פילוגניה, פילוגניה - התפתחות היסטוריתעולם האורגניזמים החיים, הן בכלל והן בקבוצות טקסונומיות בודדות: ממלכות, טיפוסים (חלוקות), מחלקות, סדרים (סדרים), משפחות, סוגים, מינים. המונח פילוגניה הוצג על ידי E. Haeckel.
המערכת המודרנית של בעלי חיים, הבנויה על עקרונות הקרבה, יכולה להיות מיוצגת כעץ גנאלוגי (או פילוגנטי), המשקף את השלבים העיקריים של התפתחות בעלי החיים. פילוגנזה קבוצות שונותאורגניזמים שנחקרו בצורה לא אחידה. הפילוגניה של בעלי חוליות (במיוחד של הקבוצות הגבוהות יותר) נחקרה בצורה הטובה ביותר; מבין חסרי החוליות, הפילוגניה של רכיכות, אכינודרמים ופרוקי רגליים נחקרה בצורה הטובה ביותר. הפילוגניה של פרוקריוטות מובנת בצורה גרועה.
בבסיס העץ האבולוציוני של בעלי החיים הם חד-תאיים. באורגניזמים חד-תאיים, כל הפונקציות של הגוף מבוצעות על ידי תא אחד. ישנן מספר השערות לגבי המקור אוקריוטים חד תאיים s: 1) אוקריוטים התפתחו מפרוקריוטים על ידי סימביוגנזה. השערה זו נתמכת על ידי העובדה שלמיטוכונדריה אוקריוטים ולכלורופלסטים יש DNA משלהם; 2) אוקריוטים התרחשו עקב התמיינות של תאים פרוקריוטים לציטופלזמה ולאברונים. כל סוגי הפרוטוזואה המודרניים הם צאצאים של sarcoflagellates עתיקים.
בעלי חיים רב תאיים, מאורגנים יותר, שבהם פונקציות בודדותאורגניזמים מבוצעים על ידי תאים או איברים בודדים. כל האורגניזמים הרב-תאיים בהתפתחות אינדיבידואלית מתחילים בתא בודד - זיגוטה, ו בשלבים הראשוניםהעובר (בלסטולה) דומה למושבה חד-תאית.
קיימות מספר השערות למקורן של בעלי חיים רב-תאיים. הידועים ביותר הם ההשערות של E. Haeckel ו-I.I. מכניקוב. על פי התיאוריה של האקל, כל החיות הרב-תאיות צאצאיות מצורת אבות אחת - גסטרה. I.I. מכניקוב הניח כי האב הקדמון של בעלי חיים רב-תאיים הוא מושבה של דגלים. ראשון אורגניזם רב תאיהיה בעל צורה כדורית, היה מכוסה בשכבת תאים עם דגלים והיו להם תאי אמבוי עיכול בתוך הגוף, צפו בחופשיות במים. I.I. מכניקוב קרא לזה phagocytella. התפתחות נוספת של אורגניזמים רב-תאיים התנהלה בארבעה כיוונים. אחד מכיווני האבולוציה הוביל להיווצרות ה-Lamellar Type, המורכב משכבה אחת של תאים ומוביל אורח חיים זוחל. כיוון נוסף של אבולוציה הוביל להיווצרות של Sponge Type עם שתי שכבות של תאים שלא נוצרו לרקמה. אין להם עדיין מערכת עצבים. אלה הם בעלי חיים בנטיים חסרי תנועה. הכיוון השלישי של האבולוציה הוביל להיווצרות ה-Type Coelenterates. Coelenterates הם כבר בעלי חיים דו-שכבתיים אמיתיים (אקטודרם ואנדודרם) עם מעי ראשוני, פה, סימטריה רדיאלית, מערכת עצבים מפוזרת. ביניהם יש חיות שחיה חופשיות ומנהלות אורח חיים צמוד. הכיוון הרביעי של האבולוציה הוביל להיווצרותם של בעלי חיים תלת-שכבתיים מאורגנים אפילו יותר. הרקמות והאיברים של בעלי חיים אלו נוצרו משלוש שכבות של תאים (שכבות נבט) אקטודרם, אנדודרם ומזודרם.
התלת-שכבתיים הראשונים היו בעלי חיים הדומים למשטחי מעיים. האבולוציה של בעלי חיים תלת-שכבתיים הלכה בשלושה כיוונים. כיוון אחד הוביל לגיבוש תולעים שטוחות, שבו אין חלל גוף, והפרנכימה ממוקמת בין האיברים. תולעים שטוחות הן החיות הדו-צדדיות הראשונות (לכל הסוגים הבאים יש סימטריה זו). הם מופיעים לראשונה מערכת השרירים(חלק), גנגלי עצב. עם זאת, המעי שנוצר הוא סגור עיוור, אין לו פי הטבעת.
כיוון אחר הוביל להיווצרותן של חיות חלל ראשוניות תולעים עגולות. המעיים שלהם הופכים מקצה לקצה, מורכב משלושה חלקים עם פִּי הַטַבַּעַת. יש ריכוז נוסף תאי עצבים, יצירת טבעת עצב פריפרינגיאלית. המשותף לתולעים שטוחות ול-annlids הוא נוכחות של שק עור-שרירי, מערכת העיכול, ההפרשה ומערכות הרבייה.
הכיוון השלישי של האבולוציה של התלת-שכבות מוביל להיווצרות של בעלי חיים מנוולים. היווצרות של חלל גוף משני סיפקה ניידות ויציבות רבה יותר סביבה פנימיתאורגניזם. חללים משניים מאורגנים יותר: יש להם מערכת זרימת דם. אלה כוללים את כל חסרי החוליות האחרים והקורדטים.
האבולוציה של הדאוטרוסטומים הובילה להיווצרותם של פרוטוסטומים ודוטרוסטומים. אבולוציה של פרוטוסטומים. מקורם של רכיכות ואנלידים ראשוניים מהאב הקדמון של הפרוטוסטומים. בתורם, מקורם של annelids ופרוקי רגליים מודרניים מהאנלידים העיקריים.
לאנלידים יש מאפיינים ארגוניים מתקדמים: חלל גוף משני, גוף מנותח בצורה מטאמרית, מערכת זרימת דם סגורה (לא סגורה בחסרי חוליות אחרים), טבעת היקפית ושרשרת עצבים.
פרוקי רגליים משיגים שלמות גדולה יותר: הלב מתעורר במערכת הדם; במערכת העצבים, ריכוז הגרעינים עולה; נוצר שלד חיצוני(כיסוי כיטין); מגוון איברי הנשימה (זימים, קנה הנשימה, ריאות) אופייני; נוצרים איברים וכנפיים מפרקים. מבנה מושלם יותר של פרוקי רגליים אפשר להם לאכלס באופן נרחב את כדור הארץ.
לרכיכות יש חלל גוף משני, מורכב איברים פנימיים(נוצרים כליות, לב תלת חדרים, זימים, ריאות), לרובם יש מעטפת גירנית המגנה על הגוף. עם זאת, ברכיכות, מערכת הדם אינה סגורה, ומערכת העצבים היא מסוג נודולרי מפוזר.
אבולוציה של Deuterostome. Echinoderms ו-chordates צאצאי אבותיהם של deuterostomes.
Echinoderms. הם מאופיינים ב: חלל משני של הגוף, מערכת הדם יוצרת פערים, מעין מערכת מים-מוטורית (אמבולקרלית), סימטריה רדיאלית, מערכת עצבים בצורת טבעת עם קרניים רדיאליות.
כורדיטים ראשוניים כנראה התפתחו מאבות קדמונים דמויי תולעת דוירוס. הבידוד של chordates קשור לעלייה בניידות. היחסים הפילוגנטיים של chordates עם חסרי חוליות מעידים על הדברים הבאים: 1) רקמות ואיברים נוצרים משלוש שכבות נבט (הנוצרות לראשונה בתולעים שטוחות); 2) סימטריה דו-צדדית (מתרחשת בתולעים שטוחות); 3) חלל גוף משני (מתרחש ב-annelids); 4) פה משני (קשור לאכינודרמים); 5) מערכת מחזור סגורה (annelids);
מהקורדאטים הראשוניים, מקורם של בעלי חיים מודרניים שאינם גולגולתיים (זמל) ובעלי חוליות. כורדאטים זחלים הם ענף צדדי של כורדאטים, אשר ברובם עוברים למחזור חיים בישיבה, ולכן יש להם סימני כורדאטים רק בשלב הזחל.
קפלותורדטים הם אקורדים פרימיטיביים. יש להם נפרידיה, בדומה למטנפרידיה של אנלידים, מערכת מחזור סגורה ללא לב, כמו אנלידים.
אבותיהם של בעלי החוליות הופיעו לפני כ-500 מיליון שנה. האבולוציה שלהם התנהלה בשני כיוונים: חסרי לסת (ציקלוסטומים) ולסתות (כל שאר בעלי החוליות).
דג הלסת הראשון - משוריין התפתח משיריון ללא לסת. כבר היה להם חזק שלד פנימי, גולגולת מוחית וקרביים, לסתות עם שיניים, סנפירים מזווגים. הדגים האלה נכחדו. הם הוחלפו בסחוס ו דג גרמישגופו היה מכוסה בקשקשים.
דגי סחוס היו נפוצים בים, ולפי סוג המזון הם היו טורפים.
דגים גרמיים הופיעו במקור במים מתוקים. הם פיתחו אופרקולום (מגן על הזימים מפני סתימה) ושלפוחית שחייה (מקלה על שחייה במים מתוקים). בייבוש מקווי מים (בפליאוזואיקון) הופיעו דגי אונה (דג ריאה ואונה סנפיר), שבהם בנוסף לזימים התפתחו שקי ריאה לנשימה באוויר, והשרירים שנוצרו על הסנפירים אפשרו להישען. על מצע מוצק. לאחר מכן, דגים גרמיים חיו במקווי מים מתוקים וימיים. כיום, דגים גרמיים הם אחת הקבוצות המשגשגות ביותר של בעלי חוליות (20 אלף מינים).
דו-חיים הופיעו בדבון. דו-חיים צאצאי דגים עתיקים עם אונות סנפיר. הדו-חיים הראשונים היו Ichthyostegidae (דג בעל ארבע רגליים). להם, כמו לדגים, היה סנפיר זנב, כיסוי זימים, קשקשים. אבל בניגוד לדגים, היו להם שני זוגות של גפיים וריאות עם חמש אצבעות. הם חיו בעיקר במים.
מאיכטיוסטג הגיעו דו-חיים אמיתיים - סטגוקפלים (בעלי ראש קונכייה). ראשם של סטגופליאנים היה מכוסה במעטפת של עצמות עור. סטגוצפלים חיו עד תחילת תקופת הקרטיקון. הם כנראה הולידו דו-חיים מודרניים ללא שריון ואת הזוחלים הראשונים.
הנחיתה לוותה בארומורפוזה: נשימת זימים השתנתה לנשימה ריאתית, וסנפירים לגפיים עם חמש אצבעות, הלב נעשה תלת-חדרי, נוצר מעגל שני של זרימת דם, אברי החישה השתנו ומערכת העצבים הפכה מושלמת יותר.
הזוחלים הראשונים הופיעו בפחמן. לשרידי המאובנים של סימוריה היו מאפיינים של דו-חיים וזוחלים. סביר להניח שהזוחלים הראשונים, הקוטילוזאורים, מקורם בצורות כאלה. הייתה להם גם גולגולת בצורת קופסת עצם (בדומה לסטגוקפלים) עם חורים לעיניים, לנחיריים ולאיבר הקודקוד. תקופת הזוהר של הזוחלים הייתה במזוזואיקון.
מתרחשת: קרטיניזציה של האפידרמיס והיווצרות קשקשים, או סקוטים (מגן על הגוף מפני התייבשות), סיבוך של מבנה הריאות, הלב, שיפור השלד, היווצרות קרומי הנבט של הסרוס. ומי שפיר (בתנאי שניתן להתרבות ביבשה).
זוחלים הולידו שני סוגים חדשים של חולייתנים: יונקים וציפורים.
ציפורים יצאו מהציפורים הראשונות מקבוצת הדינוזאורים הלטאות שרצה הלאה רגליים אחוריות. מבין הציפורים הראשונות העתיקות, נמצאו שרידי הארכיאופטריקס ארוך הזנב. לארכיאופטריקס היו סימנים של זוחלים (זנב ארוך, שיניים על הלסתות, טפרים על הכנפיים) וציפורים (נוצות, כנפיים, מראה). ציפורים אמיתיות הופיעו מהציפורים הראשונות בסוף תקופת המזוזואיקון. היו להם גם שיניים קטנות.
תכונות פרוגרסיביות של ציפורים: דם חם, לב בעל ארבעה חדרים, קשת אבי העורקים אחת (יוצר הפרדה מלאה של מחזור הדם המערכתי והריאתי).
יונקים הופיעו מוקדם יותר מאשר ציפורים. ההנחה היא שמקורם בלטאות בעלות שיניים של בעלי חיים, שהיו להן מספר תכונות פרוגרסיביות: השיניים ממוקמות במככיות, בחך העצמות המשני, הגפיים ממוקמות מתחת לגוף, ולא בצדדים, היו שינויים בשלד. צינודנטים היו הקרובים ביותר ליונקים מבין הזוחלים בעלי השיניים של היונקים. המשך האבולוציה של היונקים קשור לשינוי במערכת השיניים. שינויים אלו הובילו להיווצרותן של שתי קבוצות של בעלי חיים. צאצאים של קבוצה אחת - רב פקעות היו חיות קטנות, היו בעלי פקעות רבות על טוחנות, חותכות מפותחות מאוד ולא היו להן ניבים. ייתכן שקבוצה זו הולידה מונוטרמים.
הקבוצה השנייה התפתחה בצורה הדרגתית יותר. אלו היו חיות קטנות שאכלו חרקים, חיות קטנות. בעתיד, קבוצה זו מחולקת לשני ענפים עצמאיים - התחתון, כיס והגבוה, שליה.
חיות כיס התעוררו בתקופת הקרטיקון. הקבוצה הקדומה ביותר של חיות הכיס היא משפחת הפוסומים.
יונקי שליה הופיעו גם בתקופת הקרטיקון. קבוצת השליה הקדומה ביותר נחשבת לחוליית אוכלי החרקים. ההנחה היא שהם הולידו את רוב קבוצות השליה הבאות. יתכן שהפרימטים צאצאו ישירות מאוכלי חרקים.
אבולוציה מתקדמת של יונקי סטלן עם רכישת ההתאמות החשובות ביותר: דם חם, קצב חילוף חומרים גבוה, היווצרות השליה, האכלת הצעירים בחלב, טיפול בצאצאים. זה הוקל על ידי שינויים מתקדמים בדרכי הנשימה, מחזור הדם, מערכות עצבים. ליונקים יש לב בן ארבעה חדרים, קשת אבי העורקים אחת, עורקי ו דם ורידי, ריאות של מבנה alveolar, קמרון העצבים המשני - neopalium מפותח היטב.
13... המבנה של תא פרוקריוטי. צמיחה, התפתחות, רבייה של חיידקים. סוגי מזון של חיידקים. פוטוסינתזה, כימוסינתזה, פוטוסינתזה. ערכם של מיקרואורגניזמים בטבע ובכלכלה הלאומית תאים פרוקריוטייםכוללים חיידקים ואצות כחולות ירוקות. בְּ פרוקריוטיםלחומר הגרעיני אין קרום גרעיני; קרום הפלזמה שלהם מכיל פחמימות וחומצות אמינו. לפרוקריוטים יש רק כרומוזום אחד, המורכב רק מ-DNA. לכרומוזום זה צורת טבעת ואינו מכיל חלבונים אלקליים - היסטונים. קבוצת הכרומוזומים בפרוקריוטים היא הפלואידית.אורגניזמים חד-תאיים אלו חסרים מיטוזה ואין להם מנגנון מיטוטי. לפרוקריוטים אין מיוזה, ובמהלך התהליך המיני מועבר רק חלק מהמידע התורשתי, והאורגניזם עצמו הוא הגמטה. DNA ציטופלזמי בפרוקריוטים ממוקם בפלסמידים, וממברנות ציטופלזמיות נעדרות. דופן התא של פרוקריוטים מכיל פפטידוגליקנים. מיטוכונדריה אינן נצפות בפרוקריוטים, אנזימים נשימתיים ממוקמים במזוזומים, הממוקמים על פני השטח הפנימיים של קרום הפלזמה.תהליכי פוטוסינתזה מתבצעים באמצעות פיגמנט הבקטריוכלורופיל, ותרכובות גופרית משמשות כחומרים מפחיתים. ריבוזומים פרוקריוטיים קטנים יותר מריבוזומים אוקריוטיים. אין תנועה של הציטופלזמה; ואקואולים נדירים בה מאוד. הדגלים של הפרוקריוטים הינם תת-מיקרוסקופיים בגודלם, הדגלים מכילים מיקרוטובוליה אחת או יותר (סיבים) פרוקריוטים רגישים לאנטיביוטיקה ועמידים בפני קרני רנטגן ו טמפרטורה גבוהה, עמיד בחימום עד + 75 ... + 90 ° С. תכונה אופייניתפרוקריוטים הם שהם אינם מסוגלים להתקיים יחד עם סוגים אחרים של תאים. בַּקטֶרִיָה- אורגניזמים חד תאיים; ניתן לראות אותם רק במיקרוסקופ. לכן, הם נקראים מיקרובים או מיקרואורגניזמים. חיידקים שלטו במגוון רחב של בתי גידול: הם חיים באדמה, אבק, מים, אוויר, על הכיסויים החיצוניים של בעלי חיים וצמחים ובתוך הגוף. ניתן למצוא אותם אפילו במעיינות חמים, שם הם חיים בטמפרטורות של בסביבות 60 מעלות צלזיוס ומעלה. הפעילות של מיקרואורגניזמים היא חֲשִׁיבוּתלכל שאר היצורים החיים, שכן חיידקים ופטריות הורסים חומר אורגני ומשתתפים במעגל החומרים בטבע. בנוסף, חיידקים רוכשים הכל ערך גדול יותרבחייהם של אנשים, ולא בגלל שהם גורמים למחלות שונות, אלא בגלל שניתן להשתמש בהם כדי להשיג מוצרים נחוצים רבים.
מבנה התא. פרוקריוטי: לא נוצר גרעין, מיטוכונדריה, פלסטידים. במרכז הציטופלזמה נמצא נוקלואיד המכיל "כרומוזום" אחד עם מולקולת DNA מעגלית; אינו מופרד מהציטופלזמה על ידי ממברנה. בתוך התא, הצמחים של הממברנה מבצעים את הפונקציות של ה-EPS, מנגנון הגולגי; המזוזום מעורב בנשימה; יש ריבוזומים. הציטופלזמה מכוסה על ידי קרום חיצוני ודופן תא צפופה. לפעמים התא שקוע בקפסולה קולואידית נוזלית למחצה. על פי מבנה דופן התא ניתן לחלק את החיידקים לשתי קבוצות, חלקם צבועים גראם, לכן הם נקראים גרם חיוביים, בעוד שאחרים הופכים חסרי צבע כאשר הצבע נשטף ולכן הם נקראים גרם שליליים. צמיחה ורבייה.צמיחה אינדיבידואלית ורבייה א-מינית של תאים.יחס פני השטח/נפח של תאי חיידקים גבוה מאוד. זה תורם השתלטות מהירה חומרים מזיניםמ סביבהבאמצעות דיפוזיה ותחבורה אקטיבית. IN תנאים נוחיםחיידקים גדלים מהר מאוד. הצמיחה תלויה בעיקרעל הטמפרטורה וה-pH של המדיום, זמינות חומרי הזנה וריכוז היונים. גם אירובים מחוייבים צריכים חמצן, ואנאירובים מחויבים, להיפך, צריכים שהוא ייעדר לחלוטין. לאחר שהגיע לגודל מסוים, החיידקים עוברים ל רבייה א-מינית(ביקוע בינארי), כלומר, הם מתחילים להתחלק עם היווצרות של שני תאי בת. המעבר לחלוקה מוכתב על ידי היחס בין נפח הגרעין לנפח הציטופלזמה. לפני חלוקת התא מתרחשת שכפול דנ"א, שבמהלכו המזוזומים מחזיקים את הגנום במיקום מסוים. מזוזומים יכולים להיצמד גם למחיצות חדשות בין תאי הבת ואיכשהו להשתתף בסינתזה של חומרי דופן התא. החיידקים הגדלים הכי מהר מתחלקים כל 20 דקות.
4 סוגי תזונה של חיידקים וחלק ממאפייניהם.
ידועות שלוש קבוצות עיקריות פוטוסינתזה(חיידקים שמקבלים אנרגיה מהשמש): 1. חיידקי גופרית ירוקה . חיידקים אנאירוביים המשתמשים במימן גופרתי (H2S) ובתרכובות גופרית מופחתות אחרות כתורמי מימן ואלקטרונים.
דוגמה: אור
2H2S + CO2 →→→→→→→→→ [CH2O] + 2S + H2O.
בקטריוכלורופיל
במקרה זה, גופרית מופקדת.
2. חיידקי גופרית סגולים. לתאים יש צבע אדום כהה, שכן יש הרבה פחות בקטריוכלורופיל מאשר פיגמנטים אדומים וחומים. אלו הם בעיקר אנאירובים; הפוטוסינתזה בהם מתרחשת באותו אופן כמו בקבוצה הקודמת.
3. חיידקים סגולים שאינם גופרית . עבור חיידקים אלו, תרכובות אורגניות שונות משמשות כמקור למימן, המשמש להפחתת CO2 (בפוטו-אוטוטרופים) או תרכובת אורגנית כלשהי (בפוטו-הטרוטרופים).
כימוסינתזהאורגניזמים (כימואוטוטרופים) הם חיידקים המשתמשים ב-CO2 כמקור פחמן, אך מקבלים אנרגיה לא מהשמש, אלא בעזרת תגובה כימית. אנרגיה יכולה להשתחרר, למשל, במהלך חמצון של מימן, מימן גופרתי, גופרית, ברזל (II), אמוניה, ניטריט ותרכובות אנאורגניות אחרות. בחיידקי ברזל : חַמצָן
Fe2+ →→→→→ Fe3+ + אנרגיה;
בחיידקי גופרית חסרי צבע : חמצן או
S→→→→→→→→ SO42-+ אנרגיה;
גופרית סולפט
בתנאים אנאירוביים, מינים מסוימים משתמשים בחנקה כמקבל מימן ובכך מבצעים דניטריפיקציה.
בחיידקים מחנקים: חמצן
2NH4+→→→→→ NO-2 + אנרגיה
אמוניה ניטריט
כל החיידקים שנחשבו זה עתה הם אירוביים, מכיוון שחמצן משמש כמקבל אלקטרונים (ומימן).
חיידקים כימוסינתטיים ממלאים תפקיד חשוב מאוד בביוספרה; הם מעורבים בעיקר במחזור החנקן ובכך שומרים על פוריות הקרקע.
Photoreduction - עבור serobacter ירוק וסגול, שאינם משתמשים ב-H2O, אלא ב-H2 S כמקור
חשיבות בטבע ובפעילות כלכלית אנושית.
חיידקי תסיסה לקטית(ספרוטרופים הטרוטרופיים) המפרקים פחמימות: חִיוּבִי:הכנת מוצרי חומצה לקטית (גבינת קוטג', חלב מכורבל, חמאה, שמנת חמוצה), הזנת ספיגה, כרוב מחמצת, מלפפונים כבישים ועגבניות. שלילי:קלקול של מוצרים.
חיידקי תסיסה אצטית(ספרוטרופים הטרוטרופיים) המפרקים פחמימות: חִיוּבִי:חמצון אלכוהול ל חומצה אצטית, המשמש לכבישה, שימורים של פירות וירקות. שלילי:קלקול של מוצרים.
חיידקי ריקבון(הטרוטרופים) המפרקים חלבונים: חִיוּבִי:תפקיד סניטרי - מינרליזציה של שאריות אורגניות. שלילי:קלקול מזון; כדי למנוע ריקבון, ייבוש, המלחה, כבישה, עיקור, פסטור וסוכר משמשים.
חיידקי גושים(סימביונטים הטרוטרופיים): חיידקי הגושים rhizobium חודרים לשורשים של צמחי קטניות (תלתן, תורמוס, אספסת ועוד) ונכנסים איתם לסימביוזה. כתוצאה מכך נוצרים גידולים על השורשים - גושים מלאים בחיידקים, המסנתזים תרכובות חנקן מחנקן אטמוספרי, הזמינות הן לצמח המארח והן לצמחים אחרים. העשרה טבעית זו של הקרקע בדשני חנקן נלקחת בחשבון בעת עריכת מחזורי גידולי שדה, הכוללים בהכרח קטניות.
14. מבנה של תא איקריוטי. קרום תלת שכבתיריבוזומיםמתחם גולגימיטוכונדריהליזוזומיםכלורופלסטיםכרומופלסטיםLeucoplastsמרכז תא או צנטרוזוםתכליליםואקווליםציטושלדקריופלזמהגרעין1. 2 3 4 5
15.אונטוגניה ותקופותיה. עוברים מוקדמים. פיתוח ישיר ועקיף.(51)
אונטוגנזה- ההתפתחות האישית של האורגניזם מלידה ועד מוות. זה ימשיך עבור ארגונים שונים אינו זהה ויכול להימשך מכמה מחשבות במשך מאות שנים (צב). אונטוגניה קשורה קשר הדוק לפילוגניה אך אינה חזרה מוחלטת של פילוגניה. בהשפעת הסביבה היא משתנה ויכולות להתרחש שונות של מוטציות. תקופות של אונטוגניה:
1 טרום עוברי
2. נבט (עוברית)
3. פוסט-עובר
בתקופה 1 התפתחות תאי איברי המין והכנתם להמשך התפתחות לאחר הפריה (spermatogenesis ו-ovogenesis) spermatogenesis מ-4 תקופות: 1. רבייה; 2. צמיחה; 3. הבשלה; 4. עיצוב
Spermatogenesis: Spermatogonia 2n "spermatocytes of the 1st order 2n" spermatocytes 2 pores 2n "spermatozoa n.
אוווגנזה: O vogonia 2n "ביציות 1 pore2n" ביציות 2 pore2n "ביצה אחת + 3 גופים מכוונים (מופחתים)
בתקופה השנייה, התפתחות העובר התרחשה ברחם (ברחם האם) או בסביבה החיצונית. תהליכים מהירים של הר-שיה של עיצוב, עיבוד טווח קצרלהוביל להופעתם של ריבוי org-ma. יש 3 מחזורים: 1. ראשוני- שבוע 1 התפתחות עובריתמתחיל מרגע ההפריה ומסתיים בהשתלת העובר ברירית האם. 2 נבט.זמן מההתחלה 2 עד 8 שבועות מההשתלה ועד להשלמת האורגנוגזה. 3 עוברים- התפתחות העובר מ-9 שבועות ועד לידה. צמיחה משופרת, עיצוב, תזונה המטוטרופית.
מחזור 3 לאחר ebr מתחיל מרגע הלידה, כלומר. שחרור האורגניזם מנבט הקליפות. במהלך תקופה זו של התפתחות, התעוררה תכונה אנטומית ותפקודית ספציפית, הנקראת גיל.תקופתיות:
1-10 ימים - יילוד
11 ד - תינוק בן שנה
1g-11 ילדים (בנות ובנים)
11-15י (ד) ו-12-16 (מ) - נער
16-20 (ד) ו-17-21 (מ) - נוער
21-55 (מ') ו-22-60 (מ') - בוגר
56, 61-74 - קשישים
75-90 סנילים, מעל 90 בני מאה
התפתחות ישירה (לא זחל), הקשורה למספר החלמון בביצה, או להתפתחות של org. באורגניזם של האם, הוא אופייני לדגים, יונקים, ציפורים, זוחלים.
התפתחות עקיפה - (זחל) עם מטמורפוזה - אופי. ד / חוסר עקביות, סרטנים, חרקים, תולעים, אופי. תקופה עוברית קצרה במהלך החתול. הזחל נוצר. עצמו אלא להתקיים. טרנספורמציה לא מלאה - ביצה-זחל-אימאגו. טרנספורמציה מלאה - ביצה - זחל - גולם - אימאגו. העובר מוקדם:הזרע מוכנס לתא הביצית, הציטופלזמה מתמוססת - להקת הסינקריון (הגרעינים ממזגים את הזיגוטה ה"דיפלואידית" 2n-div. על בלסטומרים-4, 8,16,32,64,128 ..-arr. blastula (עובר עם חלל בפנים) - על ידי אינוואגינציה arr. - דו שכבתי - גסטרולה, ect.ent) - (blastocoel, blastopore) - התפתח. 3 נבט. ליסטוק. אנדודרם, מזודרם, אקטודרם
ריסוק יכול להיות אחיד ולא אחיד (discoidal)
קיבה מתרחשת ב-4 דרכים:
1 על ידי אינטוסוסספציה (אינוואגינציה)
2 אפיבולי (עיבוי)
3 דלמינציה (ריבוד)
4 עלייה
זווית נוירולציה של הצינור העצבי, המיתר, קפלי מי השפיר והגזע
ectoderm יוצר עצב צלחת-st. neurula (הנחת צינור העצב, אקורדים), שמיעה, ריח, אינטגמנט חיצוני. Endoderm הוא ההתחלה של המעיים, הכבד, הריאות, בלוטת הלבלב. צורת מזודרם. שרירים (דרמוטום, מיוטום), רקמת שרירים, מערכת הדם, כל האלמנטים הסחוסיים והעצם של השלד, הדגשה, מערכת מינית.
16.נתיבים עיקריים של פילוגניה. סטייה, התכנסות, מקבילות.(52)
התפתחות אבולוציונית יכולה ללכת דרכים שונותתלוי באובייקט וביחס שלו עם הסביבה. ישנן מספר צורות של פילוגנזה.
אבולוציה פילטית היא סידור מחדש אבולוציוני הדרגתי של קבוצה טקסונומית מסוימת, מה שמוביל להופעתה של קבוצה חדשה השונה מהמקורית.דוגמה לאבולוציה פילטית יכולה להיות מספר אבות קדמונים של סוסים: eogippus - epigippus - merik-hipus - gashogippus - סוס מודרני.
הִסתַעֲפוּת - הדרך העיקרית של האבולוציה,שהיה מוטבע בבסיס התיאוריה הדרוויניסטית. אבולוציה מתפצלת נקבעת על ידי התפצלות של טקסון למספר קבוצות בהשפעת גורמים יסודיים של אבולוציה והסתגלות ל תנאים שוניםקִיוּם.עם סטייה, הדמיון נקבע על ידי מוצא משותף, והבדלים נקבעים על ידי הסתגלות לסביבה אחרת. תחילת ההתבדלות בתוך מין במהלך היווצרותם של אקוטייפים, גזעים אקולוגיים, תת-מינים גיאוגרפיים. כאשר הדמויות מתפצלות, הזנים הופכים למינים, המינים עוברים התמיינות נוספת ויוצרים סוגים, סוגים הופכים למשפחות, וכן הלאה.
סטייה יכולה ללבוש מספר צורות: הסתעפות דיכוטומית (פירוק של מין לשני מיני ילדים), קרינה (גן עדן נופל, של מין למספר מיני ילדים), התפצלות (ישות אבות קיימת בו-זמנית עם חברות בנות, שנוצרות מבודדות ב זמן שונהאוכלוסיות).סטייה מתפתחת על בסיס איברים הומולוגיים. דוגמה קלאסיתניתן לקרוא לסטייה הפער בין דמויות בחוחיות של ארכיפלג גלפגוס.
המשמעות הביולוגית של ההתבדלות נעוצה בעובדה שהיא מובילה למגוון של אורגניזמים חיים, לפיזור אחיד של מינים על פני גומחות אקולוגיות, ומחליקה את המתח של מאבק הקיום.
התכנסות היא תהליך הפוך של סטייה. הוא מורכב מרכישה עצמאית, באותם תנאים, של דמויות דומות בצורות שבעצם אינן קשורות על בסיס איברים או מבנים דומים.למשל, איבריו הנקברים של דוב השומה דומים; לוונים ודגים יש את אותה צורת גוף. ההתכנסות קובעת את הדמיון הכללי בין עופות שכשוך "קרסוליים" (עגורים ואנפות). תופעות התכנסות כוללות "מקבילים" גיאוגרפיים (אקלימיים), המתבטאים במספר גדול יותר של בעלי חיים בעלי דם חם של מדינות קרות בהשוואה לנציגים הקרובים ביותר של אזורים ממוזגים, במספר גדול יותר של מצמדים בציפורים ולשלשת ביונקים של הארקטי, בפיגמנטציה מוגברת של ציפורים טרופיות.
ההתכנסות בקבוצות מרוחקות של בעלי חיים נובעת מכך תנאים חיצונייםקרובי משפחה חייבים גם לקחת בחשבון קרבה. בהקשר זה, נצפית התכנסות בתוך הכיתה.
מַקבִּילוּת.אם קבוצות שונות שנפרדו בעבר נופלות בתנאי קיום שונים, הן יכולות באופן עצמאי, במקביל, ליצור תכונות הסתגלות דומות.לדוגמה, בדגי ים עמוק, איברים של זוהר התעוררו במקביל ובאופן עצמאי; התפתחות של שיניים חרבות בחתולים שונים לפעמים מקבילות מכסות מערכות איברים שלמות או חלק מהארגון כולו. לדוגמה, התפתחות מקבילה של דגים דו-חיים נושמים. סביר להניח שהדמיון בין דו-נושמים ודו-חיים נקבע על פי מערכת היחסים של קבוצות אלה. משמעותיות במיוחד הן ההקבלות במבנה הלב ו מערכת דם. ככל שיש יותר קשרים בין אורגניזמים, כך התפתחות מקבילה מכסה לעתים קרובות יותר חלקים משמעותיים של הארגון כולו. עדות לכך גם נתונים פליאונטולוגיים, למשל, התפתחות של חרטום. ככל שאורגניזמים קרובים יותר זה לזה, ככל שתגובותיהם לאותו שינוי בסביבה דומות יותר, כך השינויים במבנה שלהם דומים יותר (מקבילים). עבור התפתחות מקבילה, הן הסביבה החיצונית והן הגורמים הפנימיים (כלומר, המבנים התורשתיים המבוססים היסטורית של האורגניזם) חשובים. יַחַס גורמים פנימייםעם חיצוני קובע את האבולוציה של אורגניזמים. לדוגמה, פרסות שחיים באזורי אקלים שונים בורחים מפני טורפים בטיסה. לכן, אצל פרסות, נצפית התפתחות של דרגות קיצוניות של בהונות ולאחר מכן הפחתה הדרגתית של אצבעות הרגליים השוליות.
בסיסים גנטיים של אבולוציה מקבילה.עם שינוי מסוים בגורמים סביבתיים, אורגניזמים קשורים משתנים באופן דומה. במאבק על הקיום באמצעות הברירה הטבעית, השינויים הללו לובשים צורה "תורשתית". החומר לבחירה הוא סטיות בודדות במבנה, כלומר מוטציות. צורות קשורות או קרובות, ככלל, נותנות את אותן מוטציות (לדוגמה, סוגים שונים של תסיסנית). עם זאת, אין זה אומר שמוטציות מקבילות מבוססות על אותם שינויים באותם גנים.
ידוע שגנים שונים של אותו אורגניזם מייצרים מוטציות דומות. במיוחד אם השינויים הללו הם מינוריים. ומכיוון שרק שילובים של מוטציות קטנות נבחרות במהלך האבולוציה, ניתן לבנות שינויים מקבילים על בסיס גנטי שונה.
עבודת קורס
בביולוגיה
"מקורו של האיקריוט
הושלם על ידי: Severova Vera
כיתה י', בית ספר N55
נבדק על ידי: Strukova N.A.
ריאזאן 1999
1 . תיאוריה סימביוטית:
1.1 הבחנה בסיסית בין פרוקריוטים לאאוקריוטים;
1.2 אורגניזמים פרוקריוטים;
1.3 תאים אוקריוטיים;
1.4 מיטוכונדריה ופלסידים;
1.5 יכולתם של איקריוטים לבצע פוטוסינתזה;
2. מבשרי חיים;
3. אורגניזמים מתסיסים:
3.1 מקורות אנרגיה של תאים מוקדמים;
3.2 מתנוגנזה כמקור אנרגיה;
3.3 ציאנובקטריה וכלורווקסיבקטריה;
4. פוטוסינתזה ואוויר;
5. אסוציאציות ואיקריוטים:
5.1 תיאוריות של סימביוזות עוקבות;
5.2 רכישה סימביוטית של ניידות;
5.3 איקריוטים - פוטוסינתזה;
6. מבט על התפתחות התא:
6.1 השערה של השתייכות ישירה;
6.2 השערה: מציאנובקטריה ועד אצות;
6.3 תאימות של 2 השערות: השתייכות ישירה ו
אנדוזימביוזה;
7. מסקנות;
8. ספרות.
התיאוריה של אנדוזימביוזה
כל היצורים החיים על פני כדור הארץ מחולקים בדרך כלל לפרוקריוטים ואיקריוטים. תכונה עיקריתפרוקריוטים, בניגוד לאאוקריוטים, הם היעדר שלם גרעין התאמכוסה בקרום. תאים פרוקריוטים הם קטנים מאוד בגודלם, בסדר גודל של 1 מיקרומטר. כרך תאים איקריוטייםמכיל גרעין מלא, פי 800 - 1000 מהנפח של תאים פרוקריוטים. בהקשר זה, תאים - אאוקריוטים זקוקים למנגנון ממברנה גדול בהרבה כדי לבצע פונקציות חיוניות. ואכן, בתא איקריוטי זוהו מבנים תת-תאיים המוקפים בממברנות בלתי תלויות בפלזמה ובזה. מבנים אלו נקראים אברונים. לכל אברון: מיטוכונדריה, כלורופלסט, גרעין, ליזוזומים, מנגנון גולגי וכו' יש תפקידים ספציפיים משלו בחיי התא.
בעזרת גישות מולקולריות – ביולוגיות הושגו עדויות משכנעות שיש לחלק את כל היצורים החיים ל-3 ממלכות: חיידקים, ארכאים ואיקריוטים. מקורם של חיידקים וארכיאה על פני כדור הארץ מאב קדמון משותף - מה שנקרא פרוגנטו - לפני כ-4 מיליארד שנים, ולפי פליאונטולוגים, איקריוטים הופיעו על פני כדור הארץ 500 מיליון שנה לאחר הפרוקריוטים. הרעיון הפופולרי והמבוסס ביותר על מקורם של תאים אוקריוטים הוא כיום הרעיון של אנדוזימביוזה מרובה של פרוקריוטים שונים. לפי המיקרוביולוגים הידועים א' קלואיבר וק' ואן ניאל, מהפרוקריוטים העתיקים התרחשה בחירת התאים עם התגובות הביוכימיות החסכוניות ביותר ולעתים קרובות הרב-פונקציונליות, שהפכה לבסיס. הדרכים החשובות ביותרחילוף חומרים.
התיאוריה הסימביוטית של מקורם ואבולוציה של תאים מבוססת על שני מושגים חדשים בביולוגיה.לפי הראשון מבין המושגים הללו, ההבחנה הבסיסית ביותר בטבע החי היא ההבחנה בין פרוקריוטים לאאוקריוטים, כלומר. בין חיידקים לאורגניזמים המורכבים מתאי עם גרעינים אמיתיים - פרוטיסטים, בעלי חיים, פטריות וצמחים. הרעיון השני הוא שחלקים מסוימים של תאים אוקריוטים מקורם באבולוציה של סימביוזות, כלומר. יצירת קשרים קבועים בין אורגניזמים סוגים שונים. ההנחה היא ששלוש מחלקות של אברונים - מיטוכונדריה, ריסים ופלסידים פוטוסינתטיים - מקורם בחיידקים חיים חופשיים, שכתוצאה מסימביוזה נכללו ברצף מסוים בתאי מארחים פרוקריוטיים. תיאוריה זו מסתמכת במידה רבה על רעיונות ניאו-דרוויניים שפותחו על ידי גנטיקאים, אקולוגים, ציטולוגים ומדענים אחרים שקישרו את הגנטיקה המנדליאנית עם הרעיון הדרוויני. ברירה טבעיתהוא גם מסתמך על תחומים מדעיים חדשים לחלוטין או שחדשו לאחרונה: על ביולוגיה מולקולרית, במיוחד על מבנה החלבונים ורצף חומצות האמינו, על מיקרופליאונטולוגיה, החוקרת את עקבות החיים המוקדמים ביותר על פני כדור הארץ, ואפילו על הפיזיקה והכימיה של האטמוספרה, מכיוון שמדעים אלה קשורים לגזים ממקור ביולוגי. ניתן לקבץ את כל האורגניזמים התאיים לחמש ממלכות: הממלכה הפרוקריוטית (Monera, הכוללת חיידקים) וארבע הממלכות האיקריוטיות (Protoctista, Animalia, Fungi, Plantae). פרוטוקטיסטים הם אורגניזמים אוקריוטיים שאינם בעלי חיים, פטריות או צמחים. הממלכה הפרוטוקטית כוללת אצות, פרוטוזואה, עובשי רפש ושאר אורגניזמים אוקריוטיים שמקורם לא ברור. פרוטיסטים מוגדרים בצורה מגבילה יותר כאאוקריוטים חד-תאיים. לפיכך, הממלכה פרוטוקטיסטה כוללת לא רק פרוטיסטים - אוקריוטים חד-תאיים, אלא גם את צאצאיהם הרב-תאיים הקרובים ביותר, כמו אדום ו אצות חומות וגם מיקרואורגניזמים רבים הדומים לפטריות, כגון כיטרידים. על פי התפיסה המסורתית של פיליון ישיר, אברוני תאים כגון מיטוכונדריה ופלסידים התעוררו באמצעות מידור של התא עצמו.
נוצרים מספר פרוקריוטים
איקריוט אחד
תאים הם מערכות הקשורות לממברנה, הקטנה ביותר מבין היחידות הביולוגיות המסוגלות להתרבות עצמית. אפילו התאים הקטנים ביותר מכילים גנים בצורת מולקולות DNA (אחת או יותר) ומנגנון לסינתזה של חלבונים, המורכב מכמה סוגים של RNA וחלבונים רבים. כל התאים מכילים ריבוזומים, גופים בקוטר של כ-0.02 מיקרומטר, המורכבים משלושה סוגי RNA לפחות וכחמישים חלבונים שונים. אורגניזמים פרוקריוטיים הם היחידות הביולוגיות הקטנות ביותר העונות על הגדרה זו של תא. אין להם גרעינים. מצד שני, תאים אוקריוטיים בודדים (בעלי גרעין) הם, במובן מסוים, כלל לא יחידות. אלו הם קומפלקסים של יחידות שונות של סינתזה של חלבון (נוקלאוציטופלזמה, מיטוכונדריה ופלסטידים), שסוגם ומספרם משתנה בהתאם למין. לפיכך, לפי תיאוריה זו, תאים איקריוטיים התעוררו כתוצאה משיתוף פעולה של אלמנטים עצמאיים בתחילה, המאוחדים ב סדר מסוים. טיילור כינה את הרעיון הזה של המקור וההתפתחות הנפרדים של חלקים של תא אוקריוטי ואיחודם לאחר מכן בתור התיאוריה של אנדוסימביוזות עוקבות. תנאים טמפרטורה גבוההוחומציות הביאה לייצור בנוקלוציטופלזמה של מחלקה מיוחדת של חלבונים - היסטונים - כדי להגן על ה-DNA שלו. מיטוכונדריה הצליחו לפרק תרכובות שלושה פחמן לפחמן דו חמצני ומים. הקשר הסימביוטי של נוקלוציטופלזמה ומיטוכונדריה החל להשתנות בכיוון של חיזוק התלות ההדדית ביניהם. קומפלקס זה כלל אברונים מוטוריים - ריסים ודגלים. טרנספורמציה זו גררה טרנספורמציה מורפוגנטית, שכללה התפתחות של אסימטריות תאיות בולטות. עם שילוב של נוקלוציטופלזמה תלויה הדדית, מיטוכונדריה, אונדוליפודיה, הושלמה יצירת מבנה פוליגנומי של תא אוקריוטי. התא האיקריוטי הראשון היה הטרוטרופי. היא אכלה תרכובות אורגניות מוכנות. באוקריוטים אלה התעוררו תהליכי המיטוזה, ולאחר מכן המיוזה. ולבסוף, רכישת יכולת הפוטוסינתזה על ידי איקריוטים התרחשה בתהליך הפיכתו לארגון האיקריוטי הראשי ולאחר השלמתו - האירוע האחרון באפוס הזה. הסימביוזות הלכו והשתלבו. נכון לעכשיו, התלות של כל אברון בתוצרים המטבוליים של אברונים אחרים הפכה כה מלאה עד שרק שיטות מודרניותניתוח מאפשר לך להתחקות אחר המסלולים המטבוליים של השותפים המקוריים. אוקריוטים מגוונים במבנה שלהם, אך מבחינה מטבולית הם נשארים אחידים. הטריקים המטבוליים של איקריוטים פותחו חיידקים שוניםעוד לפני הזמן שבו התאחדו, והפכו למארח ולאברוניו. פרוקריוטים השיגו גיוון רב באופן שבו הם משיגים אנרגיה ובמורכבות הביוסינתזה. הם השאירו זכר לקיומם הרבה לפני שנוצרו אורגניזמים גדולים כל כך כמו חיות עם שלד מהתא האיקריוטי.
Pro Schemeו התכנסות של תאים אוקריוטיים על ידי סימביוזה
מבשרי חיים
אבותיהם של האורגניזמים של ימינו צמחו לפני יותר משלושה מיליארד שנים מתרכובות אורגניות שנוצרו על פני כדור הארץ. הערעור של מושג זה נובע בחלקו מיכולת הבדיקה שלו.
ס' מילר וה' אוריי הראו כי בתנאים האטמוספריים שהתרחשו על כדור הארץ הצעיר, יכולות להיווצר חומצות אמינו ומולקולות אורגניות אחרות. סוגים שוניםמולקולות אורגניות נוצרו באופן ספונטני במהלך מיליארד השנים הראשונות לקיום כדור הארץ. מולקולות אורגניות רבות נמצאו בחלל הבין-כוכבי ובמטאוריטים. אומר, תרכובות אורגניותלהתעורר במהלך האינטראקציה של אלמנטים קלים, בנוכחות מקורות אנרגיה. רבגוניות מנגנון כימירבייה ביולוגית פירושה שכל החיים על פני כדור הארץ מגיעים מאבות קדמונים משותפים: כל החיים הנוכחיים מקורם בתאים עם מערכות מידע המבוססות על שכפול DNA ועל סינתזת חלבון המכוונת על ידי RNA שליח.
אורגניזמים מתסיסים
ייתכן שחומצות גרעין הופרדו מהסביבה כבר מההתחלה על ידי ממברנות ליפופרוטאין. מידע החל להיות מקודד על ידי רצף הנוקלאוטידים במולקולות ה-DNA. התאים העתיקים ביותר השתמשו ב-ATP ובחומצות אמינו, שהן חלק מחלבונים, כמקורות ישירים לאנרגיה וכפי רכיבים מבניים. אז - החיים העתיקים ביותר היו קיימים בצורה של חיידקים הטרוטרופיים שקיבלו מזון ואנרגיה מחומר אורגני ממקור אביוטי. אבולוציה אורגנית ניאו-דרוויניסטית דורשת שלוש תופעות: רבייה, מוטציה ולחץ סביבתי סלקטיבי. רבייה היא מצב בלתי ישים של אבולוציה ביולוגית. עם רבייה מדויקת מספיק, מוטציות וברירה טבעית הן בלתי נמנעות. להסביר את מקורה של רבייה כזו היא להבין את מקור החיים עצמם. מוטציה, המקור העיקרי לשונות באבולוציה, מובילה לשינויים תורשתיים במערכת רבייה עצמית. לאחר שפותחה רבייה אמינה, החלה האבולוציה הניאו-דרוויניסטית. אורגניזמים שפיתחו מסלולים סינתטיים כלוב נחוץלרכיבים, היה יתרון על פני אחרים. האוטוטרופים העתיקים ביותר, כלומר, אורגניזמים המסוגלים לספק את צרכיהם לאנרגיה ו חומר אורגניאה, בגלל מקורות אנאורגניים, היו חיידקים אנאירוביים המייצרים מתאן. יכולת הפוטוסינתזה פותחה לראשונה בחיידקים הרגישים לחמצן. מְרַתֵק אוֹר שֶׁמֶשׁבעזרת כלורופיל חיידקי, הם המירו פחמן דו חמצני אטמוספרי לחומרים אורגניים.הופעת הפוטוסינתזה האנאירובית שינתה באופן בלתי הפיך את פני השטח והאטמוספירה של כדור הארץ.
פחמן דו חמצני הוצא מהאטמוספרה והפך לחומר אורגני בהתאם למשוואת הפוטוסינתזה הכללית: 2H2X+CO2=CH2O+2X+H2O. חיידקים פוטוסינתטיים שהשתמשו במימן גופרתי להפחתת CO2 הפקידו גופרית יסודית. הם הולידו חיידקים המסוגלים לבצע פוטוסינתזה עם שחרור חמצן. סוג זה של פוטוסינתזה מקורו כנראה כתוצאה מהצטברות של מוטציות בחיידקי גופרית פוטוסינתטיים. נתיב זה הוביל להופעתם של אצות כחולות ירוקות או ציאנובקטריה. גילה ולמד לאחרונה קבוצה חדשהפרוקריוטים פוטוסינתטיים המייצרים O2 - chloroxybacteria. ניתן לשער שדווקא ציאנו- וכלורווקסיבקטריה היו אבותיהם של הפלסטידים המצויים בתאי האצות והצמחים. כאשר O2 הצטבר כתוצאה מפוטוסינתזה מיקרוביאלית, אטמוספירת כדור הארץ הפכה להתחמצנות, מה שגרם למשבר של כל החיים - עד לאותה תקופה העולם היה מיושב בחיידקים אנאירוביים. כדי לשרוד, החיידקים נאלצו להימנע מחמצן על ידי הסתתרות בביצות, או שהם היו צריכים לפתח חילוף חומרים המסוגל להגן עליהם מפני כמות החמצן הגוברת.
פוטוסינתזה ואוויר
לפני כ-2 מיליארד שנים, בזה אחר זה, התרחשו תהליכים אבולוציוניים שונים ליצירת מגוון מטבולי. את היכולת לייצר אנזימים ספציפיים ניתן להעביר בתוך אותו מין באמצעות חלקיקים דמויי וירוס הנקראים פלסמידים. היכולות הביוסינתטיות של פרוקריוטות בולטות: מפחמן דו חמצני, מספר מלחים ותרכובות אנאורגניות מופחתות המשמשות כמקורות אנרגיה, מיקרואורגניזמים כימואוטוטרופיים יכולים לסנתז במים לא מוארים המכילים חמצן את כל המקרומולקולות המורכבות ביותר הנחוצות לצמיחה ורבייה. תהליכים ביוכימיים אלו הובילו למערכות של מחזור היסודות גם כאשר לא היו צמחים, פטריות, בעלי חיים.
אסוציאציות ואיקריוטים.
מבט על התפתחות התא.
סניף ישיר.
ההשקפה המסורתית לגבי מקורם והתפתחותם של תאים אוקריוטיים היא שכל היצורים החיים צאצאו ישירות מאוכלוסיית אבות אחת כתוצאה מהצטברות של מוטציות בהשפעת הברירה הטבעית.
מוטציות נקודתיות, כפילויות, מחיקות ושינויים תורשתיים אחרים, שידוע כי מילאו תפקיד באבולוציה של בעלי חיים וצמחים, אחראים גם להתמיינות של תאים אוקריוטיים מתאים פרוקריוטיים - תורת הפיליון הישירה. כלומר, הרעיון של מקור לא סימביוטי של תאים. אולי אצות ירוקות הן צאצאים של אבותיהם של כל הצמחים. תכונות רבות של אצות אלה מעידות על כך שיש להן ולצמחים אב קדמון משותף. הפילוגניה של בעלי חיים מורכבת יותר, אבל המושג של השתייכות ישירה חל כנראה גם על הנתיבים האבולוציוניים שלהם. ספוגים ככל הנראה התפתחו מפרוטיסטים, ללא תלות בקבוצות אחרות של בעלי חיים. אז, לפחות עבור בעלי חיים וצמחים, ההשערה שאורגניזמים אוקריוטיים מורכבים התפתחו מאיקריוטים מסודרים יותר באמצעות הצטברות של מוטציות בודדות בהשפעת הברירה הטבעית היא לגיטימית למדי. חידושים אבולוציוניים מסתמכים על שימור של גנים חיוביים מושלמים ביותר. אז, אורגניזמים חדשים בעלי יכולת מוגברת לשלוט בסביבה שלהם התפתחו יותר על ידי שמירה על שכפול ושילוב מחדש של גנים מותאמים מאוד מאשר על ידי רכישת תכונות חדשות באמצעות מוטציות נקודתיות פשוטות ואקראיות. ההשערה של השתייכות ישירה אינה מסבירה את מקורם של איקריוטים מפרוקריוטים. לדוגמא, קשה להבין את ההפסקה בין ציאנובקטריות שאינן מסוגלות למיטוזה לבין צורות כאלה עם מיטוזה מפותחת כמו אצות אדומות מנקודת מבט של השתייכות ישירה, אך היא נובעת באופן טבעי מהתיאוריה הסימביוטית.
על פי תורת ההשתייכות הישירה ותיאוריית הסימביוזה, כל האורגניזמים על פני כדור הארץ מקורם באבות חיידקים דומים מבחינה ביוכימית, וחיידקים אוקריוטיים הולידו צמחים ובעלי חיים. חיידקים פוטוסינתטיים הולידו אצות, ובסופו של דבר צמחים, וכמה אצות איבדו את הפלסטידים שלהם והפכו לאבותיהם הקדמונים של פטריות ובעלי חיים. תיאורטיקנים של פיליה ישירה מסכימים שפלסטידים ואברונים אוקריוטיים אחרים, כולל הגרעין, נוצרו על ידי התמיינות בתוך התאים עצמם ומכנים זאת מיתוס בוטני.
פילוגניה של סוגי צמחים
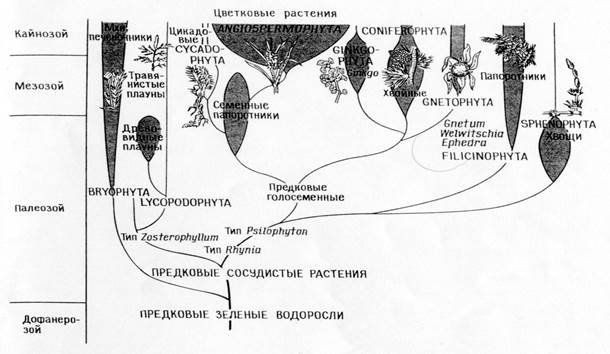
השוואה של פילוגניות התואמות לתיאוריית ההשתייכות הישירה (A)
ותאוריית הסימביוזה (ב)
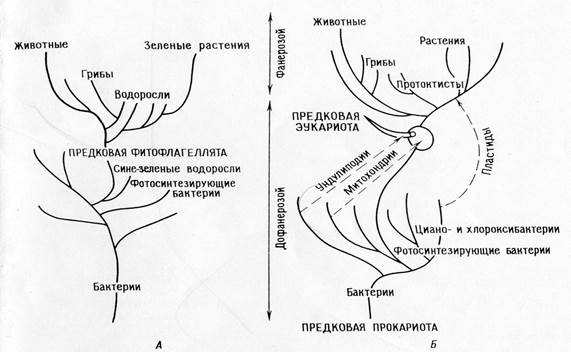
הבדלים בין תיאוריית ההשתייכות הישירה לתיאוריה הסימביוטית:
התיאוריה של השתייכות ישירה.
דיכוטומיה עיקרית: בעלי חיים - צמחים
איקריוטים פוטוסינתטיים (אצות וצמחים ירוקים) התפתחו מפרוקריוטים פוטוסינתטיים (אצות כחולות-ירקות)
צמחים ומערכות הפוטוסינתזה שלהם התפתחו באופן מונופילטי בפרקמבריון.
בעלי חיים ופטריות התפתחו מאוטטרופים שאיבדו פלסטידים.
המיטוכונדריה התמיינה תוך תאית באצות פוטוסינתטיות.
בעשבים הימניים, האונדוליפודיה, המנגנון המיטוטי וכל שאר האברונים האוקריוטיים התמיינו תוך תאי.
כל האורגניזמים צאצאו מאבותיהם - פרוקריוטים על ידי הצטברות של מוטציות בודדות.
תוצאה: על פי התיאוריה, חייבים להיות עשבים נכונים; קשה לקשר את התיאוריה הזו לתיעוד המאובנים ולבנות פילוגניה מתאימה.
התיאוריה של אנדוסימביוזות עוקבות:
הדיכוטומיה העיקרית: פרוקריוטים - איקריוטים.
איקריוטים פוטוסינתטיים (אצות וצמחים גרעיניים) ואיקריוטים שאינם פוטוסינתטיים (בעלי חיים, פטריות, פרוטוזואה) התפתחו מאבות קדמונים הטרוטרופיים נפוצים (אמבופלגלטים).
האבולוציה של הפוטוסינתזה בארכאים הייתה מונופילטית בחיידקים (כולל ציאנו-וכלורווקסיבקטריות), פרוטיסטים פוטוסינתטיים התעוררו לפתע כאשר הפרוטיסטים-אבותיהם ההטרוטרופיים נכנסו לסימביוזה עם פרוקריוטים פוטוסינתטיים (פרוטופלסטידים).
בעלי חיים ורוב האצות והפטריות הם צאצאים ישירות מפרוטיסטים הטרוטרופיים.
מיטוכונדריה כבר היו נוכחים בפרוטיסטים הטרוטרופיים כאשר האחרונים רכשו פלסטידים כתוצאה מסימביוזה. פרומיטוכונדריה נרכשו בעבר באופן סימביוטי.
מיטוזיס התפתחה בפרוטיסטים אוקריוטים הטרוטרופיים על ידי התמיינות של המערכת הבלתי-ליפודיאלית.
פרוקריוטים התפתחו מאבות פרוקריוטים מוקדמים דרך סדרה של אירועים מוטציוניים בודדים, איקריוטים התפתחו גם מאבות אוקריוטים דרך אירועים מוטציוניים בודדים, ואבות איקריוטים התפתחו מפרוקריוטים דרך סדרה של סימביוזות.
השלכות: התיאוריה מסבירה מסלולים ביוכימיים ואת יתירותם באברונים. זה מאפשר בניית פילוגניה עקבית שמסבירה את "הפער הביולוגי" בין הזמנים הפרוטרוזואיקונים והפנרוזואיקונים. התיאוריה חוזה שאברונים חייבים לשמור על שרידים של מערכות שכפול.
מיתוס בוטני
מתקופתו של ג'וסייה צמחים נמוכים יותרהנקראים טלופיטים.וגבוהים יותר - בריאופיטים וטרכאופיטים. על פי המיתוס הבוטני, האבות הקדמונים המשותפים של חיידקים פוטוסינתטיים ואצות כחולות-ירוקות, באמצעות שיוך ישיר, הולידו אצות אוקריוטיות, כולל אצות ימיות. קבוצות של אורגניזמים שהפילוגניה שלהם מושפעת יותר מכל מהמושג של פיליון ישיר הגיעו בסופו של דבר לתחום הבוטניקה - אלו הם חיידקים, אצות, פטריות וצמחים.
אורגניזמים היפותטיים שיכולים להוות קישור בין ציאנובקטריה לאצות ירוקות נקראו פיטומונאדים קדומים (פיטומונדים קדומים) או עשבים ימניים. כמה מחברים טענו שאאוקריוטים Cyanidium ו- Cyanophora, הדומים במערכות הפיגמנט שלהם לציאנובקטריה, עשויים להיות מועמדים מתאימים לתפקיד של עשבים ימניים. בטלופיטים לא נמצאו צורות ביניים בין המצב הלא-מיטוטי של ציאנובקטריה לבין המיטוזה האופיינית המפותחת במלואה של אצות ירוקות וצמחים. למרות פטריות ו צמחים פורחיםנטולי undulipodia לחלוטין, הם מאופיינים במיטוזה ומיוזה, הדורשים צירים המורכבים ממיקרוטובולים. ישנה הסכמה כללית שכל האורגניזמים הרב-תאיים עם מיקרו-צינוריות הם צאצאים ממיקרואורגניזמים אוקריוטיים עם אונדוליפודיה, אך איש אינו יודע מאיזו קבוצה של פרוטיסטים. חיזק את המיתוס הבוטני היה העתיקות והמורכבות של הפוטוסינתזה, כמו גם העובדה שלאורגניזמים רבים (חיידקים, אוגלנואידים, צמחים טפילים), הן בטבע והן במעבדה, היה אובדן פוטוסינתזה כתוצאה ממוטציות. כיוון שפוטוסינתזה היא תהליך אנאירובי המבטיח בסופו של דבר את קיומם של כל האורגניזמים, היא בוודאי התפתחה בשלב מוקדם מאוד בהיסטוריה של החיים על הפלנטה שלנו, עוד לפני שהתעוררה מיטוזה הדורשת חמצן. פיגמנטים, ליפידים ואנזימים רבים מעורבים בפוטוסינתזה, מסודרים בצורה מסודרת בממברנות מורכבות מאוד. יחד עם זאת, הפרטים של מנגנוני פוטוסינתזה באצות ובצמחים מחד ובציאנובקטריה מאידך דומים להפליא. כל האורגניזמים הללו משחררים גז חמצן מהמים, המשמש כמקור לאטומי מימן המשמשים להפחתת CO2 לחומר האורגני של התא. לאצות ולפרוטוזואים רבים אין לא מיטוזה ולא מערכת גנטית דומה לזו של בעלי חיים וצמחים. לסילאטים מערכת גנטית מיוחדת עם 2 סוגי גרעינים, ולרוב התאים הם מרובי גרעינים. סוג אחד של גרעינים הוא מקרו-גרעינים, והשני הוא מיקרו-גרעינים קטנים בהרבה. מיקרו-גרעינים הם חסרי משמעות פיזיולוגית. עם זאת, הם מאחסנים עותקים של גנים, ובמהלך התהליך המיני, הם עוברים מיוזה. גרעיני מקרו חיוניים מבחינה פיזיולוגית משמשים כאתר של שעתוק RNA ובכך שולטים בפיזיולוגיה של התא. אחד הקשיים בבירור הקשר בין ציאנובקטריה לאצות אוקריוטיות הוא המחסור בנתונים על מקורה של אונדוליפודיה: לא נמצאו קשרי ביניים בין ציאנובקטריה, שאף פעם לא קיימת בה, לבין האצות הרבות שיש. בעיה נוספת היא היעדרות מוחלטתתהליך מיני בציאנובקטריה: בין ציאנובקטריה א-מינית לאצות, שרובם מאופיינים בתהליך מיני, לא היו צורות ביניים. נמצא שלרוב האצות יש אונדוליפודיה בשלב מסוים מעגל החיים. התברר שהמבנים הניידים באצות, פרוטוזואה, בעלי חיים וצמחים זהים: כולם מורכבים ממיקרו-צינוריות בקוטר של 0.024 מיקרון. התברר שהציר המיטוטי, גם בפטריות שאין להן אונדוליפדיה, מורכב ממיקרוטובולים חלבוניים בקוטר זהה לזה של אונדוליפדיה. במשך חצי מאה, זה נראה סביר ביותר שאצות אוקריוטיות התפתחו מציאנובקטריה. המיתוס הבוטני הזה הוביל להנחות שבמשך שנים רבות שימשו הן כגירוי למחקר והן מערכת קואורדינטות, באשר בוצעה הערכת התוצאות שהתקבלו. הקביעה שהקשרים בין ציאנובקטריה לאצות איקריוטיות גוועו, ולא הותירו עקבות מאובנים ושרידים חיים, הפכה פחות ופחות משכנעת ככל שנתונים חדשים על חיידקים ומאובנים חיים. קשרי הביניים העיקריים בין פרוקריוטים פוטוסינתטיים לאאוקריוטים לא גוועו בלי זכר, כמתחייב מהמיתוס הבוטני - הם מעולם לא היו קיימים. לפי התיאוריה של אנדוסימביוזות עוקבות, ציאנובקטריות הן אכן האבות הקדמונים, אבל רק פלסטידים, ולא שאר התא האיקריוטי, ובוודאי לא הגרעין שלו. ובכל זאת, בשאלת מקורו של הגרעין, השערת ההשתייכות הישירה ותורת הסימביוזה תואמות. שינויים אפשריים לגרסה הקיצונית של תיאוריית הסימביוזה כוללים, למשל, את הרעיון של מקור סימביוטי של פלסטידים, אך לא מיטוכונדריה, או של מקור סימביוטי של פלסטידים ומיטוכונדריה, אך לא אונדוליפודיה. תומכי ההשתייכות הישירה הציגו גם הנחה לגבי מקורו של הגרעין באמצעות סימביוזה.
עדות קריטית למקור האנדוזימביוטי של כלורופלסטים ומיטוכונדריה.
העדות המשמעותית ביותר למקורם של האברונים החשובים ביותר של תאים אוקריוטים - כלורופלסטים ומיטוכונדריה מפרוקריוטים התקבלה ב השנים האחרונותכאשר לומדים את המבנה של RNA ריבוזומלי. ביולוג מולקולרי
K. Woz הראה כבר בשנות ה-80 שהמבנה של RNA מהריבוזומים של כלורופלסטים צמחיים כמעט זהה למבנה של RNA מכמה ציאנובקטריות. במקביל, קבוצת חוקרים מצאה שה-RNA של ריבוזומים שמקורו במיטוכונדריה של אוקריוטים שונים דומה ל-RNA של כמה חיידקים, בפרט של חיידקים מהסוג Paracoccus. סט אנזימי הנשימה של חיידק זה דומה מאוד ל
"אנסמבל נשימתי" של מיטוכונדריה של בעלי חיים.
נתונים אלה תומכים בבירור ב יותר סבירמקור אנדוזימביוטי של תאים אוקריוטיים. תופעת האנדוזימביוזה עדיין נפוצה ביותר בטבע. זה ידוע כי על הגוף של כמה דגים ימייםלעתים קרובות מתיישבים חיידקים זוהרים, אשר לא רק מקבלים יתרונות גדולים עבור חייהם, אלא גם שימושיים מאוד עבור אורגניזמים מארח. כל האמור לעיל מעיד באופן משכנע בעד המקור האנדוסימביוטי של תאים אוקריוטיים מתאי פרוקריוטים קטנים בהרבה. עם זאת, השאלה העיקרית היא לקבוע את אופיו של התא המארח, מכיוון שלתאים אוקריוטים יש גרעין בנוי היטב עם קליפה, בעוד לפרוקריוטים אין גרעין כזה. ל' מרגליס, למשל, סבור שתפקידם של תאי מארח יכול להיות שמילאו פרוקריוטים - קודמיו חיידקים מודרנייםקשור ל-mycoplasmas - פרוקריוטים הבנויים בצורה מאוד פרימיטיבית, כמעט נטולי דפנות תאים. IN לָאַחֲרוֹנָהעבודותיו של המדען היפני הופיעו ונדונות רבות
ט אושימה. בהתבסס על מחקר יסודי של המבנה של RNAs ריבוזומליים של פרו-אוקריוטים רבים, כולל נציגים רבים של ממלכת העל של הארכאה, שנחקרה מעט עד כה, מסיק מחבר זה שהתא המארח היה אחד מאבותיה הקדמונים של הארכאה המודרנית. יש לומר כי לנציגי ממלכת העל הזו, בניגוד לחיידקים, יש הרבה תכונות ביולוגיות ביוכימיות ומולקולריות במשותף לאאוקריוטים. בעבודותיו האחרונות של T. Oshima, ניתנים טיעונים בעד העובדה שהתא המארח בזמן הופעת האאוקריוטים היה האב הקדמון של הארכאה, בדומה לאחד הנציגים המודרניים של הפרוקריוטים הללו - תרמופלזמה. מה יהיה הפתרון הסופי לבעיית ביסוס טבעו של התא המארח שהוליד את התא האוקריוטי בתהליך של אנדוזימביוזה נפוצה יראה על ידי מחקרים נוספים.
כעת מוצגת ההשערה הפופולרית והמבוססת ביותר לגבי מקורם של תאים אוקריוטיים
אנדוזימביוזה של פרוקריוטים שונים השייכים לחיידקים ולארכיאה, שהתרחשה פעמים רבות בתהליך האבולוציה במהלך 1.5-2 מיליארד שנות קיומם הראשונות על פני כדור הארץ של תאים חיים. עדות נחרצת למקור האנדוזימביוטי של הכלורופלסטים והמיטוכונדריות התקבלה על ידי הביולוגים המובילים בעולם בחקר מבנה ה-RNA הריבוזומלי. נתונים אלה תומכים בבירור ב סבירות גבוההמקור אנדוזימביוטי של תאים אוקריוטיים מתאי פרוקריוטים קטנים בהרבה (במספר סדרי גודל).
סִפְרוּת
1. כתב עת חינוכי של סורוס. ביולוגיה, כימיה, גיאוגרפיה, פיזיקה, מתמטיקה. N 5 1998
2 Margelis L. תפקיד הסימביוזה באבולוציה של התא. לְכָל. מאנגלית, M. MIR 1983
3 יסודות המיקרוביולוגיה. מ' "רפואה" 1992.
4 Khakhina L.N. הקונספט של A.S. פמינסין על חשיבות הסימביוזה באבולוציה של ל.; מדע, 1981, עמ'. 165 - 181 .
5 Alberts B. ביולוגיה מולקולרית של התא; לְכָל. מאנגלית. M. Mir 1986 v.1
המונח Prokaryotae (קדם-גרעיני) מתייחס למיקרואורגניזמים בעלי ארגון פרימיטיבי של מבנים גרעיניים. השם הזה הוצע על ידי המדען הקנדי מורי ב-1968.
במחצית השנייה של המאה העשרים. התברר שכל היצורים החיים עם מבנה תאי, ניתן לשייך לאחת משתי קבוצות - פרוקריוטים או אוקריוטים. תאים פרוקריוטים הם פשוטים יותר - הם נטולי גרעין מוקף בקליפה. במשך זמן מה, האמינו שהמונחים "פרוקריוטים" ו"חיידקים" הם מילים נרדפות וכי חיידקים מייצגים ענף עצמאי ומאוחד של האבולוציה של יצורים חיים. עם זאת, הצטברו בהדרגה עדויות לכך שההבדלים בין חיידקים מסוימים יכולים להיות משמעותיים ולהצביע על כך שאין ביניהם קשר אבולוציוני הדוק. במיוחד חשיבות רבהמצורף לנתונים על המבנה של חומצות ריבו-נוקלאיות ריבונומיות ותחבורה (RNA). נכון לעכשיו, בהתבסס על תוצאות ההשוואה של רצפי בסיס במולקולות RNA ריבוזמליות, נשפט הקשר בין אורגניזמים. סוג זה של נתונים, כמו גם מאפיינים אחרים של פרוקריוטים, הראו שלמעשה, יש לחלק פרוקריוטים לפחות לשתי קבוצות בעלות מוצא שונה ונתיבים אבולוציוניים שונים. בנוסף, תאים איקריוטים לא רק מאורגנים בצורה מורכבת יותר, אלא גם מכילים אלמנטים שהם לא יכלו לקבל מאבותיהם הקדמונים של הפרוקריוטים המודרניים. אלו הם, קודם כל, ריבוזומים ציטופלזמיים, השונים מריבוזומים פרוקריוטיים. דמיון בסיסי קוד גנטי, הארגון של מקרומולקולות והמנגנון הביוכימי של סינתזת חלבון מצביע על אחדות המקור של כל האורגניזמים החיים. ההנחה היא שהיה אב קדמון משותף מסוים - "פרוגנוט", אבל מה הוא היה לא ידוע. פרוגנט זה יכול להוליד שלושה ענפים עצמאיים של העץ האבולוציוני. מנקודת מבט של שיטתיות היררכית, ענפים אלה מוצעים להיחשב כתחומים בעלי דרגה גבוהה יותר מממלכות מסורתיות. תחומים אלו (אימפריות) הם Eubacteria, Archaebacteria, Eukaryota (eubacteria, archaebacteria ו-eukaryotes). מאפיינים כלליים ומבדילים של ארכיבקטריות, אובקטריות ואוקריוטות ניתנות בטבלה.
המאפיין העיקרי של תא פרוקריוטי הוא היעדר גרעין, מוגבל מהציטופלזמה על ידי ממברנה כפולה. החומר התורשתי מרוכז בכרומוזום החיידקים, המיוצג בדרך כלל כמולקולת DNA דו-גדילית מעגלית. לחלק מהספירושטים יש כרומוזומים ליניאריים, בעוד שלאקטינומיצטים יש כרומוזומים "פסאודו-טבעות". מולקולת ה-DNA דמוי החוט פועלת כגרעין וממוקמת באזור המרכזי של התא, הנקרא "נוקלואיד". במיקרוסקופ אלקטרונים, על קטע של תא חיידקי, אזור זה נראה קל יותר משאר הציטופלזמה.
לתא הפרוקריוטי יש מספר מאפיינים בסיסיים הקשורים הן לאולטרה-מבנה שלו והן ארגון כימי. קודם כל, אין רשת ממברנות בתא החיידק - הרשת האנדופלזמית. בתאים פרוקריוטיים, הממברנה הציטופלזמית השוכנת מתחת לדופן התא ועוטפת את הפרוטופלסט מבחוץ היא הממברנה היחידה בתא.
הממברנה הציטופלזמית המקיפה כל תא קובעת את גודלו ומבטיחה שמירת הבדלים משמעותיים בין תכולת התא והסביבה. אבל הממברנה היא לא רק מחיצה מכנית. הוא משמש גם כמסנן סלקטיבי ביותר השומר על הבדל בריכוזי היונים משני צידי הממברנה ומאפשר לחומרים מזינים להיכנס לתא וליציאת תוצרי פסולת. הממברנה הציטופלזמית היא מכלול של מולקולות שומנים וחלבונים המוחזקים יחד על ידי אינטראקציות לא קוולנטיות. IN מיקרוסקופ אלקטרוניממברנות נראות כמו יריעות בעובי של כ-7 ננומטר עם מבנה תלת-שכבתי בולט. השכבה הכפולה של מולקולות השומנים קובעת את המאפיינים המבניים של הממברנה, בעוד שחלבונים אחראים לרוב תפקודי הממברנה.
המאפיינים העיקריים של archaebacteria, eubacteria ו-eukaryotes
| שלטים | archaebacteria | eubacteria | איקריוטים |
| אורגניזמים אופייניים | מתנוגנים, תרמופילים קיצוניים, הלופילים | קבוצות עיקריות של חיידקים | פרוטיסטים, פטריות, צמחים, בעלי חיים |
| מידות אופייניות, מיקרון | 0,5-4 | 0,5-4 | מעל 5 |
| גנום | כרומוזום טבעת | כרומוזום טבעת | גרעין עם כרומוזומים מורכבים רבים |
| מבני ממברנה: ליזוזומים מורכבים של רטיקולום אנדופלזמי Golgi | לא לא לא | לא לא לא | כן כן כן |
| מיטוכונדריה | לא | לא | יש |
| כלורופלסטים | אין (לפעמים thylakoids) | לא | לא (בפטריות ובעלי חיים), כן (בצמחים ואצות) |
| קירות תאים | חלבון, פסאודומוראין | murein, ליפופוליסכריד | שׁוֹנִים |
| ריבוזומים | 70S* | 70S | 80S (בציטופלזמה) + 705 ריבוזומים של אברונים, כמו בחיידקים |
| RNA פולימראז | מורכב | פָּשׁוּט | מורכב |
| פלאגלה | פָּשׁוּט | פשוט, מורכב מסיבי אחד או יותר | מבנה מורכב (2х9)+2 |
| קיבוע חנקן | יש | יש | לא |
| נשימה אנאירובית | יש | יש | לא |
| כימוסינתזה | יש | יש | לא |
S* - יחידת שקיעה של Svedberg המאפיינת את גודל הריבוזום
בממברנה הציטופלזמית של פרוקריוטות ישנן פלישות מרובות - פלישות המבצעות פונקציות שונות. הם עשויים להכיל אנזימים ולהיראות כמו שלפוחיות או שהם מזוזומים ומעורבים בחלוקת תאים. הממברנה הציטופלזמית של הפרוקריוטים היא אתר לוקליזציה של אנזימי חילוף החומרים באנרגיה, ולכן, מבצעת את הפונקציה של מיטוכונדריה, אשר נעדרות בתאים פרוקריוטים. לכן, חיידקים עם רמה גבוההמטבוליזם נשימתי, הממברנה הציטופלזמית מפותחת במיוחד, כלומר. יש לו פלישות רבות, מה שמגדיל את פני השטח שלו. הממברנה הציטופלזמית מעורבת גם בחלוקת הנוקלואיד. DNA מעגלי מקובע על הממברנה ולאחר שכפול (הכפלה) מתפצל לשני תאים חדשים עקב צמיחת הממברנה. כיסויי אנדוספור נוצרים מהממברנה הציטופלזמית. זהו גם המחסום העיקרי לחדירת חומרים לתא. הממברנה הציטופלזמית אינה מאפשרת למולקולות גדולות לעבור דרכה, היא אינה מסוגלת לפינוציטוזיס ואקסוציטוזיס.
מבנים הממוקמים מחוץ לממברנה הציטופלזמית - דופן התא, קפסולה, קרום רירי, דגלים, פימבריה, פילי, וילי - הם מבני פני השטח.
דופן התא היא מרכיב מבני חשוב ומחייב של רוב התאים הפרוקריוטים (חיידקים אמיתיים, לא סופרים ארכאיבקטריות ומיקופלזמות), הממוקמים מתחת לקפסולה או לקרום הרירי או, בהיעדר האחרון, במגע עם הסביבה. דופן התא משמשת מחסום מכני בין הפרוטופלסט לבין סביבה חיצוניתונותן לתא צורה מסוימת הטבועה בו.
מבחינת מבנה והרכב כימי, דופן התא של פרוקריוטים שונה באופן חד מזה של אורגניזמים אוקריוטיים. דופן התא של פרוקריוטים מורכבת מכמה שכבות, שהשכבה העיקרית - המסגרת - האחראית לחוזק, נוצרת מחומר מיוחד - murein peptidoglycan, המסונתז רק על ידי התא הפרוקריוטי ואינו נמצא באורגניזמים אחרים. בקבוצת הפרוקריוטים האמיתיים מבחינים בשני סוגים עיקריים של מבנה דופן התא. שני סוגים אלו תואמים לחיידקים גראם חיוביים וגרם שליליים. שמות הקבוצות הללו באים מהיכולת או חוסר היכולת חיידקים שוניםכתם לפי גרם. ב-1884 הציע הרופא הדני ק.גראם, שעסק בצביעת רקמות, שיטת צביעה, ששימשה מאוחר יותר לחיידקים. אם תאים קבועים של eubacteria מטופלים תחילה עם תמיסה של סגול קריסטל ולאחר מכן עם יוד, אז נוצר קומפלקס צבעוני. במהלך הטיפול הבא באלכוהול, בהתאם למבנה דופן התא, גורלו של הקומפלקס שונה: במינים שנקראים גרם חיוביים, הקומפלקס הזה נשמר על ידי התא והאחרונים נשארים צבעוניים, בגראם שלילי. מינים, הקומפלקס הצבעוני נשטף מהתא, והם הופכים לדהויים. נמצא כי הקומפלקס הצבעוני נוצר על הפרוטופלסט, אך החזקה שלו על ידי התא או שטיפה ממנו במהלך הטיפול הבא באלכוהול נקבעת על פי המאפיינים המבניים של דופן התא. נמצאה שיטת כתם גראם יישום רחבבתרגול מיקרוביולוגי להתמיינות של חיידקים.
דפנות התא של אובקטריות גראם חיוביות וגראם שליליות שונות באופן חד הן בהרכב הכימי והן במבנה האולטרה.
עיקר דופן התא של חיידקים גרם חיוביים הוא הטרופולימר ספציפי - peptidoglycan murein. עמוד השדרה הפוליסכריד של המולקולה בנוי משאריות מתחלפות של N-acetylglucosamine וחומצה N-acetylmuramic, המחוברים ביניהם בקשרים גליקוזידיים. בנוסף לפפטידוגליקן, דפנות התא של אובקטריות גרם חיוביות כוללות חומצות טייכואיות ופוליסכרידים. חומצות Teichoic הן פולימרים המבוססים על אלכוהולים רב-הידריים(ריביטול וגליצרול), מחוברים ביניהם בקשרי פוספודיסטר.
לחיידקים גראם-שליליים יש מבנה דופן תא מורכב הרבה יותר מאשר לגראם-חיובי. הוא מכיל הרבה יותרמקרומולקולות שונות סוג כימי. פפטידוגליקן יוצר רק את השכבה הפנימית של דופן התא, נצמד באופן רופף לממברנה הציטופלזמית. מחוץ לפפטידוגליקן נמצאת שכבה נוספת של דופן התא - קרום חיצוני, המורכב מפוספוליפידים (אופייני לממברנות אלמנטריות), חלבונים, ליפופרוטאין וליפופוליסכריד.
הבדלים שנמצאו אמפירית בין שתי קבוצות החיידקים ביחס לכתמי גראם נמצאו בקורלציה עם תכונות חשובות רבות אחרות. לדוגמה, חיידקים גרם חיוביים מתרבים בביקוע בינארי עקב היווצרות מחיצה רוחבית בתא, גרם שלילי - על ידי יצירת כיווץ. הראשונים רגישים לפניצילין, האחרונים לא. חיידקים גראם שליליים אינם יוצרים אנדוספורים, בעוד שחלק גראם חיוביים עושים זאת.
דופן התא של פרוקריוטות מבצעת מגוון פונקציות: היא מגינה על התא מהשפעות סביבתיות, שומרת על צורתו החיצונית ומאפשרת לתא להתקיים בפתרונות היפוטוניים. קודם כל, זהו ה"כשרון" של פפטידוגליקן. חדירות והובלה של חומרים לתוך התא קשורה לדופן התא.
מתחת לממברנה הציטופלזמה בחיידקים נמצאת הציטופלזמה. מדובר במערכת קולואידית המורכבת ממים, חלבונים, שומנים, פחמימות, תרכובות מינרלים וחומרים נוספים, שיחסם משתנה בהתאם לסוג החיידקים וגילם. הציטופלזמה של תא פרוקריוטי אינה נתונה לתנועה מחזורית, כמו בצמחים, או תופעות התכווצות, כמו באמבות. תאים חיידקייםמבצעים תנועה עקב מנגנונים אחרים: או בעזרת דגלים מסודרים פשוטים, השונים מהדגלים המורכבים של אוקריוטים, הם שוחים באופן פעיל במדיום נוזלי או גולשים, מפרישים ריר ונעים לאורך פני השטח של מצע צפוף.
תאים תנועתיים של חיידקים נעים באופן פעיל בכיוון שנקבע על ידי גורמים מסוימים. תנועות מכוונות כאלה של חיידקים נקראות "מוניות". יש chemotaxis, phototaxis, magnetotaxis, thermotaxis, viscositaxis (תגובה לשינוי בצמיגות התמיסה).
תאי חיידקים במנוחה מיוצגים בחיידקים גרם חיוביים על ידי אנדוספורות, בחיידקים גרם שליליים - על ידי ציסטות. לנבגי חיידקים יש עמידות ייחודית להשפעות שליליות. בולטת במיוחד העמידות של נבגי חיידקים לחום: חלק מהנבגים סובלים רתיחה במשך שעה או יותר ואינם מאבדים את הכדאיות. עמיד מאוד בפני נבגים ולייבוש. יציבות הנבגים קשורה בנוכחות של שכבה מיוחדת בדופן התא שלהם - קליפת המוח.
במהלך היווצרות נבגים, תאי חיידק לפעמים מקבלים צורה יוצאת דופן של ציר, לימון או מַקוֹשׁ. האנדוספור הוא בדרך כלל אחד לתא, אך ישנם חיידקים עם יותר מארבעה נבגים. כמה חיידקים אנאירובייםעל נבגים, נוצרים כובעים תאיים, המורכבים מ-vacuoles של גז, או סוגים שונים של תולדות
נבגי חיידקים אינם צורות התרדמה היחידות בפרוקריוטים. חיידקים הקוראים גראם, שאינם מסוגלים ליצור אנדוספורים, הופכים במקרים מסוימים לציסטות - תאים מיוחדיםעם כיסויים חיצוניים עבים (למשל. , Azotobacter chroococcum)
בחוץ, קירות התא של פרוקריוטים מוקפים לעתים קרובות בחומר רירי. תצורות כאלה, בהתאם לתכונות המבניות, נקראות קפסולות, שכבות ריריות, נדן או נדן (בחיידקים חוטיים). כולם הם תוצאה של ביוסינתזה של פולימרים אורגניים על ידי פרוקריוטים ותצהיר שלהם סביב תאים.
כמוסות, תצורות ריר ונדן עשויות להכיל רכיבים זהים לדופן התא, אך הם תרכובת כימיתלא מזוהה. ככלל, ההרכב הכימי של קפסולות הנוצרות על ידי חיידקים הוא ספציפי לסוג ולמין. המרכיבים הכימיים העיקריים של רוב הקפסולות הפרוקריוטיות הם פוליסכרידים. היוצא מן הכלל הוא הקפסולות של כמה סוגים של bacilli, הבנויים מפוליפפטיד, שהוא פולימר של חומצה גלוטמית. מקרים - עוד מבנים מורכבים, יש להם הרכב כימי מורכב יותר ומכילים סוכרים, חלבונים, שומנים.
לקפסולות יש עובי שונה והן מזוהות בקלות מתחת
תחת מיקרוסקופ עם ניגודיות שלילית של התכשיר עם דיו שחור.
קפסולות וכיסויים מגינים על התא מפני נזקים והתייבשות, יוצרים מחסום אוסמוטי נוסף, משמשים כמכשול לחדירת פאג'ים, יוצרים סביב התאים סביבה מיוחדת המספקת משטר המים. לקפסולות יש פונקציות ספציפיות. בחיידקי גושים, למשל, הם קובעים את ההתקשרות הסלקטיבית לשערות השורשים של צמחים ממינים שונים. לפעמים הקפסולות יכולות לשמש כחומרי הזנה רזרבה. בעזרת ריר מתבצעת תקשורת בין תאים במושבה, הצמדות תאים למשטחים שונים
פוליסכרידים כמוסות מקיימים אינטראקציה עם קטיונים של תמיסות אדמה, הם יכולים להרוס מינרלים בקרקע, להשתתף בתהליכי צבירה ויצירת מבנה אדמה עמיד במים. אולי הכללת ריר חוץ-תאי מיקרוביאלי בתהליך היווצרות של חומרים הומוסיים.
צורת התא של פרוקריוטים מגוונת מאוד. פרוקריוטים ידועים הם כדוריים, בצורת מוט, מעוותים ספירלית, בצורת פסיק, בצורת טבעת - סגורה או פתוחה, עם יציאות - פשוטות. תאים עשויים להיות מרובעים או בצורת כוכב. פרוקריוטים רב-תאיים הם בצורת חוט או בצורת תולעת.
כל הפרוקריוטים שייכים לקבוצת האורגניזמים המיקרוסקופיים, גודל התאים שלהם נמדד במיקרונים (מיקרומטרים) (1 מיקרומטר = 10 3 מ"מ), ופרטיהם נמדדים בננומטרים (1 ננומטר = 10 מ"מ). כמעט 40% מהחיידקים בקרקע הם מעבר ליכולת של מיקרוסקופ אור, הם נראים רק במיקרוסקופ אלקטרונים. בגדלים קטנים כאלה, יחס פני השטח לנפח גבוה מאוד, וכתוצאה מכך קצב חילוף חומרים פוטנציאלי גבוה.
כל הפרוקריוטים מחולקים לארבע קבוצות על פי מבנה האינטגמנט החיצוני של התא: חיידקי גרם שליליים (Gracilicutes), eubacteria גרם חיוביים (Firmicutes), מיקופלזמות (Molicutes), נטולי דפנות תאים, וארכיבקטריה, או ארכיאה. (Archaebacteria). שלוש הקבוצות הראשונות מאוחדות כ-eubacteria (Eubacteria), archaebacteria הם קבוצה נפרדת של אורגניזמים (Archaebacteria) הנבדלים מאחרים במבנה של מולקולות RNA ריבוזומליות ומעבירות, RNA פולימראזות, שומנים ומבנה דופן התא.
לחיידקים גראם שליליים יש דפנות תאים, כאשר בנוסף לשכבת המסגרת הראשית של murein, ישנה גם קרום חיצוני ושכבת חלבון ו/או גליקופרוטאין חופפת אותה מלמעלה. יש רווח פריפלזמי בין הממברנה הציטופלזמית לשכבת ה-murein. קבוצת חיידקים זו היא הנרחבת והמגוונת ביותר מבחינת מספר המינים ו פונקציות פיזיולוגיות. קבוצת הפרוקריוטים משלבת חיידקים פוטוטרופיים וכימוטרופיים. ביניהם ספרוטרופים נפוצים, קופיוטרופים, אוליגוטרופים, טפילים תוך תאיים של חיידקים אחרים, צורות פתוגניות לבני אדם, לבעלי חיים ולצמחים עם צרכים תזונתיים מורכבים. לפי מורפולוגיה, מדובר בחיידקים מגוונים מפסאודומונדים פשוטים בצורת מוט ועד מיקסובקטריה עם מחזור התפתחות מורכב.
מיקופלזמות הן חיידקים שאין להם דופן תא, שתפקידו מתבצע על ידי הממברנה הציטופלזמית.
Archaebacteria מאחדים פרוקריוטים בעלי מבנה שונה של המכלול החיצוני, שחסר להם מוראין, האופייני לחיידקים אמיתיים. לחלק מהנציגים של archaebacteria יש פסאודומוראין בדופן התא, בעוד שלאחרים יש דופן תא המורכבת משכבה של חלבונים כדוריים, או גליקופרוטאין.
חלוקת החיידקים לקבוצות על סמך מאפיינים פנוטיפיים (מורפולוגיה, פיזיולוגיה) נוחה למטרות מעשיות לזיהוי מהיר של אורגניזמים. אורגניזמים מקובצים לקבוצות מלאכותיות, או "מקטעים", שאין להם מעמד טקסונומי ולעתים קרובות מסומנים על ידי רובם סוג אופייניאו כותרת. לפי עיקרון זה, נבנה ה-Bacteria's Determinant של Burgey.
על ידי לימוד מגוון האורגניזמים המאכלסים ומאכלסים את כדור הארץ, טקסונומים מבקשים ליצור מערכת פילוגנטית שתשקף את הקשר הגנאלוגי והמקור המשותף של אורגניזמים המאוחדים בקבוצה כזו או אחרת - טקסון. מטרה זו משרתת סיסטמטיקה מולקולרית, המבוססת על הדמיון במבנה הביופולימרים - DNA, RNA, חלבונים. עבור היכולת לזהות את הדמיון של גנים, זה נקרא לעתים קרובות שיטת גנים. אחד ההישגים החשובים ביותר של שיטת הגנים הוא גילוי של קבוצה חדשה של מיקרואורגניזמים מבודדים בממלכה נפרדת - ארכיבקטריה. מידת הדמיון של גנומים חיידקיים נקבעת על ידי השוואת רצפי הנוקלאוטידים במולקולת 16S rRNA. השימוש בשיטה זו בטקסונומיה של חיידקים שימש בסיס לבניית העץ הפילוגנטי של החיידקים על ידי ק.ווס.
קשה מאוד להשוות בין שיטתיות פילוגנטית ופנוטיפית. אותם ענפים פילוגנטיים מכילים אורגניזמים מגוונים ביותר מבחינת הפיזיולוגיה. כאשר עוברים מענפים גדולים של חיידקים גרם חיוביים וגרם שליליים לרמות ההיררכיות הנמוכות יותר, חוסר התאמה כזה הולך וגובר.
עבודת קורס
בביולוגיה
"מקורו של האיקריוט
הושלם על ידי: Severova Vera
כיתה י', בית ספר N55
נבדק על ידי: Strukova N.A.
ריאזאן 1999
1 . תיאוריה סימביוטית:
1.1 הבחנה בסיסית בין פרוקריוטים לאאוקריוטים;
אורגניזמים פרוקריוטיים;
תאים איקריוטיים;
מיטוכונדריה ופלסידים;
היכולת של אוקריוטים לעשות פוטוסינתזה;
2. מבשרי חיים;
3. אורגניזמים מתסיסים:
מקורות אנרגיה של תאים מוקדמים;
מתנוגנזה כמקור אנרגיה;
ציאנובקטריה וכלורווקסיבקטריה;
פוטוסינתזה ואוויר;
אסוציאציות ואיקריוטים:
תיאוריות של סימביוזות עוקבות;
רכישה סימביוטית של ניידות;
אוקריוטים הם פוטוסינתטיים;
מבט על התפתחות התא:
6.1 השערה של השתייכות ישירה;
השערה: מציאנובקטריה ועד אצות;
תאימות של 2 השערות: השתייכות ישירה ו
אנדוזימביוזה;
סִפְרוּת.
התיאוריה של אנדוזימביוזה כל האורגניזמים החיים על פני כדור הארץ מחולקים בדרך כלל לפרוקריוטים ולאאוקריוטים. המאפיין העיקרי של פרוקריוטים, בניגוד לאאוקריוטים, הוא היעדר גרעין תא מלא המכוסה בקרום. תאים פרוקריוטים הם קטנים מאוד בגודלם, בסדר גודל של 1 מיקרומטר. נפח התאים האוקריוטיים המכילים גרעין מלא גדול פי 800-1000 מנפח התאים הפרוקריוטים. בהקשר זה, תאים - אאוקריוטים זקוקים למנגנון ממברנה גדול בהרבה כדי לבצע פונקציות חיוניות. ואכן, בתא איקריוטי זוהו מבנים תת-תאיים המוקפים בממברנות בלתי תלויות בפלזמה ובזה. מבנים אלו נקראים אברונים. לכל אברון: מיטוכונדריה, כלורופלסט, גרעין, ליזוזומים, מנגנון גולגי וכו' יש תפקידים ספציפיים משלו בחיי התא. בעזרת גישות מולקולריות – ביולוגיות הושגו עדויות משכנעות שיש לחלק את כל היצורים החיים ל-3 ממלכות: חיידקים, ארכאים ואיקריוטים. מקורם של חיידקים וארכיאה על פני כדור הארץ מאב קדמון משותף - מה שנקרא פרוגנטו - לפני כ-4 מיליארד שנים, ולפי פליאונטולוגים, איקריוטים הופיעו על פני כדור הארץ 500 מיליון שנה לאחר הפרוקריוטים. הרעיון הפופולרי והמבוסס ביותר על מקורם של תאים אוקריוטים הוא כיום הרעיון של אנדוזימביוזה מרובה של פרוקריוטים שונים. על פי המיקרוביולוגים הידועים א' קלואיבר וק' ואן ניאל, בפרוקריוטים העתיקים התרחשה בחירת התאים עם התגובות הביוכימיות החסכוניות ביותר ולעתים קרובות הרב-פונקציונליות, שהפכו לבסיס של המסלולים המטבוליים החשובים ביותר.
התיאוריה הסימביוטית של מקורם ואבולוציה של תאים מבוססת על שני מושגים חדשים בביולוגיה.לפי הראשון מבין המושגים הללו, ההבחנה הבסיסית ביותר בטבע החי היא ההבחנה בין פרוקריוטים לאאוקריוטים, כלומר. בין חיידקים לאורגניזמים המורכבים מתאי עם גרעינים אמיתיים - פרוטיסטים, בעלי חיים, פטריות וצמחים. הרעיון השני הוא שחלקים מסוימים של תאים אוקריוטים מקורם באבולוציה של סימביוזות, כלומר. היווצרות אסוציאציות קבועות בין אורגניזמים ממינים שונים. ההנחה היא ששלוש מחלקות של אברונים - מיטוכונדריה, ריסים ופלסידים פוטוסינתטיים - מקורם בחיידקים חיים חופשיים, שכתוצאה מסימביוזה נכללו ברצף מסוים בתאי מארחים פרוקריוטיים. תיאוריה זו מסתמכת במידה רבה על רעיונות ניאו-דארוויניסטים שפותחו על ידי גנטיקאים, אקולוגים, ציטולוגים ומדענים אחרים שחיברו את הגנטיקה המנדליאנית עם הרעיון הדרוויניאני של הברירה הטבעית. היא מסתמכת גם על כיוונים מדעיים חדשים לחלוטין או שהתחדשו לאחרונה: על ביולוגיה מולקולרית , במיוחד על מבנה החלבון ונתוני רצף חומצות אמינו, על מיקרופליאונטולוגיה, החוקרת את עקבות החיים המוקדמים ביותר על פני כדור הארץ, ואפילו על פיזיקה וכימיה של האטמוספירה, שכן מדעים אלו קשורים לגזים ממקור ביולוגי. ניתן לקבץ את כל האורגניזמים התאיים לחמש ממלכות: הממלכה הפרוקריוטית (Monera, הכוללת חיידקים) וארבע הממלכות האיקריוטיות (Protoctista, Animalia, Fungi, Plantae). פרוטוקטיסטים הם אורגניזמים אוקריוטיים שאינם בעלי חיים, פטריות או צמחים. הממלכה הפרוטוקטית כוללת אצות, פרוטוזואה, עובשי רפש ושאר אורגניזמים אוקריוטיים שמקורם לא ברור. פרוטיסטים מוגדרים בצורה מגבילה יותר כאאוקריוטים חד-תאיים. לפיכך, הממלכה פרוטוקטיסטה כוללת לא רק פרוטיסטים - אוקריוטים חד-תאיים, אלא גם את צאצאיהם הרב-תאיים הקרובים ביותר, כגון אצות אדומות וחומות, וכן מיקרואורגניזמים רבים הדומים לפטריות, כגון כיטרידים. על פי התפיסה המסורתית של פיליון ישיר, אברוני תאים כגון מיטוכונדריה ופלסידים התעוררו באמצעות מידור של התא עצמו.
מספר פרוקריוטים יוצרים איקריוט אחד
תאים הם מערכות הקשורות לממברנה, הקטנה ביותר מבין היחידות הביולוגיות המסוגלות להתרבות עצמית. אפילו התאים הקטנים ביותר מכילים גנים בצורת מולקולות DNA (אחת או יותר) ומנגנון לסינתזה של חלבונים, המורכב מכמה סוגים של RNA וחלבונים רבים. כל התאים מכילים ריבוזומים, גופים בקוטר של כ-0.02 מיקרומטר, המורכבים משלושה סוגי RNA לפחות וכחמישים חלבונים שונים. אורגניזמים פרוקריוטיים הם היחידות הביולוגיות הקטנות ביותר העונות על הגדרה זו של תא. אין להם גרעינים. מצד שני, תאים אוקריוטיים בודדים (בעלי גרעין) הם, במובן מסוים, כלל לא יחידות. אלו הם קומפלקסים של יחידות שונות של סינתזה של חלבון (נוקלאוציטופלזמה, מיטוכונדריה ופלסטידים), שסוגם ומספרם משתנה בהתאם למין. לפיכך, לפי תיאוריה זו, תאים אוקריוטיים התעוררו כתוצאה משיתוף פעולה של אלמנטים עצמאיים בתחילה, מאוחדים בסדר מסוים. טיילור כינה את הרעיון הזה של המקור וההתפתחות הנפרדים של חלקים של תא אוקריוטי ואיחודם לאחר מכן בתור התיאוריה של אנדוסימביוזות עוקבות. תנאים של טמפרטורה וחומציות גבוהות הביאו לייצור בנוקלוציטופלזמה של מחלקה מיוחדת של חלבונים - היסטונים - כדי להגן על ה-DNA שלו. מיטוכונדריה הצליחו לפרק תרכובות שלושה פחמן לפחמן דו חמצני ומים. הקשר הסימביוטי של נוקלוציטופלזמה ומיטוכונדריה החל להשתנות בכיוון של חיזוק התלות ההדדית ביניהם. קומפלקס זה כלל אברונים מוטוריים - ריסים ודגלים. טרנספורמציה זו גררה טרנספורמציה מורפוגנטית, שכללה התפתחות של אסימטריות תאיות בולטות. עם שילוב של נוקלוציטופלזמה תלויה הדדית, מיטוכונדריה, אונדוליפודיה, הושלמה יצירת מבנה פוליגנומי של תא אוקריוטי. התא האיקריוטי הראשון היה הטרוטרופי. היא אכלה תרכובות אורגניות מוכנות. באוקריוטים אלה התעוררו תהליכי המיטוזה, ולאחר מכן המיוזה. ולבסוף, רכישת יכולת הפוטוסינתזה על ידי איקריוטים התרחשה בתהליך הפיכתו לארגון האיקריוטי הראשי ולאחר השלמתו - האירוע האחרון באפוס הזה. הסימביוזות הלכו והשתלבו. נכון לעכשיו, התלות של כל אברון בתוצרים המטבוליים של אברונים אחרים הפכה כה מלאה שרק שיטות ניתוח מודרניות מאפשרות להתחקות אחר המסלולים המטבוליים של השותפים המקוריים. אוקריוטים מגוונים במבנה שלהם, אך מבחינה מטבולית הם נשארים אחידים. הטריקים המטבוליים של האוקריוטים פותחו על ידי חיידקים שונים עוד לפני הזמן שבו הם התחברו והפכו למארח ולאברוניו. פרוקריוטים השיגו גיוון רב באופן שבו הם משיגים אנרגיה ובמורכבות הביוסינתזה. הם השאירו זכר לקיומם הרבה לפני שנוצרו אורגניזמים גדולים כל כך כמו חיות עם שלד מהתא האיקריוטי.
ערכת המקור של תאים אוקריוטיים על ידי סימביוזה
מבשרי חיים
אבותיהם של האורגניזמים של ימינו צמחו לפני יותר משלושה מיליארד שנים מתרכובות אורגניות שנוצרו על פני כדור הארץ. הערעור של מושג זה נובע בחלקו מיכולת הבדיקה שלו.
ס' מילר וה' אוריי הראו כי בתנאים האטמוספריים שהתרחשו על כדור הארץ הצעיר, יכולות להיווצר חומצות אמינו ומולקולות אורגניות אחרות. סוגים שונים של מולקולות אורגניות נוצרו באופן ספונטני במהלך מיליארד השנים הראשונות לקיומו של כדור הארץ. מולקולות אורגניות רבות נמצאו בחלל הבין-כוכבי ובמטאוריטים. המשמעות היא שתרכובות אורגניות מתעוררות במהלך האינטראקציה של יסודות קלים, בנוכחות מקורות אנרגיה. האוניברסליות של המנגנון הכימי של רבייה ביולוגית פירושה שכל החיים על פני כדור הארץ מגיעים מאבות קדמונים משותפים: כל החיים הנוכחיים מקורם בתאים עם מערכות מידע המבוססות על שכפול DNA ועל סינתזת חלבון המכוונת על ידי RNA שליח.
אורגניזמים מתסיסים
ייתכן שחומצות גרעין הופרדו מהסביבה כבר מההתחלה על ידי ממברנות ליפופרוטאין. מידע החל להיות מקודד על ידי רצף הנוקלאוטידים במולקולות ה-DNA. התאים העתיקים ביותר השתמשו ב-ATP ובחומצות אמינו, שהן חלק מחלבונים, כמקורות ישירים לאנרגיה וכמרכיבים מבניים. אז - החיים העתיקים ביותר היו קיימים בצורה של חיידקים הטרוטרופיים שקיבלו מזון ואנרגיה מחומר אורגני ממקור אביוטי. אבולוציה אורגנית ניאו-דרוויניסטית דורשת שלוש תופעות: רבייה, מוטציה ולחץ סביבתי סלקטיבי. רבייה היא מצב בלתי ישים של אבולוציה ביולוגית. עם רבייה מדויקת מספיק, מוטציות וברירה טבעית הן בלתי נמנעות. להסביר את מקורה של רבייה כזו היא להבין את מקור החיים עצמם. מוטציה, המקור העיקרי לשונות באבולוציה, מובילה לשינויים תורשתיים במערכת רבייה עצמית. לאחר שפותחה רבייה אמינה, החלה האבולוציה הניאו-דרוויניסטית. לאורגניזמים שפיתחו מסלולים לסינתזה של הרכיבים הדרושים לתא היה יתרון על פני אחרים. חיידקים המייצרים מתאן אנאירובי היו האוטוטרופים העתיקים ביותר, כלומר, אורגניזמים המסוגלים לספק את צורכיהם לאנרגיה ולחומרים אורגניים ממקורות אנאורגניים. יכולת הפוטוסינתזה פותחה לראשונה בחיידקים הרגישים לחמצן. ספגו את אור השמש בעזרת כלורופיל חיידקי, הם המירו את הפחמן הדו-חמצני של האטמוספירה לחומרים אורגניים.הופעת הפוטוסינתזה האנאירובית שינתה באופן בלתי הפיך את פני השטח ואת האטמוספירה של כדור הארץ.
פחמן דו חמצני הוצא מהאטמוספרה והפך לחומר אורגני בהתאם למשוואת הפוטוסינתזה הכללית: 2H2X+CO2=CH2O+2X+H2O. חיידקים פוטוסינתטיים שהשתמשו במימן גופרתי להפחתת CO2 הפקידו גופרית יסודית. הם הולידו חיידקים המסוגלים לבצע פוטוסינתזה עם שחרור חמצן. סוג זה של פוטוסינתזה מקורו כנראה כתוצאה מהצטברות של מוטציות בחיידקי גופרית פוטוסינתטיים. נתיב זה הוביל להופעתם של אצות כחולות ירוקות או ציאנובקטריה. לאחרונה התגלתה ונחקרה קבוצה חדשה של פרוקריוטים המייצרים O2 פוטוסינתטי, כלורווקסיבקטריה. ניתן לשער שדווקא ציאנו- וכלורווקסיבקטריה היו אבותיהם של הפלסטידים המצויים בתאי האצות והצמחים. כאשר O2 הצטבר כתוצאה מפוטוסינתזה מיקרוביאלית, אטמוספירת כדור הארץ הפכה להתחמצנות, מה שגרם למשבר של כל החיים - עד לאותה תקופה העולם היה מיושב בחיידקים אנאירוביים. כדי לשרוד, החיידקים נאלצו להימנע מחמצן על ידי הסתתרות בביצות, או שהם היו צריכים לפתח חילוף חומרים המסוגל להגן עליהם מפני כמות החמצן הגוברת.
פוטוסינתזה ואוויר
לפני כ-2 מיליארד שנים, בזה אחר זה, התרחשו תהליכים אבולוציוניים שונים ליצירת מגוון מטבולי. את היכולת לייצר אנזימים ספציפיים ניתן להעביר בתוך אותו מין באמצעות חלקיקים דמויי וירוס הנקראים פלסמידים. היכולות הביוסינתטיות של פרוקריוטות בולטות: מפחמן דו חמצני, מספר מלחים ותרכובות אנאורגניות מופחתות המשמשות כמקורות אנרגיה, מיקרואורגניזמים כימואוטוטרופיים יכולים לסנתז במים לא מוארים המכילים חמצן את כל המקרומולקולות המורכבות ביותר הנחוצות לצמיחה ורבייה. תהליכים ביוכימיים אלו הובילו למערכות של מחזור היסודות גם כאשר לא היו צמחים, פטריות, בעלי חיים.
אסוציאציות ואיקריוטים.
מבט על התפתחות התא.
סניף ישיר.
ההשקפה המסורתית לגבי מקורם והתפתחותם של תאים אוקריוטיים היא שכל היצורים החיים צאצאו ישירות מאוכלוסיית אבות אחת כתוצאה מהצטברות של מוטציות בהשפעת הברירה הטבעית.
מוטציות נקודתיות, כפילויות, מחיקות ושינויים תורשתיים אחרים, שידוע כי מילאו תפקיד באבולוציה של בעלי חיים וצמחים, אחראים גם להתמיינות של תאים אוקריוטיים מתאים פרוקריוטיים - תורת הפיליון הישירה. כלומר, הרעיון של מקור לא סימביוטי של תאים. אולי אצות ירוקות הן צאצאים של אבותיהם של כל הצמחים. תכונות רבות של אצות אלה מעידות על כך שיש להן ולצמחים אב קדמון משותף. הפילוגניה של בעלי חיים מורכבת יותר, אבל המושג של השתייכות ישירה חל כנראה גם על הנתיבים האבולוציוניים שלהם. ספוגים ככל הנראה התפתחו מפרוטיסטים, ללא תלות בקבוצות אחרות של בעלי חיים. אז, לפחות עבור בעלי חיים וצמחים, ההשערה שאורגניזמים אוקריוטיים מורכבים התפתחו מאיקריוטים מסודרים יותר באמצעות הצטברות של מוטציות בודדות בהשפעת הברירה הטבעית היא לגיטימית למדי. חידושים אבולוציוניים מסתמכים על שימור של גנים חיוביים מושלמים ביותר. אז, אורגניזמים חדשים בעלי יכולת מוגברת לשלוט בסביבה שלהם התפתחו יותר על ידי שמירה על שכפול ושילוב מחדש של גנים מותאמים מאוד מאשר על ידי רכישת תכונות חדשות באמצעות מוטציות נקודתיות פשוטות ואקראיות. ההשערה של השתייכות ישירה אינה מסבירה את מקורם של איקריוטים מפרוקריוטים. לדוגמא, קשה להבין את ההפסקה בין ציאנובקטריות שאינן מסוגלות למיטוזה לבין צורות כאלה עם מיטוזה מפותחת כמו אצות אדומות מנקודת מבט של השתייכות ישירה, אך היא נובעת באופן טבעי מהתיאוריה הסימביוטית.
על פי תורת ההשתייכות הישירה ותיאוריית הסימביוזה, כל האורגניזמים על פני כדור הארץ מקורם באבות חיידקים דומים מבחינה ביוכימית, וחיידקים אוקריוטיים הולידו צמחים ובעלי חיים. חיידקים פוטוסינתטיים הולידו אצות, ובסופו של דבר צמחים, וכמה אצות איבדו את הפלסטידים שלהם והפכו לאבותיהם הקדמונים של פטריות ובעלי חיים. תיאורטיקנים של פיליה ישירה מסכימים שפלסטידים ואברונים אוקריוטיים אחרים, כולל הגרעין, נוצרו על ידי התמיינות בתוך התאים עצמם ומכנים זאת מיתוס בוטני.
פילוגניה של סוגי צמחים
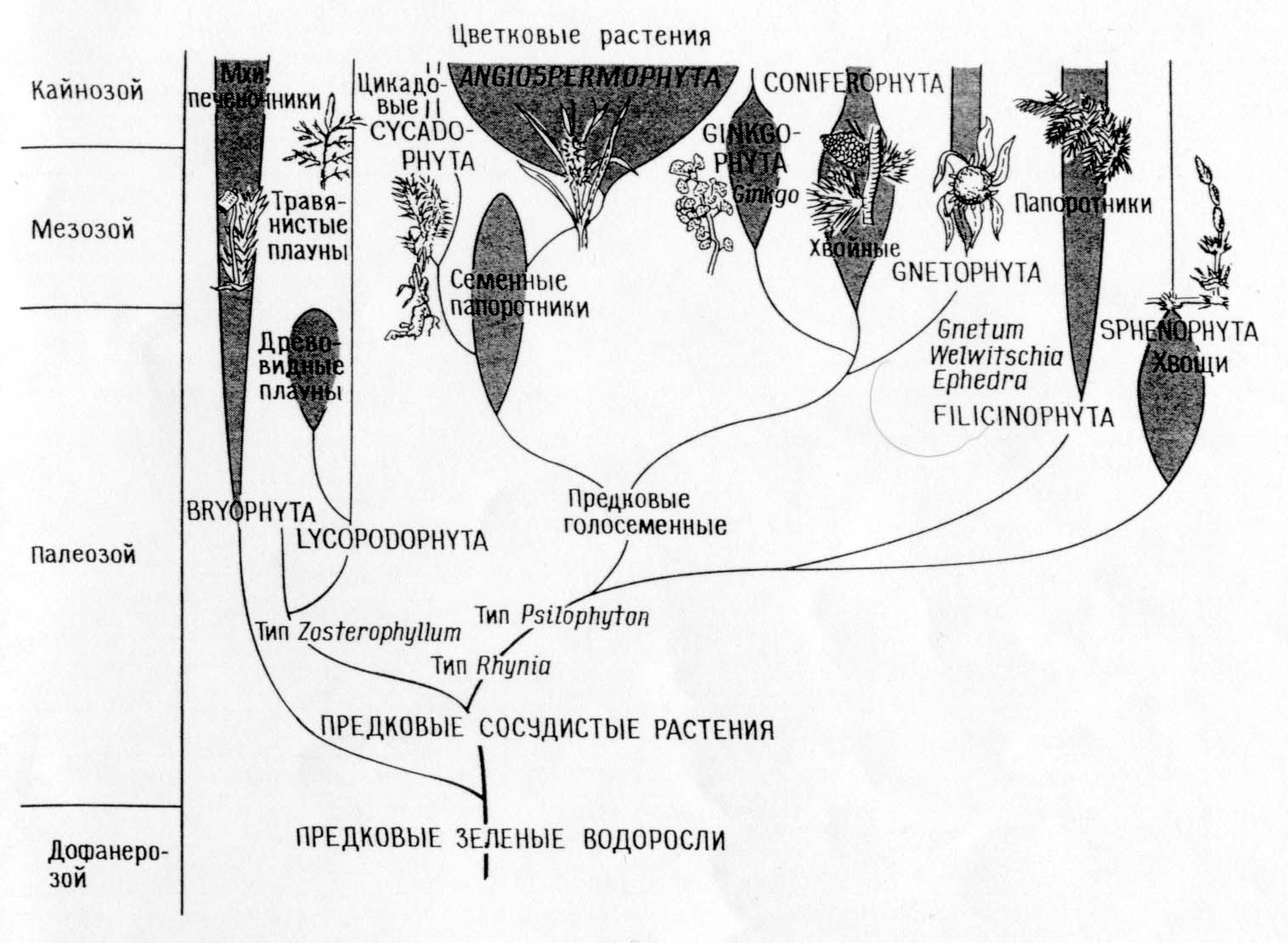
השוואה של פילוגניות התואמות לתיאוריית ההשתייכות הישירה (A)
ותאוריית הסימביוזה (ב)
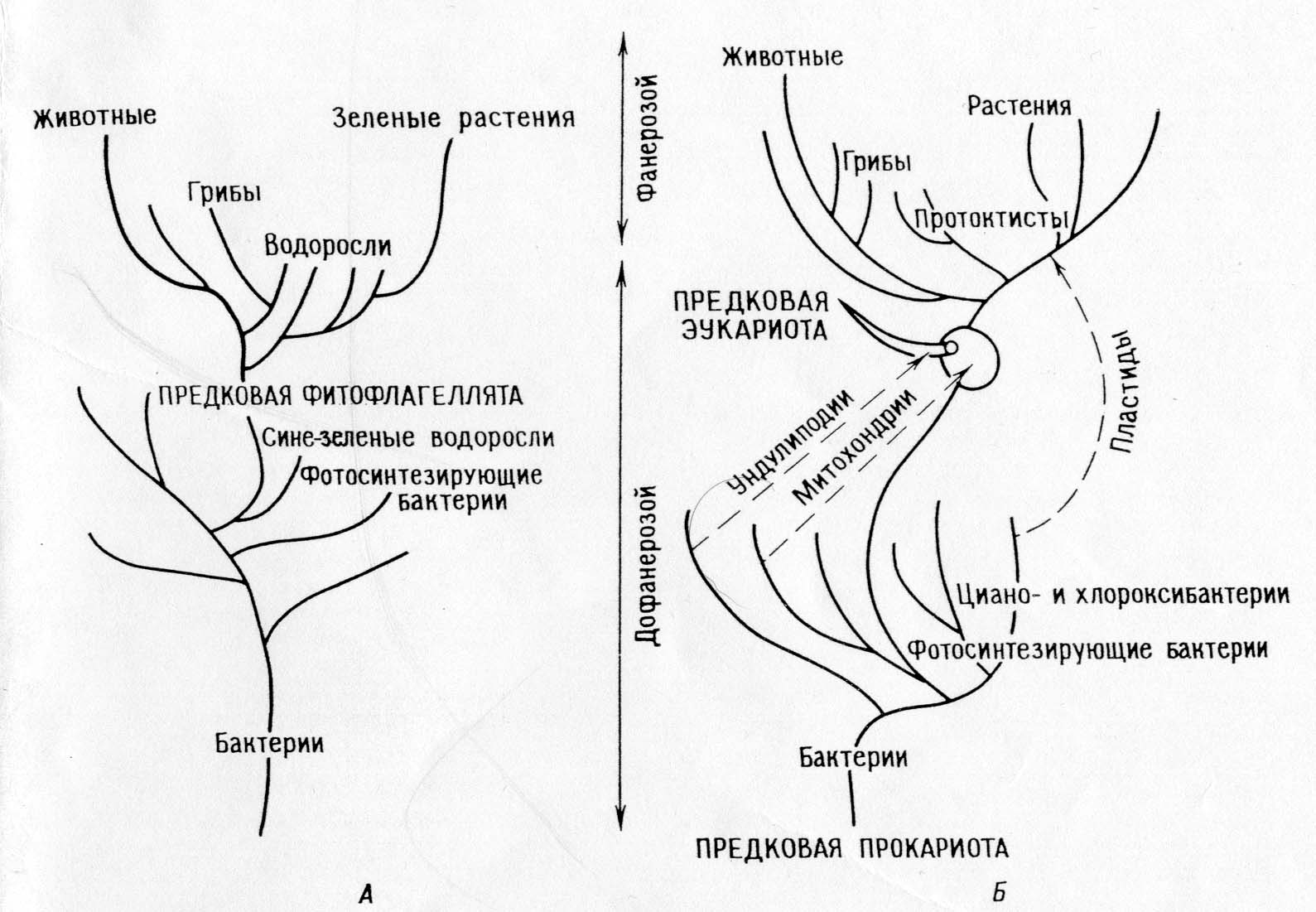
הבדלים בין תיאוריית ההשתייכות הישירה לתיאוריה הסימביוטית:
התיאוריה של השתייכות ישירה.
דיכוטומיה עיקרית: בעלי חיים - צמחים
איקריוטים פוטוסינתטיים (אצות וצמחים ירוקים) התפתחו מפרוקריוטים פוטוסינתטיים (אצות כחולות-ירקות)
צמחים ומערכות הפוטוסינתזה שלהם התפתחו באופן מונופילטי בפרקמבריון.
בעלי חיים ופטריות התפתחו מאוטטרופים שאיבדו פלסטידים.
המיטוכונדריה התמיינה תוך תאית באצות פוטוסינתטיות.
בעשבים הימניים, האונדוליפודיה, המנגנון המיטוטי וכל שאר האברונים האוקריוטיים התמיינו תוך תאי.
כל האורגניזמים צאצאו מאבותיהם - פרוקריוטים על ידי הצטברות של מוטציות בודדות.
תוצאה: על פי התיאוריה, חייבים להיות עשבים נכונים; קשה לקשר את התיאוריה הזו לתיעוד המאובנים ולבנות פילוגניה מתאימה.
התיאוריה של אנדוסימביוזות עוקבות:
הדיכוטומיה העיקרית: פרוקריוטים - איקריוטים.
איקריוטים פוטוסינתטיים (אצות וצמחים גרעיניים) ואיקריוטים שאינם פוטוסינתטיים (בעלי חיים, פטריות, פרוטוזואה) התפתחו מאבות קדמונים הטרוטרופיים נפוצים (אמבופלגלטים).
האבולוציה של הפוטוסינתזה בארכאים הייתה מונופילטית בחיידקים (כולל ציאנו-וכלורווקסיבקטריות), פרוטיסטים פוטוסינתטיים התעוררו לפתע כאשר הפרוטיסטים-אבותיהם ההטרוטרופיים נכנסו לסימביוזה עם פרוקריוטים פוטוסינתטיים (פרוטופלסטידים).
בעלי חיים ורוב האצות והפטריות הם צאצאים ישירות מפרוטיסטים הטרוטרופיים.
מיטוכונדריה כבר היו נוכחים בפרוטיסטים הטרוטרופיים כאשר האחרונים רכשו פלסטידים כתוצאה מסימביוזה. פרומיטוכונדריה נרכשו בעבר באופן סימביוטי.
מיטוזיס התפתחה בפרוטיסטים אוקריוטים הטרוטרופיים על ידי התמיינות של המערכת הבלתי-ליפודיאלית.
פרוקריוטים התפתחו מאבות פרוקריוטים מוקדמים דרך סדרה של אירועים מוטציוניים בודדים, איקריוטים התפתחו גם מאבות אוקריוטים דרך אירועים מוטציוניים בודדים, ואבות איקריוטים התפתחו מפרוקריוטים דרך סדרה של סימביוזות.
השלכות: התיאוריה מסבירה מסלולים ביוכימיים ואת יתירותם באברונים. זה מאפשר בניית פילוגניה עקבית שמסבירה את "הפער הביולוגי" בין הזמנים הפרוטרוזואיקונים והפנרוזואיקונים. התיאוריה חוזה שאברונים חייבים לשמור על שרידים של מערכות שכפול.
מיתוס בוטני
מימי ז'וסייה נקראים הצמחים הנמוכים טלופיטים, והגבוהים נקראים בריופיטים וטראכאופיטים. על פי המיתוס הבוטני, האבות הקדמונים המשותפים של חיידקים פוטוסינתטיים ואצות כחולות-ירוקות, באמצעות שיוך ישיר, הולידו אצות אוקריוטיות, כולל אצות ימיות. קבוצות של אורגניזמים שהפילוגניה שלהם מושפעת יותר מכל מהמושג של פיליון ישיר הגיעו בסופו של דבר לתחום הבוטניקה - אלו הם חיידקים, אצות, פטריות וצמחים.
אורגניזמים היפותטיים שיכולים להוות קישור בין ציאנובקטריה לאצות ירוקות נקראו פיטומונאדים קדומים (פיטומונדים קדומים) או עשבים ימניים. כמה מחברים טענו שאאוקריוטים Cyanidium ו- Cyanophora, הדומים במערכות הפיגמנט שלהם לציאנובקטריה, עשויים להיות מועמדים מתאימים לתפקיד של עשבים ימניים. בטלופיטים לא נמצאו צורות ביניים בין המצב הלא-מיטוטי של ציאנובקטריה לבין המיטוזה האופיינית המפותחת במלואה של אצות ירוקות וצמחים. למרות שפטריות וצמחים פורחים נטולי אונדוליפודיה לחלוטין, הם מתאפיינים במיטוזה ומיוזה, הדורשים צירים מיקרוטובוליים. ישנה הסכמה כללית שכל האורגניזמים הרב-תאיים עם מיקרו-צינוריות הם צאצאים ממיקרואורגניזמים אוקריוטיים עם אונדוליפודיה, אך איש אינו יודע מאיזו קבוצה של פרוטיסטים. חיזק את המיתוס הבוטני היה העתיקות והמורכבות של הפוטוסינתזה, כמו גם העובדה שלאורגניזמים רבים (חיידקים, אוגלנואידים, צמחים טפילים), הן בטבע והן במעבדה, היה אובדן פוטוסינתזה כתוצאה ממוטציות. כיוון שפוטוסינתזה היא תהליך אנאירובי המבטיח בסופו של דבר את קיומם של כל האורגניזמים, היא בוודאי התפתחה בשלב מוקדם מאוד בהיסטוריה של החיים על הפלנטה שלנו, עוד לפני שהתעוררה מיטוזה הדורשת חמצן. פיגמנטים, ליפידים ואנזימים רבים מעורבים בפוטוסינתזה, מסודרים בצורה מסודרת בממברנות מורכבות מאוד. יחד עם זאת, הפרטים של מנגנוני פוטוסינתזה באצות ובצמחים מחד ובציאנובקטריה מאידך דומים להפליא. כל האורגניזמים הללו משחררים גז חמצן מהמים, המשמש כמקור לאטומי מימן המשמשים להפחתת CO2 לחומר האורגני של התא. לאצות ולפרוטוזואים רבים אין לא מיטוזה ולא מערכת גנטית דומה לזו של בעלי חיים וצמחים. לסילאטים מערכת גנטית מיוחדת עם 2 סוגי גרעינים, ולרוב התאים הם מרובי גרעינים. סוג אחד של גרעינים הוא מקרו-גרעינים, והשני הוא מיקרו-גרעינים קטנים בהרבה. מיקרו-גרעינים הם חסרי משמעות פיזיולוגית. עם זאת, הם מאחסנים עותקים של גנים, ובמהלך התהליך המיני, הם עוברים מיוזה. גרעיני מקרו חיוניים מבחינה פיזיולוגית משמשים כאתר של שעתוק RNA ובכך שולטים בפיזיולוגיה של התא. אחד הקשיים בבירור הקשר בין ציאנובקטריה לאצות אוקריוטיות הוא המחסור בנתונים על מקורה של אונדוליפודיה: לא נמצאו קשרי ביניים בין ציאנובקטריה, שאף פעם לא קיימת בה, לבין האצות הרבות שיש. בעיה נוספת היא היעדר מוחלט של התהליך המיני בציאנובקטריה: בין ציאנובקטריה א-מינית לאצות, שרובן מאופיינות בתהליך המיני, לא היו צורות ביניים. רוב האצות נמצאו כסובלות מ-undolipodia בשלב כלשהו במחזור החיים שלהן. התברר שהמבנים הניידים באצות, פרוטוזואה, בעלי חיים וצמחים זהים: כולם מורכבים ממיקרו-צינוריות בקוטר של 0.024 מיקרון. התברר שהציר המיטוטי, גם בפטריות שאין להן אונדוליפדיה, מורכב ממיקרוטובולים חלבוניים בקוטר זהה לזה של אונדוליפדיה. במשך חצי מאה, זה נראה סביר ביותר שאצות אוקריוטיות התפתחו מציאנובקטריה. מיתוס בוטני זה הוביל להנחות שבמשך שנים רבות שימשו הן כגירוי למחקר והן כמסגרת התייחסות שבה הוערכו התוצאות. הקביעה שהקשרים בין ציאנובקטריה לאצות איקריוטיות גוועו, ולא הותירו עקבות מאובנים ושרידים חיים, הפכה פחות ופחות משכנעת ככל שנתונים חדשים על חיידקים ומאובנים חיים. קשרי הביניים העיקריים בין פרוקריוטים פוטוסינתטיים לאאוקריוטים לא גוועו בלי זכר, כמתחייב מהמיתוס הבוטני - הם מעולם לא היו קיימים. לפי התיאוריה של אנדוסימביוזות עוקבות, ציאנובקטריות הן אכן האבות הקדמונים, אבל רק פלסטידים, ולא שאר התא האיקריוטי, ובוודאי לא הגרעין שלו. ובכל זאת, בשאלת מקורו של הגרעין, השערת ההשתייכות הישירה ותורת הסימביוזה תואמות. שינויים אפשריים לגרסה הקיצונית של תיאוריית הסימביוזה כוללים, למשל, את הרעיון של מקור סימביוטי של פלסטידים, אך לא מיטוכונדריה, או של מקור סימביוטי של פלסטידים ומיטוכונדריה, אך לא אונדוליפודיה. תומכי ההשתייכות הישירה הציגו גם הנחה לגבי מקורו של הגרעין באמצעות סימביוזה.
עדות קריטית למקור האנדוזימביוטי של כלורופלסטים ומיטוכונדריה.
העדות המשמעותית ביותר למקורם של האברונים החשובים ביותר של תאים אוקריוטיים – כלורופלסטים ומיטוכונדריה מפרוקריוטים התקבלה בשנים האחרונות כאשר חוקרים את מבנה ה-RNA הריבוזומלי. ביולוג מולקולרי
K. Woz הראה כבר בשנות ה-80 שהמבנה של RNA מהריבוזומים של כלורופלסטים צמחיים כמעט זהה למבנה של RNA מכמה ציאנובקטריות. במקביל, קבוצת חוקרים מצאה שה-RNA של ריבוזומים שמקורו במיטוכונדריה של אוקריוטים שונים דומה ל-RNA של כמה חיידקים, בפרט של חיידקים מהסוג Paracoccus. סט אנזימי הנשימה של חיידק זה דומה מאוד ל
"אנסמבל נשימתי" של מיטוכונדריה של בעלי חיים.
נתונים אלה מעידים בצורה ברורה למדי לטובת ההסתברות הגדולה יותר למקור האנדוסימביוטי של תאים אוקריוטיים. תופעת האנדוזימביוזה עדיין נפוצה ביותר בטבע. ידוע שלעתים קרובות חיידקים זוהרים מתיישבים על גופם של כמה דגים ימיים, אשר לא רק מקבלים יתרונות גדולים עבור חייהם, אלא גם שימושיים מאוד עבור אורגניזמים מארח. כל האמור לעיל מעיד באופן משכנע בעד המקור האנדוסימביוטי של תאים אוקריוטיים מתאי פרוקריוטים קטנים בהרבה. עם זאת, השאלה העיקרית היא לקבוע את אופיו של התא המארח, מכיוון שלתאים אוקריוטים יש גרעין בנוי היטב עם קליפה, בעוד לפרוקריוטים אין גרעין כזה. ל' מרגליס, למשל, סבור שתפקידם של תאי מארח יכול להיות שמילאו פרוקריוטים - קודמי החיידקים המודרניים, השייכים למיקופלזמות - פרוקריוטים הבנויים בצורה פרימיטיבית ביותר, נטולי דפנות תאים כמעט. לאחרונה, יצירותיו של המדען היפני הופיעו ונדונות רבות.
ט אושימה. בהתבסס על מחקר יסודי של המבנה של RNAs ריבוזומליים של פרו-אוקריוטים רבים, כולל נציגים רבים של ממלכת העל של הארכאה, שנחקרה מעט עד כה, מסיק מחבר זה שהתא המארח היה אחד מאבותיה הקדמונים של הארכאה המודרנית. יש לומר כי לנציגי ממלכת העל הזו, בניגוד לחיידקים, יש הרבה תכונות ביולוגיות ביוכימיות ומולקולריות במשותף לאאוקריוטים. בעבודותיו האחרונות של T. Oshima, ניתנים טיעונים בעד העובדה שהתא המארח בזמן הופעת האאוקריוטים היה האב הקדמון של הארכאה, בדומה לאחד הנציגים המודרניים של הפרוקריוטים הללו - תרמופלזמה. מה יהיה הפתרון הסופי לבעיית ביסוס טבעו של התא המארח שהוליד את התא האוקריוטי בתהליך של אנדוזימביוזה נפוצה יראה על ידי מחקרים נוספים.
כעת מוצגת ההשערה הפופולרית והמבוססת ביותר לגבי מקורם של תאים אוקריוטיים
אנדוזימביוזה של פרוקריוטים שונים השייכים לחיידקים ולארכיאה, שהתרחשה פעמים רבות בתהליך האבולוציה במהלך 1.5-2 מיליארד שנות קיומם הראשונות על פני כדור הארץ של תאים חיים. עדות נחרצת למקור האנדוזימביוטי של הכלורופלסטים והמיטוכונדריות התקבלה על ידי הביולוגים המובילים בעולם בחקר מבנה ה-RNA הריבוזומלי. נתונים אלה מעידים בצורה ברורה למדי לטובת ההסתברות הגבוהה למקור אנדוזימביוטי של תאים אוקריוטיים מתאי פרוקריוטים קטנים בהרבה (במספר סדרי גודל).
סִפְרוּת
כתב עת חינוכי של סורוס. ביולוגיה, כימיה, גיאוגרפיה, פיזיקה, מתמטיקה. N 5 1998
Margelis L. תפקיד הסימביוזה באבולוציה של התא. לְכָל. מאנגלית, M. MIR 1983
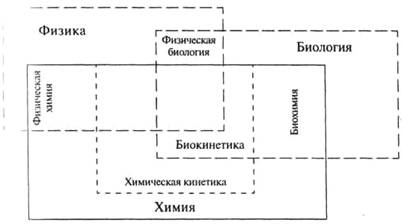
מתוך החלקים העיקריים של הקינטיקה, הוא חוקר את הקינטיקה של תגובות ביולוגיות; סעיף זה נקרא ביו-קינטיקה. ביו-קינטיקה היא מדע גבול שצמח בצומת של ביוכימיה וקינטיקה כימית (ראה איור 1). הבחירה בביו-קינטיקה כדיסציפלינה נפרדת אינה מקרית, היא מוצדקת מבחינה לוגית וקשורה לחשיבות יוצאת הדופן של תהליכים קינטיים עבור כל האורגניזמים החיים. ביו-קינטיקה...
גנים, אך גם יצרו את התנאים המוקדמים להצטברות שינויים בהם, התבדלות הגנים והגדלת מגוון המוצרים הנשלט על ידם. 2. אלמנטים גנטיים ניידים תפקיד מסוים באבולוציה של הגנום של תאים פרו-אוקריוטיים כאחד שייך למה שנקרא אלמנטים גנטיים ניידים - טרנספוזונים. הן יחידות אוטונומיות הנושאות את הנוקלאוטיד...
כַּמָה גישות אפשריותלפיתוח העתיד תיאוריה כללית מערכות דינמיות. בפרק 10, התהליכים המעגליים של החיים, אנו רואים את הארגון המחזורי האופייני של ארגון עצמי מערכות פיזור. הסכימה המוכללת מציבה את המאפיינים של יצירת מחזורי שינוי ומחזורים של תגובות קטליטיות, כמו גם היפר-מחזורים קטליטיים בהתאם ל...