشاء السيتوبلازمي المحيط بكل خلية حجمها ويضمن الحفاظ على اختلافات كبيرة بين المحتويات الخلوية والبيئة. لكن الغشاء ليس مجرد قسم ميكانيكي. كما أنه يعمل كمرشح انتقائي للغاية يحافظ على اختلاف في تركيزات الأيونات على جانبي الغشاء ويسمح للمواد المغذية بالدخول إلى الخلية وخروج الفضلات. الغشاء السيتوبلازمي عبارة عن مجموعة من الجزيئات الدهنية والبروتينية متماسكة معًا بواسطة تفاعلات غير تساهمية. في ميكروسكوب الكترونيتبدو الأغشية مثل صفائح يبلغ سمكها حوالي 7 نانومتر مع هيكل ثلاثي الطبقات واضح. تحدد الطبقة المزدوجة من جزيئات الدهون السمات الهيكلية للغشاء ، بينما البروتينات مسؤولة عن معظم وظائف الغشاء.
الملامح الرئيسية للبكتيريا البدائية والبكتيريا eubacteria وحقيقيات النوى
| علامات | بدائيات | البكتيريا | حقيقيات النواة |
| الكائنات الحية النموذجية | الميثانوجينات ، الحرارة الشديدة ، الهالوفيل | المجموعات الرئيسية من البكتيريا | الطلائعيات والفطريات والنباتات والحيوانات |
| أبعاد نموذجية ، ميكرون | 0,5-4 | 0,5-4 | أكثر من 5 |
| الجينوم | حلقة كروموسوم | حلقة كروموسوم | نواة تحتوي على العديد من الكروموسومات المعقدة |
| الهياكل الغشائية: الشبكة الإندوبلازمية لجسيمات جولجي المعقدة | لا لا لا | لا لا لا | نعم نعم نعم |
| الميتوكوندريا | لا | لا | هنالك |
| البلاستيدات الخضراء | لا شيء (في بعض الأحيان ثايلاكويدات) | لا | لا (في الفطريات والحيوانات) ، نعم (في النباتات والطحالب) |
| جدران الخلايا | البروتين ، السودومورين | مورين ، عديدات السكاريد الدهنية | متنوع |
| الريبوسومات | 70 ثانية * | 70 ثانية | 80S (في السيتوبلازم) + 705 ريبوسومات من العضيات ، كما في البكتيريا |
| بوليميراز الحمض النووي الريبي | معقد | بسيط | معقد |
| الأسواط | بسيط | بسيطة ، تتكون من ليفية واحدة أو أكثر | هيكل معقد (2х9) +2 |
| تثبيت النيتروجين | هنالك | هنالك | لا |
| التنفس اللاهوائي | هنالك | هنالك | لا |
| التخليق الكيميائي | هنالك | هنالك | لا |
S * - وحدة ترسيب سفيدبرج تميز حجم الريبوسوم
في الغشاء السيتوبلازمي بدائيات النوى ، هناك العديد من الغزوات - الغزوات التي تؤدي وظائف مختلفة. قد تحتوي على إنزيمات وتشبه الحويصلات أو تكون ميزوسومات وتشارك في انقسام الخلايا. الغشاء السيتوبلازمي بدائيات النوى هو موقع توطين إنزيمات استقلاب الطاقة ، وبالتالي يؤدي وظيفة الميتوكوندريا ، التي لا توجد في الخلايا بدائية النواة. لذلك ، البكتيريا ذات مستوى عالالتمثيل الغذائي التنفسي ، تم تطوير الغشاء السيتوبلازمي بشكل خاص ، أي له العديد من الفتحات مما يزيد من سطحه. يشارك الغشاء السيتوبلازمي أيضًا في انقسام النواة. يتم تثبيت الدنا الدائري على الغشاء وبعد التكاثر (المضاعفة) يتباعد إلى خليتين جديدتين بسبب نمو الغشاء. تتشكل أغطية Endospore من الغشاء السيتوبلازمي. إنه أيضًا الحاجز الرئيسي لاختراق المواد إلى الخلية. لا يسمح الغشاء السيتوبلازمي للجزيئات الكبيرة بالمرور ، فهو غير قادر على كثرة الخلايا الصنوبرية وإخراج الخلايا.
الهياكل الموجودة خارج الغشاء السيتوبلازمي - جدار الخلية ، الكبسولة ، الغشاء المخاطي ، الأسواط ، الخمل ، الشعيرة ، الزغابات - هي هياكل سطحية.
يعد جدار الخلية عنصرًا هيكليًا مهمًا وإلزاميًا لمعظم الخلايا بدائية النواة (البكتيريا الحقيقية ، باستثناء البكتيريا البدائية والميكوبلازما) ، الموجودة أسفل الكبسولة أو الغشاء المخاطي ، أو في حالة عدم وجود هذا الأخير ، على اتصال بالبيئة. يعمل جدار الخلية كحاجز ميكانيكي بين البروتوبلاست و بيئة خارجيةويعطي الخلية شكلاً معينًا متأصلًا فيها.
من حيث التركيب والتركيب الكيميائي ، يختلف جدار خلية بدائيات النوى بشكل حاد عن تلك الموجودة في الكائنات حقيقية النواة. يتكون جدار الخلية بدائيات النوى من عدة طبقات ، تتكون منها الطبقة الرئيسية - الإطار - المسؤولة عن القوة ، من مادة خاصة - مورين ببتيدوغليكان ، والتي يتم تصنيعها فقط بواسطة خلية بدائية النواة ولا توجد في الكائنات الحية الأخرى. في مجموعة بدائيات النوى الحقيقية ، هناك نوعان رئيسيان من بنية جدار الخلية. يتوافق هذان النوعان مع البكتيريا موجبة الجرام وسالبة الجرام. تأتي أسماء هذه المجموعات من القدرة أو عدم القدرة بكتيريا مختلفةوصمة عار حسب الجرام. في عام 1884 ، اقترح الطبيب الدنماركي ك.غرام ، الذي كان متورطًا في تلطيخ الأنسجة ، طريقة تلطيخ ، والتي تم استخدامها لاحقًا للبكتيريا. إذا عولجت الخلايا الثابتة من البكتيريا الخبيثة أولاً بمحلول من البنفسجي الكريستالي ثم باليود ، فسيتم تكوين مركب ملون. أثناء العلاج اللاحق بالكحول ، اعتمادًا على بنية جدار الخلية ، يختلف مصير المركب: في ما يسمى بالأنواع إيجابية الجرام ، تحتفظ الخلية بهذا المركب وتبقى الأخيرة ملونة ، في سالبة الجرام. الأنواع ، يتم غسل المركب الملون من الخلية ، ويتغير لونها. وجد أن المركب الملون يتشكل على البروتوبلاست ، لكن احتباسه بواسطة الخلية أو الرشح منه أثناء العلاج اللاحق بالكحول يتم تحديده من خلال السمات الهيكلية لجدار الخلية. وجدت طريقة صبغ غرام تطبيق واسعفي الممارسة الميكروبيولوجية لتمييز البكتيريا.
تختلف جدران الخلايا للبكتيريا eubacteria إيجابية الجرام وسالبة الجرام بشكل حاد في كل من التركيب الكيميائي والبنية التحتية الدقيقة.
الجزء الأكبر من جدار الخلية للبكتيريا موجبة الجرام هو بوليمر مغاير محدد - ببتيدوغليكان مورين. تم بناء العمود الفقري متعدد السكاريد للجزيء من بقايا متناوبة من N-acetylglucosamine و N-acetylmuramic acid ، المترابطة بواسطة روابط جليكوسيدية. بالإضافة إلى الببتيدوغليكان ، تشتمل جدران خلايا البكتيريا eubacteria إيجابية الجرام على أحماض teichoic و polysaccharides. أحماض Teichoic عبارة عن بوليمرات تعتمد على كحول متعدد الهيدروكسيل(ريبيتول وجليسيرول) ، مترابطان بواسطة روابط فوسفوديستر.
تحتوي البكتيريا eubacteria سالبة الجرام على هيكل جدار خلوي أكثر تعقيدًا من تلك التي تحتوي على موجبة الجرام. يحتوي على الكثير أكثرالجزيئات الكبيرة من مختلف النوع الكيميائي. تشكل الببتيدوغليكان فقط الطبقة الداخلية لجدار الخلية ، وتلتصق بشكل فضفاض بالغشاء السيتوبلازمي. خارج الببتيدوغليكان توجد طبقة إضافية من جدار الخلية - الغشاء الخارجي، والتي تتكون من الدهون الفوسفورية (نموذجية للأغشية الأولية) والبروتينات والبروتينات الدهنية وعديدات السكاريد الدهنية.
تم العثور على الاختلافات الموجودة تجريبيا بين مجموعتي البكتيريا فيما يتعلق ببقع غرام لتكون مرتبطة بالعديد من السمات الهامة الأخرى. على سبيل المثال ، تتكاثر البكتيريا موجبة الجرام عن طريق الانشطار الثنائي بسبب تكوين الحاجز المستعرض في الخلية ، سالب الجرام - عن طريق تكوين انقباض. الأولى حساسة للبنسلين ، والأخيرة ليست كذلك. لا تشكل البكتيريا سالبة الجرام أبواغًا داخلية ، في حين أن بعض البكتريا موجبة الجرام تفعل ذلك.
يؤدي جدار الخلية بدائيات النوى مجموعة متنوعة من الوظائف: فهو يحمي الخلية ميكانيكيًا من التأثيرات البيئية ، ويحافظ على شكلها الخارجي ، ويمكّن الخلية من التواجد في حلول ناقصة التوتر. بادئ ذي بدء ، هذه هي "ميزة" الببتيدوغليكان. ترتبط النفاذية ونقل المواد إلى الخلية بجدار الخلية.
تحت الغشاء السيتوبلازمي في البكتيريا يوجد السيتوبلازم. هذا نظام غرواني يتكون من الماء والبروتينات والدهون والكربوهيدرات والمركبات المعدنية والمواد الأخرى ، وتختلف نسبتها حسب نوع البكتيريا وعمرها. لا يخضع السيتوبلازم لخلية بدائية النواة للحركة الدورية ، كما هو الحال في النباتات ، أو الظواهر الانقباضية ، كما هو الحال في الأميبات. الخلايا البكتيريةتنفيذ الحركة بسبب آليات أخرى: إما بمساعدة سوط مرتبة ببساطة ، والتي تختلف عن الأسواط المعقدة لحقيقيات النوى ، فإنها تسبح بنشاط في وسط سائل أو تنزلق ، وتفرز المخاط وتتحرك على طول سطح ركيزة كثيفة.
تتحرك الخلايا البكتيرية المتحركة بنشاط في الاتجاه الذي تحدده عوامل معينة. تسمى هذه الحركات الموجهة للبكتيريا "سيارات الأجرة". هناك انجذاب كيميائي ، محور ضوئي ، انجذاب مغناطيسي ، انجذاب حراري ، انجذاب لزج (استجابة لتغير في لزوجة المحلول).
يتم تمثيل الخلايا البكتيرية المستريحة في البكتيريا موجبة الجرام عن طريق الأبواغ ، في البكتيريا سالبة الجرام - بواسطة الأكياس. تتمتع الجراثيم البكتيرية بمقاومة فريدة للتأثيرات الضارة. اللافت للنظر بشكل خاص هو مقاومة الجراثيم البكتيرية للحرارة: فبعض الأبواغ تتحمل الغليان لمدة ساعة أو أكثر ولا تفقد قابليتها للحياة. مقاومة شديدة للجراثيم وللتجفيف. يرتبط استقرار الجراثيم بوجود طبقة خاصة في جدارها الخلوي - القشرة.
أثناء تكوين البوغ ، تكتسب الخلايا البكتيرية أحيانًا شكلًا غير عادي من المغزل أو الليمون أو عصا الطبل. عادة ما يكون البوغ الداخلي واحدًا لكل خلية ، ولكن هناك بكتيريا بها أكثر من أربعة جراثيم. بعض البكتيريا اللاهوائيةعلى الجراثيم ، تتشكل أغطية خلوية ، تتكون من فجوات غازية ، أو أنواع مختلفة من النواتج
الجراثيم البكتيرية ليست هي الأشكال الوحيدة من السكون في بدائيات النوى. تتحول البكتيريا التي تقرأ الجرام ، غير القادرة على تكوين الأبواغ الداخلية ، في بعض الحالات إلى أكياس - خلايا خاصةمع أغطية خارجية سميكة (على سبيل المثال. , Azotobacter chroococcum)
في الخارج ، غالبًا ما تكون جدران الخلايا بدائيات النوى محاطة بمادة مخاطية. تسمى هذه التشكيلات ، اعتمادًا على السمات الهيكلية ، الكبسولات أو الطبقات المخاطية أو الأغماد أو الأغماد (في البكتيريا الخيطية). كل منهم هو نتيجة التخليق الحيوي للبوليمرات العضوية بدائيات النوى وترسبها حول الخلايا.
قد تحتوي الكبسولات والتكوينات المخاطية والأغلفة على مكونات مماثلة لجدار الخلية ، لكنها التركيب الكيميائيلم تكن متطابقة. كقاعدة عامة ، يكون التركيب الكيميائي للكبسولات التي تشكلها البكتيريا خاصًا بالجنس والأنواع. المكونات الكيميائية الرئيسية لمعظم كبسولات بدائية النواة هي السكريات المتعددة. الاستثناء هو كبسولات بعض أنواع العصيات ، المبنية من بولي ببتيد ، وهو بوليمر من حمض الجلوتاميك. حالات - أكثر الهياكل المعقدة، تحتوي على تركيبة كيميائية أكثر تعقيدًا وتحتوي على السكريات والبروتينات والدهون.
الكبسولات لها سمك مختلف ويمكن التعرف عليها بسهولة تحتها
تحت المجهر مع تباين سلبي من المستحضر بالحبر الأسود.
تحمي الكبسولات والأغلفة الخلية من التلف والجفاف ، وتخلق حاجزًا تناضحيًا إضافيًا ، وتعمل كعقبة أمام تغلغل العاثيات ، وتخلق بيئة خاصة حول الخلايا توفر نوعًا معينًا نظام الماء. الكبسولات لها وظائف محددة. في بكتيريا العقيدات ، على سبيل المثال ، تحدد الارتباط الانتقائي بشعر جذور النباتات من مختلف الأنواع. في بعض الأحيان يمكن استخدام الكبسولات كمغذيات احتياطية. بمساعدة المخاط ، يتم الاتصال بين الخلايا في المستعمرة ، وربط الخلايا بأسطح مختلفة
تتفاعل الكبسولات المتعددة السكاريد مع كاتيونات محاليل التربة ، ويمكنها تدمير معادن التربة ، والمشاركة في عمليات التجميع وتشكيل بنية تربة مقاومة للماء. ربما يتم إدراج المخاط الميكروبي خارج الخلية في عملية تكوين المواد الدبالية.
شكل خلية بدائيات النوى متنوع للغاية. بدائيات النوى المعروفة كروية ، على شكل قضيب ، ملتوية حلزونيًا ، على شكل فاصلة ، على شكل حلقة - مغلقة أو مفتوحة ، مع نواتج - بسيطة. قد تكون الخلايا مربعة أو على شكل نجمة. بدائيات النوى متعددة الخلايا هي خيوط على شكل خيوط أو دودة.
تنتمي جميع بدائيات النوى إلى مجموعة الكائنات المجهرية ، ويقاس حجم خلاياها بالميكرونات (ميكرومتر) (1 ميكرومتر = 10 3 مم) ، ويتم قياس تفاصيلها بالنانومتر (1 نانومتر = 10 مم). ما يقرب من 40 ٪ من البكتيريا في التربة تفوق قدرة المجهر الضوئي ، فهي مرئية فقط في المجهر الإلكتروني. في مثل هذه الأحجام الصغيرة ، تكون نسبة السطح إلى الحجم عالية جدًا ، مما يؤدي إلى ارتفاع معدل التمثيل الغذائي المحتمل.
تنقسم جميع بدائيات النوى إلى أربع مجموعات وفقًا لبنية التكامل الخارجي للخلية: eubacteria سالبة الجرام (Gracilicutes) ، eubacteria موجبة الجرام (Firmicutes) ، الميكوبلازما (Mollicutes) ، الخالية من جدران الخلايا ، والبكتيريا القديمة ، أو العتائق (Archaebacteria). تم توحيد المجموعات الثلاث الأولى على أنها eubacteria (Eubacteria) ، والبكتيريا الأثرية هي مجموعة منفصلة من الكائنات الحية (Archaebacteria) تختلف عن غيرها في بنية جزيئات RNA الريبوزومية ونقلها ، و RNA polymerases ، والدهون ، وهيكل جدار الخلية.
تحتوي البكتيريا سالبة الجرام على جدران خلوية ، حيث يوجد أيضًا ، بالإضافة إلى طبقة الإطار الرئيسية للمورين ، غشاء خارجي وطبقة من البروتين و / أو بروتين سكري متداخلة من الأعلى. هناك مساحة محيطية بين الغشاء السيتوبلازمي وطبقة مورين. هذه المجموعة من البكتيريا هي الأكثر انتشارًا وتنوعًا من حيث عدد الأنواع و وظائف فسيولوجية. تجمع مجموعة بدائيات النوى بين البكتيريا الضوئية والتغذية الكيميائية. ومن بين هذه الكائنات السابروتروفية الشائعة ، والنباتات المتساقطة ، وقلة التغذية ، والطفيليات داخل الخلايا للبكتيريا الأخرى ، وتشكل مسببات الأمراض للإنسان والحيوان والنباتات ذات الاحتياجات الغذائية المعقدة. حسب علم التشكل ، فهذه بكتيريا متنوعة من pseudomonads بسيطة الشكل على شكل قضيب إلى البكتيريا المخاطية مع دورة تطوير معقدة.
الميكوبلازما هي بكتيريا ليس لها جدار خلوي ، يقوم الغشاء السيتوبلازمي بوظيفتها.
توحد البكتيريا البدائية بدائيات النوى مع بنية مختلفة من التكامل الخارجي ، والتي تفتقر إلى مورين ، وهي سمة من سمات البكتيريا الحقيقية. بعض ممثلي البكتيريا البدائية لديهم pseudomurein في جدار الخلية ، بينما يمتلك البعض الآخر جدارًا خلويًا يتكون من طبقة من البروتينات الكروية أو البروتينات السكرية.
يعد تقسيم البكتيريا إلى مجموعات بناءً على الخصائص المظهرية (مورفولوجيا ، وعلم وظائف الأعضاء) مناسبًا للأغراض العملية للتعرف السريع على الكائنات الحية. يتم تجميع الكائنات الحية في مجموعات مصطنعة ، أو "أقسام" ، والتي ليس لها حالة تصنيفية وغالبًا ما يتم تحديدها من قبل معظمها نوع مميزأو العنوان. وفقًا لهذا المبدأ ، تم بناء محدد بيرجي للبكتيريا.
من خلال دراسة تنوع الكائنات الحية التي سكنت الأرض وسكنتها ، يسعى علماء التصنيف إلى إنشاء نظام نسبي يعكس علاقة الأنساب والأصل المشترك للكائنات الحية المتحدة في مجموعة أو أخرى - تصنيف. يتم تقديم هذا الهدف من خلال النظاميات الجزيئية ، بناءً على تشابه بنية البوليمرات الحيوية - الحمض النووي ، والحمض النووي الريبي ، والبروتينات. من أجل القدرة على تحديد تشابه الجينات ، غالبًا ما يطلق عليها علم النظم الجينية. أحد أهم إنجازات علم اللاهوت الجيني هو اكتشاف مجموعة جديدة من الكائنات الحية الدقيقة المعزولة في مملكة منفصلة - البكتيريا القديمة. يتم تحديد درجة تشابه الجينوم البكتيري بمقارنة تسلسل النوكليوتيدات في جزيء الرنا الريباسي 16S. استخدم استخدام هذه الطريقة في تصنيف البكتيريا كأساس لبناء شجرة النشوء والتطور للبكتيريا بواسطة K. Wese.
من الصعب للغاية مقارنة النظاميات النشوء والتطور. تحتوي نفس الفروع التطورية على كائنات متنوعة للغاية من حيث علم وظائف الأعضاء. عند الانتقال من الفروع الكبيرة للبكتيريا موجبة الجرام وسالبة الجرام إلى المستويات الهرمية السفلية ، تصبح حالات عدم التطابق هذه أكثر فأكثر.
عمل الدورة
في علم الأحياء
"أصل حقيقيات النوى
أنجزه: سيفيروفا فيرا
الصف العاشر ، المدرسة N55
فحص بواسطة: Strukova N.A.
ريازان 1999
1. النظرية التكافلية:
1.1 التمييز الأساسي بين بدائيات النوى وحقيقيات النوى ؛
كائنات بدائية النواة
الخلايا حقيقية النواة؛
الميتوكوندريا والبلاستيدات.
قدرة حقيقيات النوى على التمثيل الضوئي ؛
2. أسلاف الحياة.
3. الكائنات الحية المخمرة:
مصادر الطاقة للخلايا المبكرة ؛
توليد الميثان كمصدر للطاقة ؛
البكتيريا الزرقاء والبكتيريا الكلوروكسي.
البناء الضوئي والهواء.
الجمعيات وحقيقيات النوى:
نظريات التعايش المتتالية.
اكتساب تكافلي للتنقل ؛
حقيقيات النوى هي ضوئي.
نظرة على تطور الخلية:
6.1 فرضية البنوة المباشرة ؛
الفرضية: من البكتيريا الزرقاء إلى الطحالب.
توافق الفرضيتين: البنوة المباشرة و
التعايش الداخلي.
الأدب.
نظرية EndosyMBIOSIS تنقسم جميع الكائنات الحية على الأرض عادة إلى بدائيات النوى وحقيقيات النوى. السمة الرئيسية لدائيات النوى ، على عكس حقيقيات النوى ، هي عدم وجود نواة خلية كاملة مغطاة بغشاء. الخلايا بدائية النواة صغيرة جدًا في الحجم ، في حدود 1 ميكرومتر. حجم الخلايا حقيقية النواة التي تحتوي على نواة كاملة هو 800-1000 مرة أكبر من حجم الخلايا بدائية النواة. في هذا الصدد ، تحتاج الخلايا - حقيقيات النوى إلى جهاز غشاء أكبر بكثير لأداء الوظائف الحيوية. في الواقع ، في خلية حقيقية النواة ، تم تحديد الهياكل تحت الخلوية المحاطة بأغشية مستقلة عن غشاء البلازما وبعضها البعض. تسمى هذه الهياكل العضيات. كل عضية: الميتوكوندريا ، البلاستيدات الخضراء ، النواة ، الجسيمات الحالة ، جهاز جولجي ، إلخ ، لها وظائفها الخاصة في حياة الخلية. بمساعدة المناهج الجزيئية البيولوجية ، تم الحصول على أدلة مقنعة مفادها أن جميع الكائنات الحية يجب تقسيمها إلى ثلاث ممالك: البكتيريا والعتائق وحقيقيات النوى. نشأت البكتيريا والعتائق على الأرض من سلف مشترك - ما يسمى بروجينوت - منذ حوالي 4 مليارات سنة ، ووفقًا لعلماء الأحافير ، ظهرت حقيقيات النوى على الأرض بعد 500 مليون سنة من بدائيات النوى. الفكرة الأكثر شيوعًا والمثبتة بما فيه الكفاية عن أصل الخلايا حقيقية النواة هي حاليًا فكرة التعايش الداخلي المتعدد بدائيات النوى المختلفة. وفقًا لعلماء الأحياء المجهرية المعروفين A. Kluyver و K. Van Niel ، كان في بدائيات النوى القديمة أن تم اختيار الخلايا ذات التفاعلات الكيميائية الحيوية الأكثر اقتصادا والتي غالبًا ما تكون متعددة الوظائف ، والتي أصبحت أساس أهم مسارات التمثيل الغذائي.
تستند النظرية التكافلية لأصل الخلايا وتطورها على مفهومين جديدين في علم الأحياء.وفقًا لأول هذين المفهومين ، فإن التمييز الأساسي في الطبيعة الحية هو التمييز بين بدائيات النوى وحقيقيات النوى ، أي بين البكتيريا والكائنات الحية التي تتكون من خلايا ذات نوى حقيقية - الطلائعيات والحيوانات والفطريات والنباتات. المفهوم الثاني هو أن بعض أجزاء الخلايا حقيقية النواة نشأت من تطور التكافل ، أي تكوين روابط دائمة بين الكائنات الحية من الأنواع المختلفة. من المفترض أن ثلاث فئات من العضيات - الميتوكوندريا والأهداب والبلاستيدات الضوئية - نشأت من بكتيريا تعيش بحرية ، والتي ، نتيجة للتكافل ، تم تضمينها في تسلسل معين في خلايا المضيفات بدائية النواة. تعتمد هذه النظرية بشكل كبير على الأفكار الداروينية الجديدة التي طورها علماء الوراثة وعلماء البيئة وعلماء الخلايا وغيرهم من العلماء الذين ربطوا علم الوراثة المندلية بالفكرة الداروينية عن الانتقاء الطبيعي. كما تعتمد أيضًا على اتجاهات علمية جديدة تمامًا أو تم إحياؤها مؤخرًا: في البيولوجيا الجزيئية ، خاصة فيما يتعلق بتركيب البروتين وبيانات تسلسل الأحماض الأمينية ، وعلم الأحياء الدقيقة ، الذي يدرس الآثار المبكرة للحياة على الأرض ، وحتى في فيزياء وكيمياء الغلاف الجوي ، لأن هذه العلوم مرتبطة بالغازات ذات الأصل البيولوجي. يمكن تصنيف جميع الكائنات الخلوية في خمس ممالك: مملكة بدائية النواة (مونيرا ، التي تضم البكتيريا) وممالك حقيقيات النوى الأربع (Protoctista و Animalia و Fungi و Plantae). البدائيون هم كائنات حقيقية النواة ليست حيوانات أو فطريات أو نباتات. تضم المملكة البدائية الطحالب والأوليات وقوالب الوحل والكائنات الحية حقيقية النواة الأخرى ذات الأصل غير الواضح. يتم تعريف الطلائعيات بشكل أكثر تقييدًا على أنها حقيقيات النوى أحادية الخلية. وهكذا ، فإن مملكة Protoctista لا تشمل فقط الطلائعيات - حقيقيات النوى أحادية الخلية ، ولكن أيضًا أقرب أحفادها متعددة الخلايا ، مثل الطحالب الحمراء والبنية ، بالإضافة إلى العديد من الكائنات الحية الدقيقة المشابهة للفطريات ، مثل chytrids. وفقًا لوجهة النظر التقليدية للبنوة المباشرة ، نشأت عضيات الخلية مثل الميتوكوندريا والبلاستيدات من خلال تجزئة الخلية نفسها.
العديد من بدائيات النوى من قطعة واحدة من أحجار الأوكاريوت
الخلايا هي أنظمة مرتبطة بالغشاء ، وهي أصغر الوحدات البيولوجية القادرة على التكاثر الذاتي. حتى أصغر الخلايا تحتوي على جينات على شكل جزيئات DNA (واحد أو أكثر) وجهاز تخليق البروتين ، الذي يتكون من عدة أنواع من الحمض النووي الريبي والعديد من البروتينات. تحتوي جميع الخلايا على ريبوسومات ، يبلغ قطرها حوالي 0.02 ميكرومتر ، وتتكون من ثلاثة أنواع على الأقل من الحمض النووي الريبي وحوالي خمسين بروتينًا مختلفًا. الكائنات بدائية النواة هي أصغر الوحدات البيولوجية التي تلبي هذا التعريف للخلية. ليس لديهم نوى. من ناحية أخرى ، فإن الخلايا حقيقية النواة (التي تحتوي على نواة) هي ، بمعنى ما ، ليست وحدات على الإطلاق. هذه مجمعات من وحدات تخليق البروتين المختلفة (نيوكليوسيتوبلازم ، ميتوكوندريا ، بلاستيدات) ، يختلف نوعها وعددها اعتمادًا على الأنواع. وهكذا ، وفقًا لهذه النظرية ، نشأت الخلايا حقيقية النواة نتيجة لتعاون عناصر مستقلة في البداية ، متحدة في ترتيب معين. أطلق تايلور على فكرة الأصل المنفصل وتطور أجزاء من خلية حقيقية النواة وتوحيدها اللاحق نظرية التعايش الداخلي المتتالي. أدت ظروف درجات الحرارة المرتفعة والحموضة إلى إنتاج صنف خاص من البروتينات - الهيستونات - في الخلية النووية لحماية الحمض النووي. كانت الميتوكوندريا قادرة على تحطيم مركبات الكربون الثلاثة إلى ثاني أكسيد الكربون والماء. بدأ الارتباط التكافلي للنيوكليوسيتوبلازم والميتوكوندريا يتغير في اتجاه تعزيز اعتمادهما المتبادل. تضمن هذا المجمع عضيات حركية - أهداب وسوط. أعقب هذا التحول تحول مورفوجيني ، والذي يتكون من تطوير عدم تناسق خلوي واضح. مع تكامل nucleocytoplasm المترابطة ، الميتوكوندريا ، undulipodia ، تم الانتهاء من إنشاء بنية متعددة الجينوم لخلية حقيقية النواة. كانت أول خلية حقيقية النواة غيرية التغذية. أكلت مركبات عضوية جاهزة. في هذه حقيقيات النوى ، نشأت عمليات الانقسام ، ثم الانقسام الاختزالي. وأخيرًا ، حدث اكتساب حقيقيات النوى للقدرة على التمثيل الضوئي في عملية أن تصبح المنظمة الرئيسية حقيقية النواة وبعد اكتمالها - الحدث الأخير في هذه الملحمة. أصبحت التكافل أكثر تكاملاً. في الوقت الحاضر ، أصبح اعتماد كل عضية على المنتجات الأيضية للعضيات الأخرى مكتملاً لدرجة أن طرق التحليل الحديثة فقط تجعل من الممكن تتبع المسارات الأيضية للشركاء الأصليين. تتنوع حقيقيات النوى في بنيتها ، لكنها تظل موحدة من الناحية الأيضية. تم تطوير الحيل الأيضية لحقيقيات النوى بواسطة بكتيريا مختلفة حتى قبل الوقت الذي اجتمعت فيه لتصبح مضيفًا وعضياتها. حققت بدائيات النوى تنوعًا كبيرًا في طريقة حصولها على الطاقة وفي تعقيدات التخليق الحيوي. لقد تركوا أثرًا لوجودهم قبل فترة طويلة من تشكل الكائنات الحية الكبيرة مثل الحيوانات ذات الهيكل العظمي من الخلية حقيقية النواة.
مخطط أصل الخلايا حقيقية النواة عن طريق التكافل
مقدمات الحياة
نشأت أسلاف الكائنات الحية اليوم منذ أكثر من ثلاثة مليارات سنة من المركبات العضوية التي تشكلت على سطح الأرض. جاذبية هذا المفهوم ترجع جزئيًا إلى قابليتها للاختبار.
أظهر S. Miller و H. Urey أنه في ظل الظروف الجوية التي حدثت على الأرض الفتية ، يمكن تكوين الأحماض الأمينية والجزيئات العضوية الأخرى. تشكلت أنواع مختلفة من الجزيئات العضوية تلقائيًا خلال المليار سنة الأولى من وجود الأرض. تم العثور على العديد من الجزيئات العضوية في الفضاء بين النجوم والنيازك. هذا يعني أن المركبات العضوية تنشأ أثناء تفاعل العناصر الخفيفة ، في وجود مصادر الطاقة. تعني عالمية الآلية الكيميائية للتكاثر الحيوي أن كل أشكال الحياة على الأرض تأتي من أسلاف مشتركة: نشأت كل الحياة الحالية من خلايا ذات أنظمة معلومات تعتمد على تكرار الحمض النووي وعلى تخليق البروتين الموجه بواسطة الرنا المرسال.
الكائنات المخمرة
من الممكن أن تكون الأحماض النووية قد تم فصلها عن البيئة منذ البداية بواسطة أغشية البروتين الدهني. بدأت المعلومات في الترميز من خلال تسلسل النيوكليوتيدات في جزيئات الحمض النووي. استخدمت أقدم الخلايا ATP والأحماض الأمينية ، وهي جزء من البروتينات ، كمصادر مباشرة للطاقة ، وكمكونات هيكلية. لذلك - كانت أقدم أشكال الحياة موجودة في شكل بكتيريا غيرية التغذية التي تلقت الغذاء والطاقة من المواد العضوية ذات الأصل اللاأحيائي. يتطلب التطور العضوي الدارويني الجديد ثلاث ظواهر: التكاثر والطفرة والضغط البيئي الانتقائي. التكاثر هو شرط غير قابل للتطبيق للتطور البيولوجي. مع التكاثر الدقيق بما فيه الكفاية ، لا مفر من الطفرات والانتقاء الطبيعي. لشرح أصل هذا التكاثر هو فهم أصل الحياة نفسها. تؤدي الطفرة ، المصدر الأساسي للتنوع في التطور ، إلى تغييرات وراثية في نظام التكاثر الذاتي. بمجرد تطوير التكاثر الموثوق به ، بدأ التطور الدارويني الجديد. الكائنات التي طورت مسارات لتخليق المكونات التي تحتاجها الخلية كانت لها ميزة على غيرها. كانت البكتيريا اللاهوائية المنتجة للميثان هي أقدم الكائنات ذاتية التغذية ، أي الكائنات الحية القادرة على تلبية احتياجاتها من الطاقة والمواد العضوية من مصادر غير عضوية. تم تطوير القدرة على التمثيل الضوئي لأول مرة في البكتيريا الحساسة للأكسجين. بامتصاص أشعة الشمس بمساعدة الكلوروفيل البكتيري ، قاموا بتحويل ثاني أكسيد الكربون الموجود في الغلاف الجوي إلى مواد عضوية ، وقد أدى ظهور التمثيل الضوئي اللاهوائي إلى تغيير سطح الأرض وغلافها بشكل لا رجعة فيه.
تمت إزالة ثاني أكسيد الكربون من الغلاف الجوي وتحويله إلى مادة عضوية وفقًا لمعادلة التمثيل الضوئي العامة: 2H2X + CO2 = CH2O + 2X + H2O. البكتيريا الضوئية التي تستخدم كبريتيد الهيدروجين لتقليل ثاني أكسيد الكربون المترسب من عنصر الكبريت. لقد أدت إلى ظهور بكتيريا قادرة على التمثيل الضوئي مع إطلاق الأكسجين. ربما نشأ هذا النوع من التمثيل الضوئي نتيجة لتراكم الطفرات في بكتيريا الكبريت الضوئية. أدى هذا المسار إلى ظهور الطحالب الخضراء المزرقة أو البكتيريا الزرقاء. في الآونة الأخيرة ، تم اكتشاف ودراسة مجموعة جديدة من بدائيات النوى المنتجة للأكسجين الضوئي ، وهي بكتيريا كلورووكسي. يمكن افتراض أن بكتيريا cyano- و chlorooxybacteria هي أسلاف البلاستيدات الموجودة في خلايا الطحالب والنباتات. نظرًا لتراكم O2 نتيجة لعملية التمثيل الضوئي الميكروبي ، أصبح الغلاف الجوي للأرض مؤكسدًا ، مما تسبب في أزمة لجميع أشكال الحياة - حتى ذلك الوقت كانت البكتيريا اللاهوائية تسكن العالم. من أجل البقاء على قيد الحياة ، كان على الميكروبات تجنب الأكسجين عن طريق الاختباء في المستنقعات ، أو كان عليها تطوير عملية التمثيل الغذائي القادرة على حمايتها من كمية الأكسجين المتزايدة.
التصوير الضوئي والهواء
منذ ما يقرب من ملياري سنة ، حدثت عمليات تطورية مختلفة واحدة تلو الأخرى لخلق التنوع الأيضي. يمكن نقل القدرة على إنتاج إنزيمات معينة داخل نفس النوع باستخدام جزيئات تشبه الفيروسات تسمى البلازميدات. قدرات بدائيات النوى على التخليق الحيوي مدهشة: من ثاني أكسيد الكربون والعديد من الأملاح والمركبات غير العضوية المختزلة التي تعمل كمصادر للطاقة ، يمكن للكائنات الدقيقة ذات التغذية الكيميائية أن تصنع في الماء غير المشرق المحتوي على الأكسجين جميع الجزيئات الكبيرة الأكثر تعقيدًا اللازمة للنمو والتكاثر. أدت هذه العمليات البيوكيميائية إلى أنظمة دوران العناصر حتى في حالة عدم وجود نباتات أو فطريات أو حيوانات.
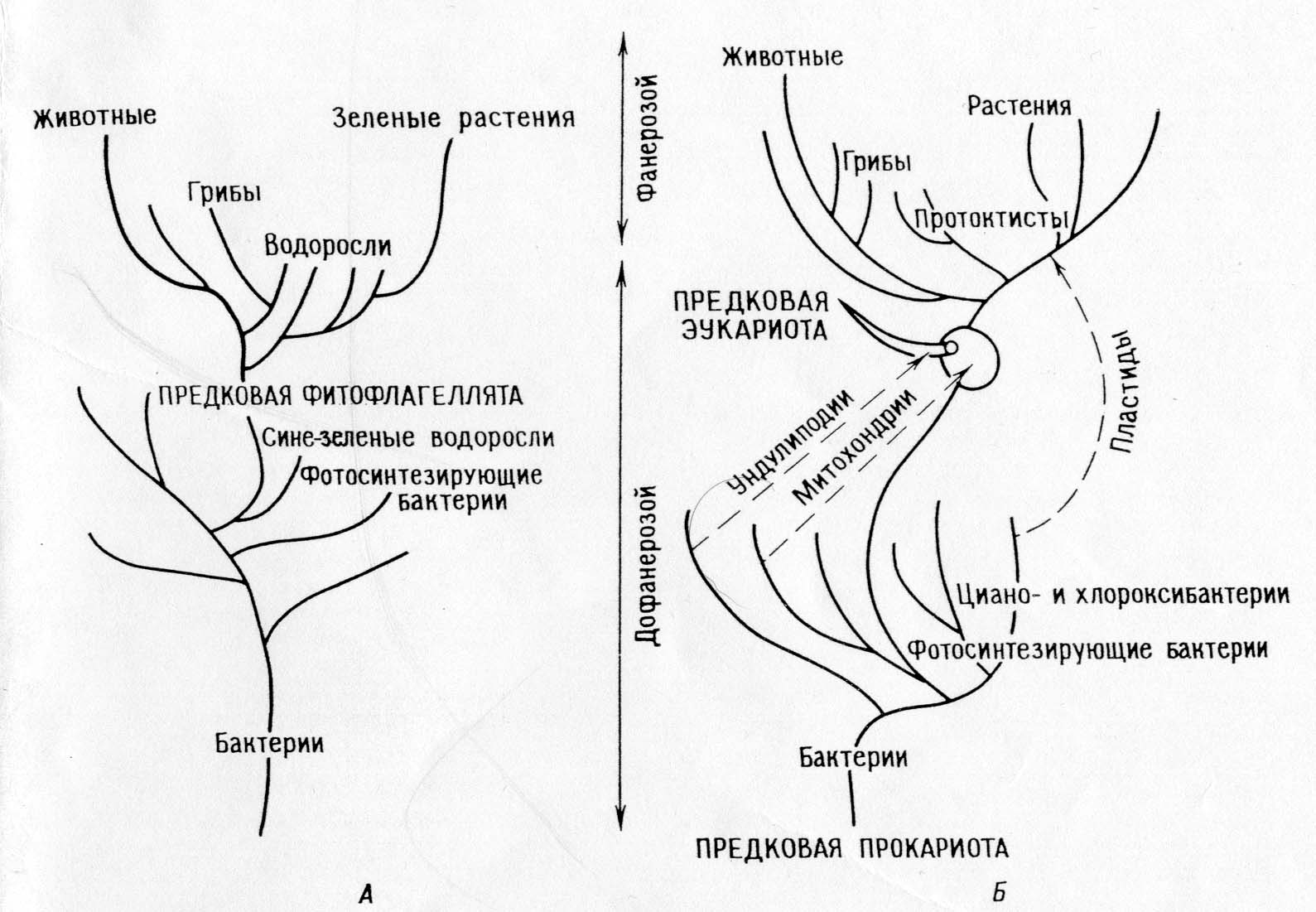
الجمعيات وحقيقيات النوى.
نظرة على تطور الخلية.
فرع مباشر.
النظرة التقليدية لأصل وتطور الخلايا حقيقية النواة هي أن جميع الكائنات الحية قد انحدرت مباشرة من مجموعة سلالة واحدة نتيجة لتراكم الطفرات تحت تأثير الانتقاء الطبيعي.
تعد الطفرات النقطية والازدواجية والحذف والتغيرات الوراثية الأخرى ، والتي من المعروف أنها لعبت دورًا في تطور الحيوانات والنباتات ، مسؤولة أيضًا عن تمايز الخلايا حقيقية النواة عن الخلايا بدائية النواة - نظرية البنوة المباشرة. أي مفهوم الأصل غير التكافلي للخلايا. ربما تكون الطحالب الخضراء من نسل أسلاف جميع النباتات. تشير العديد من سمات هذه الطحالب إلى أن لها وللنباتات سلف مشترك. تعتبر نسالة الحيوانات أكثر تعقيدًا ، ولكن ربما ينطبق مفهوم البنوة المباشرة على مساراتها التطورية أيضًا. يبدو أن الإسفنج نشأ من الطلائعيات ، بشكل مستقل عن مجموعات الحيوانات الأخرى. لذلك ، على الأقل بالنسبة للحيوانات والنباتات ، فإن الفرضية القائلة بأن الكائنات حقيقية النواة المعقدة قد تطورت من حقيقيات النوى المرتبة بشكل أكثر بساطة من خلال تراكم الطفرات الفردية تحت تأثير الانتقاء الطبيعي هي فرضية مشروعة تمامًا. تعتمد الابتكارات التطورية على الحفاظ على الجينات الملائمة المثالية للغاية. لذلك ، تطورت الكائنات الحية الجديدة ذات القدرة المتزايدة على التحكم في بيئتها عن طريق الحفاظ على تكرار الجينات شديدة التكيف وإعادة تركيبها عن طريق اكتساب خصائص جديدة من خلال طفرات نقطية بسيطة وعشوائية. لا تفسر فرضية التبويض المباشر أصل حقيقيات النوى من بدائيات النوى. على سبيل المثال ، يصعب فهم الفجوة بين البكتيريا الزرقاء غير القادرة على الانقسام ومثل هذه الأشكال ذات الانقسام الكامل مثل الطحالب الحمراء من وجهة نظر التبويض المباشر ، ولكنها تتبع بشكل طبيعي من النظرية التكافلية.
وفقًا لنظرية البنوة المباشرة ونظرية التكافل ، نشأت جميع الكائنات الحية على الأرض من أسلاف بكتيرية متشابهة كيميائيًا ، وأدت الميكروبات حقيقية النواة إلى ظهور النباتات والحيوانات. أدت بكتيريا التمثيل الضوئي إلى ظهور الطحالب ، وفي النهاية النباتات ، وفقدت بعض الطحالب بلاستيداتها وأصبحت أسلاف الفطريات والحيوانات. يتفق منظرو البنوة المباشرة على أن البلاستيدات والعضيات حقيقية النواة الأخرى ، بما في ذلك النواة ، تشكلت عن طريق التمايز داخل الخلايا نفسها ويسمي هذا أسطورة نباتية.
نسالة أنواع النباتات
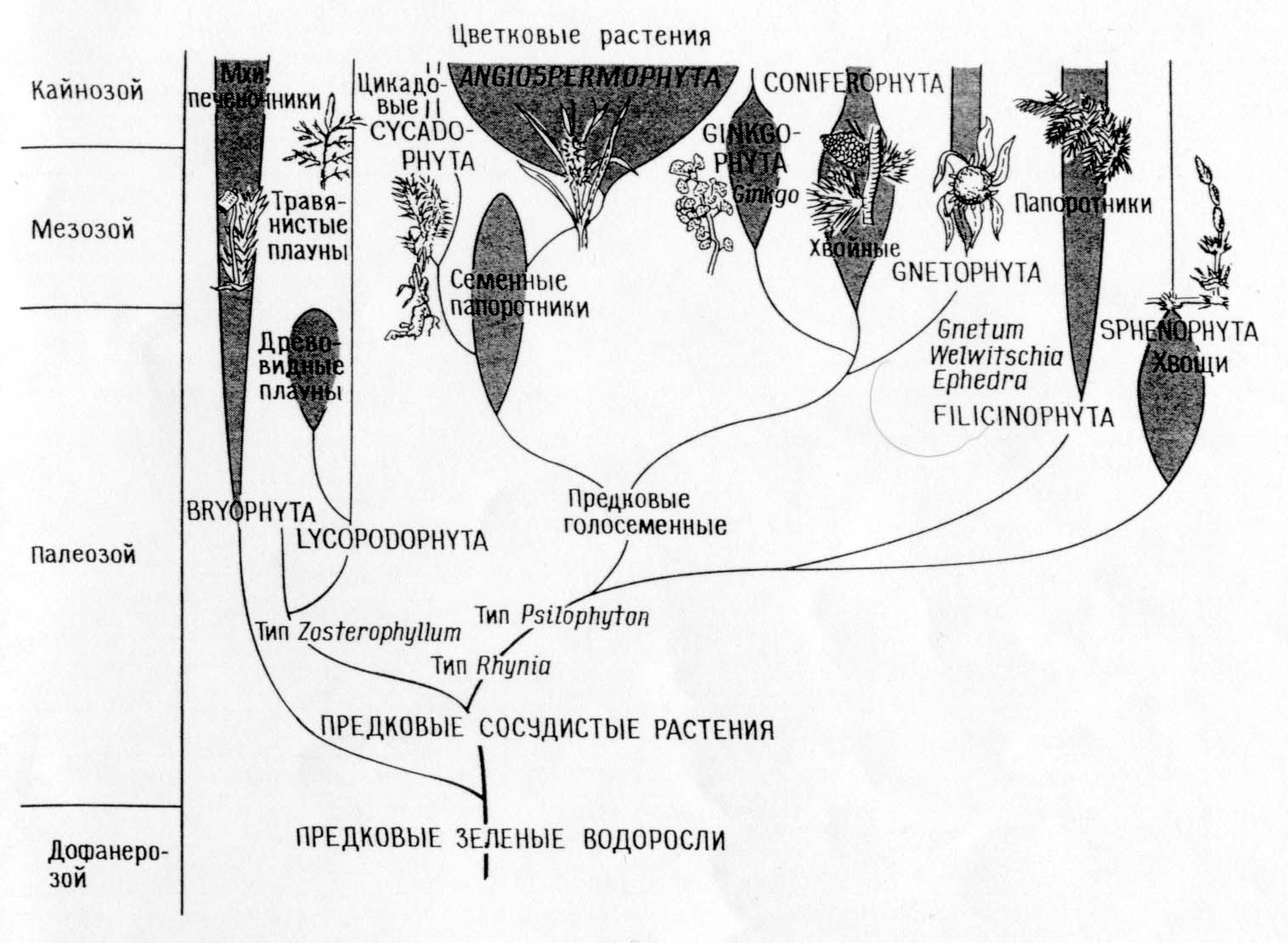
مقارنة بين السلالات المقابلة لنظرية البنوة المباشرة (أ)
ونظرية التعايش (ب)
الاختلافات بين نظرية البنوة المباشرة والنظرية التكافلية:
نظرية البنوة المباشرة.
الانقسام الرئيسي: الحيوانات - النباتات
تطورت حقيقيات النوى الضوئية (الطحالب والنباتات الخضراء) من بدائيات النوى الضوئية (الطحالب الخضراء المزرقة)
تطورت النباتات وأنظمتها الضوئية بشكل أحادي في عصر ما قبل الكمبري.
تطورت الحيوانات والفطريات من ذاتية التغذية فقدت البلاستيدات.
تتمايز الميتوكوندريا داخل الخلايا في طحالب التمثيل الضوئي.
في الطحالب اليمنى ، تمايزت أوندوليبوديا ، والجهاز الانقسام ، وجميع العضيات حقيقية النواة الأخرى داخل الخلايا.
تنحدر جميع الكائنات الحية من أسلافها - بدائيات النوى عن طريق تراكم الطفرات الفردية.
النتيجة: وفقًا للنظرية ، يجب أن يكون هناك أعشاب صحيحة ؛ من الصعب ربط هذه النظرية بسجل الحفريات وتكوين سلالة مناظرة.
نظرية التعايش الداخلي المتتالي:
الانقسام الرئيسي: بدائيات النوى - حقيقيات النوى.
تطورت حقيقيات النوى الضوئية (الطحالب والنباتات النووية) وحقيقيات النوى غير الضوئية (الحيوانات ، الفطريات ، البروتوزوا) من أسلاف مشتركة غيرية التغذية (أميبوفلاجيلات).
كان تطور التمثيل الضوئي في الأركيان أحادي الخلية في البكتيريا (بما في ذلك cyano- و chlorooxybacteria) ، نشأت الطلائعيات الضوئية فجأة عندما دخلت أسلافها غير المتجانسة في التعايش مع بدائيات النوى الضوئية (protoplastids).
تنحدر الحيوانات ومعظم الطحالب والفطريات مباشرة من الطلائعيات غيرية التغذية.
كانت الميتوكوندريا موجودة بالفعل في الطلائعيات غيرية التغذية عندما اكتسبت الأخيرة البلاستيدات نتيجة التعايش. تم الحصول على البروميتوكوندريا سابقًا بطريقة تكافلية.
تطور الانقسام المتساوي في الطلائعيات حقيقية النواة غيرية التغذية عن طريق التمايز بين النظام غير الشحمي.
تطورت بدائيات النوى من أسلاف بدائية النواة المبكرة من خلال سلسلة من الأحداث الطفرية الفردية ، كما تطورت حقيقيات النوى أيضًا من أسلاف حقيقية النواة من خلال أحداث طفرة واحدة ، وتطورت أسلاف حقيقية النواة من بدائيات النوى من خلال سلسلة من التعايش.
النتائج: تشرح النظرية المسارات البيوكيميائية وتكرارها في العضيات. إنه يسمح ببناء نسالة متسقة تشرح "الفجوة البيولوجية" بين العصر البدائي وعصر دهر الحياة. تتنبأ النظرية بأن العضيات يجب أن تحتفظ ببقايا أنظمة النسخ المتماثل.
أسطورة نباتية
من وقت Jussieu ، تسمى النباتات السفلية النباتات الطويلة ، وتسمى النباتات العليا النباتات الطحلبية والقصبة الهوائية. وفقًا للأسطورة النباتية ، فإن الأسلاف المشتركة للبكتيريا التي تعمل بالتمثيل الضوئي والطحالب الخضراء المزرقة ، من خلال التبويض المباشر ، أدت إلى ظهور طحالب حقيقية النواة ، بما في ذلك الطحالب البحرية. انتهى المطاف بمجموعات الكائنات الحية التي تأثرت نسختها بمفهوم البنوة المباشرة في مجال علم النبات - وهي البكتيريا والطحالب والفطريات والنباتات.
الكائنات الافتراضية التي يمكن أن تكون رابطًا بين البكتيريا الزرقاء والطحالب الخضراء كانت تسمى phytomonads الأسلاف (phytomonads الأجداد) أو الأعشاب الصحيحة. جادل بعض المؤلفين بأن Cyanidium و Cyanophora eukaryotes ، المتشابهة في أنظمة الصبغ الخاصة بهم مع البكتيريا الزرقاء ، قد تكون مرشحين مناسبين لدور الأعشاب الصحيحة. في النباتات الطويلة ، لم يتم العثور على أشكال وسيطة بين الحالة غير الانقسامية للبكتيريا الزرقاء والانقسام النموذجي المطور بالكامل للطحالب والنباتات الخضراء. على الرغم من أن الفطريات والنباتات المزهرة خالية تمامًا من الوحوش ، إلا أنها تتميز بالانقسام والانقسام الاختزالي ، والتي تتطلب مغازل الأنابيب الدقيقة. هناك اتفاق عام على أن جميع الكائنات متعددة الخلايا ذات الأنابيب الدقيقة تنحدر من الكائنات الحية الدقيقة حقيقية النواة ذات الأجنحة غير الشحمية ، ولكن لا أحد يعرف من أي مجموعة من الطلائعيات. كان تعزيز الأسطورة النباتية هو العصور القديمة وتعقيد عملية التمثيل الضوئي ، بالإضافة إلى حقيقة أن العديد من الكائنات الحية (البكتيريا ، الأوجلينيدات ، النباتات الطفيلية) ، سواء في الطبيعة أو في المختبر ، فقدت التمثيل الضوئي نتيجة للطفرات. نظرًا لأن عملية التمثيل الضوئي هي عملية لا هوائية تضمن في النهاية وجود جميع الكائنات الحية ، فيجب أن تكون قد تطورت في وقت مبكر جدًا من تاريخ الحياة على كوكبنا ، حتى قبل ظهور الانقسام الفتيلي ، الذي يتطلب الأكسجين. تشارك العديد من الأصباغ والدهون والإنزيمات في عملية التمثيل الضوئي ، مرتبة بطريقة منظمة في أغشية معقدة للغاية. في الوقت نفسه ، فإن تفاصيل آليات التمثيل الضوئي في الطحالب والنباتات ، من ناحية ، وفي البكتيريا الزرقاء من ناحية أخرى ، متشابهة بشكل لافت للنظر. تطلق كل هذه الكائنات غاز الأكسجين من الماء ، والذي يعمل كمصدر لذرات الهيدروجين المستخدمة لتقليل ثاني أكسيد الكربون إلى المادة العضوية للخلية. العديد من الطحالب والأوليات ليس لها انقسام ولا نظام وراثي مشابه لنظام الحيوانات والنباتات. تمتلك Ciliates نظامًا وراثيًا خاصًا به نوعان من النوى ، وغالبًا ما تكون الخلايا متعددة النوى. نوع واحد من النوى هو النوى الكبيرة ، والآخر أصغر بكثير من النوى. النوى الصغيرة غير مهمة من الناحية الفسيولوجية. ومع ذلك ، فإنهم يخزنون نسخًا من الجينات ، وأثناء العملية الجنسية ، يخضعون للانقسام الاختزالي. تعمل النوى الكبيرة من الناحية الفسيولوجية كموقع لنسخ الحمض النووي الريبي وبالتالي تتحكم في فسيولوجيا الخلية. تتمثل إحدى الصعوبات في توضيح العلاقة بين البكتيريا الزرقاء والطحالب حقيقية النواة في الافتقار إلى البيانات المتعلقة بأصل أوندوليبوديا: لم يتم العثور على روابط وسيطة بين البكتيريا الزرقاء ، التي لا تحتوي أبدًا على أوندوليبوديا ، والعديد من الطحالب التي تفعل ذلك. مشكلة أخرى هي الغياب التام للعملية الجنسية في البكتيريا الزرقاء: بين البكتيريا الزرقاء اللاجنسية والطحالب ، والتي يتميز معظمها بالعملية الجنسية ، لم تكن هناك أشكال وسيطة. تم العثور على معظم الطحالب لديها undolipodia في مرحلة ما من دورة حياتها. اتضح أن الهياكل المتنقلة في الطحالب والبروتوزوا والحيوانات والنباتات هي نفسها: كلها تتكون من أنابيب دقيقة يبلغ قطرها 0.024 ميكرون. اتضح أن المغزل الانقسامي ، حتى في الفطريات التي لا تحتوي على شحميات ، يتكون من أنابيب دقيقة بروتينية من نفس القطر كما هو الحال في undolipdia. لمدة نصف قرن ، بدا من المعقول للغاية أن الطحالب حقيقية النواة تطورت من البكتيريا الزرقاء. أدت هذه الأسطورة النباتية إلى افتراضات كانت لسنوات عديدة بمثابة حافز للبحث وإطار مرجعي تم من خلاله تقييم النتائج. أصبح التأكيد على أن الروابط بين البكتيريا الزرقاء والطحالب حقيقية النواة قد انقرضت ، ولم يترك أي آثار أحفورية وآثار حية ، وأصبح أقل إقناعًا مع توفر بيانات جديدة عن الميكروبات الحية والحفريات. لم تنقرض الروابط الوسيطة الرئيسية بين بدائيات النوى الضوئية وحقيقيات النوى بدون أثر ، كما هو مطلوب في الأسطورة النباتية - لم تكن موجودة أبدًا. وفقًا لنظرية التعايش الداخلي المتتالي ، فإن البكتيريا الزرقاء هي بالفعل الأسلاف ، ولكنها فقط البلاستيدات ، وليست بقية الخلية حقيقية النواة ، وبالتأكيد ليست نواتها. ومع ذلك ، في مسألة أصل النواة ، تتوافق فرضية البنوة المباشرة ونظرية التعايش. تتضمن التعديلات التي يمكن تصورها على النسخة المتطرفة من نظرية التعايش ، على سبيل المثال ، فكرة الأصل التكافلي للبلاستيدات ، ولكن ليس الميتوكوندريا ، أو الأصل التكافلي للبلاستيدات والميتوكوندريا ، ولكن ليس من أصل شحمي. طرح أنصار البنوة المباشرة أيضًا افتراضًا حول أصل النواة من خلال التعايش.
أدلة حاسمة على أصل التكافل الداخلي للبلاستيدات الخضراء والميتوكوندريا.
تم الحصول على أهم دليل على أصل أهم عضيات الخلايا حقيقية النواة - البلاستيدات الخضراء والميتوكوندريا من بدائيات النوى في السنوات الأخيرة عند دراسة بنية الحمض النووي الريبي الريبوسومي. عالم الأحياء الجزيئي
أظهر K. Woz بالفعل في الثمانينيات أن بنية الحمض النووي الريبي من ريبوسومات البلاستيدات الخضراء النباتية متطابقة تقريبًا مع بنية الحمض النووي الريبي من بعض البكتيريا الزرقاء. في الوقت نفسه ، وجدت مجموعة من الباحثين أن الحمض النووي الريبي للريبوزومات المشتق من الميتوكوندريا لحقيقيات النوى المختلفة يشبه الحمض النووي الريبي لبعض البكتيريا ، ولا سيما بكتيريا جنس Paracoccus. مجموعة إنزيمات الجهاز التنفسي لهذه البكتيريا تشبه إلى حد بعيد
"المجموعة التنفسية" من الميتوكوندريا الحيوانية.
تشهد هذه البيانات بوضوح تام لصالح الاحتمال الأكبر للأصل التعايش الداخلي للخلايا حقيقية النواة. لا تزال ظاهرة التعايش الداخلي منتشرة للغاية في الطبيعة. من المعروف أن البكتيريا المضيئة غالبًا ما تستقر على جسم بعض الأسماك البحرية ، والتي لا تتلقى فقط فوائد كبيرة لحياتها ، ولكنها أيضًا مفيدة جدًا للكائنات الحية المضيفة. كل ما سبق يشهد بشكل مقنع لصالح الأصل التعايش الداخلي للخلايا حقيقية النواة من خلايا بدائية النواة أصغر بكثير. ومع ذلك ، فإن السؤال الرئيسي هو تحديد طبيعة الخلية المضيفة ، لأن الخلايا حقيقية النواة لها نواة جيدة التكوين مع غلاف ، بينما لا تحتوي بدائيات النوى على مثل هذه النواة. يعتقد L.Margelis ، على سبيل المثال ، أن دور الخلايا المضيفة ربما تم لعبه بواسطة بدائيات النوى - أسلاف البكتيريا الحديثة ، التي تنتمي إلى الميكوبلازما - بدائيات النوى المبنية بشكل بدائي للغاية ، وخالية عمليًا من جدران الخلايا. في الآونة الأخيرة ، ظهرت أعمال العالم الياباني ونوقشت على نطاق واسع.
تي أوشيما. استنادًا إلى دراسة شاملة لبنية الحمض النووي الريبي الريبوسومي للعديد من الكائنات الأولية وحقيقيات النوى ، بما في ذلك العديد من ممثلي مملكة العتائق التي لم تتم دراستها كثيرًا حتى الآن ، استنتج هذا المؤلف أن الخلية المضيفة كانت أحد أسلاف العتائق الحديثة. يجب أن يقال أن ممثلي هذه المملكة الفائقة ، على عكس البكتيريا ، لديهم الكثير من السمات البيولوجية البيوكيميائية والجزيئية المشتركة مع حقيقيات النوى. في أحدث أعمال T. Oshima ، تم تقديم الحجج لصالح حقيقة أن الخلية المضيفة في وقت ظهور حقيقيات النوى كانت سلف العتائق ، على غرار أحد الممثلين المعاصرين لهذه بدائيات النوى - البلازما الحرارية. ماذا سيكون الحل النهائي لمشكلة تحديد طبيعة الخلية المضيفة التي أدت إلى ظهور الخلية حقيقية النواة في عملية التعايش الداخلي المتعدد ستظهر من خلال مزيد من الدراسات.
يتم حاليًا تقديم الفرضية الأكثر شيوعًا والأكثر إثباتًا لأصل الخلايا حقيقية النواة
التعايش الداخلي للعديد من بدائيات النوى التي تنتمي إلى البكتيريا والعتائق ، والتي حدثت عدة مرات في عملية التطور خلال أول 1.5-2 مليار سنة من وجود الخلايا الحية على الأرض. تم الحصول على دليل حاسم على أصل التكافل الداخلي للبلاستيدات الخضراء والميتوكوندريا من قبل علماء الأحياء الرائدين في العالم في دراسة بنية الحمض النووي الريبي الريباسي. تشهد هذه البيانات بوضوح تام لصالح الاحتمال الكبير للأصل التعايش الداخلي للخلايا حقيقية النواة من خلايا بدائية النواة أصغر بكثير (بعدة أوامر من حيث الحجم).
الأدب
مجلة سوروس التعليمية. علم الأحياء والكيمياء وعلوم الأرض والفيزياء والرياضيات. العدد 5 1998
مارجليس ل. دور التعايش في تطور الخلية. لكل. من الإنجليزية M. MIR 1983
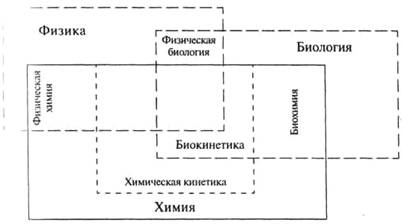
من الأقسام الرئيسية للخواص الحركية ، يدرس حركية التفاعلات البيولوجية ؛ هذا القسم يسمى الحركية الحيوية. علم الحركية الحيوية هو علم حدودي ظهر عند تقاطع الكيمياء الحيوية والحركية الكيميائية (انظر الشكل 1). إن اختيار الحرائك الحيوية كنظام منفصل ليس عرضيًا ، بل له ما يبرره منطقيًا ويرتبط بالأهمية الاستثنائية للعمليات الحركية لجميع الكائنات الحية. الحرائك الحيوية ...
الجينات ، ولكنها خلقت أيضًا المتطلبات الأساسية لتراكم التغييرات فيها ، وتباعد الجينات ، وزيادة تنوع المنتجات التي تتحكم فيها. 2. العناصر الوراثية المتنقلة ينتمي دور معين في تطور جينومات كل من الخلايا الأولية والخلايا حقيقية النواة إلى ما يسمى بالعناصر الوراثية المتنقلة - الينقولات. إنها وحدات مستقلة تحمل النيوكليوتيدات ...
عديد الطرق الممكنةلتنمية المستقبل النظرية العامة الأنظمة الديناميكية. في الفصل العاشر ، العمليات الدائرية للحياة ، نعتبر التنظيم الدوري المميز للتنظيم الذاتي أنظمة مشتتة. يضع المخطط المعمم خصائص توليد دورات التحويل ودورات التفاعلات التحفيزية ، بالإضافة إلى الدراجات الفائقة التحفيزية وفقًا ...