Mikroorganizmy (z łac. micros – małe) to organizmy niewidoczne gołym okiem. Należą do nich pierwotniaki, krętki, grzyby, bakterie, wirusy, które są badane przez mikrobiologię. Wielkość mikroorganizmów mierzona jest w mikrometrach (µm). W mikrokosmosie istnieje ogromna różnorodność form, które są podzielone na grupy na podstawie ogólne zasady klasyfikacja biologiczna.
Pierwszy generał klasyfikacja biologiczna został stworzony w XVIII wieku przez szwedzkiego naukowca K. Linneusza, na podstawie cechy morfologiczne i zawierał zwierzęta i świat roślinny. Wraz z rozwojem nauki klasyfikacja zaczęła uwzględniać nie tylko cechy morfologiczne, ale także fizjologiczne, biochemiczne i genetyczne mikroorganizmów. Obecnie nie można mówić o jednolitej klasyfikacji wszystkich organizmów żywych: przy zachowaniu tych samych zasad klasyfikacje makro- i mikroorganizmów mają swoje własne cechy.
Główne etapy wszystkich klasyfikacji to: królestwo - dział - klasa (grupa) - rząd - rodzina - rodzaj - gatunek. Główną kategorią klasyfikacyjną jest gatunek - zbiór organizmów, które mają wspólne pochodzenie, podobne morfologiczne i objawy fizjologiczne i metabolizm.
Mikroorganizmy należą do królestwa prokariontów, których przedstawiciele w przeciwieństwie do eukariontów nie posiadają sformalizowanego jądra. Informacja dziedziczna u prokariontów jest zawarta w cząsteczce DNA znajdującej się w cytoplazmie komórki.
Dla mikroorganizmów przyjęty w 1980 r., jednolity klasyfikacja międzynarodowa, który opiera się na systemie zaproponowanym przez amerykańskiego naukowca Bergiego.
Aby określić, do jakiego gatunku należy mikroorganizm, konieczne jest użycie różne metody zbadać jego cechy (kształt komórki, zarodnikowanie, ruchliwość, właściwości enzymatyczne) i znaleźć jego pozycję systematyczną za pomocą wyznacznika - zidentyfikować go.
W obrębie gatunku istnieją warianty: warianty morfologiczne różnią się morfologią, warianty biologiczne różnią się właściwości biologiczne, chemowarianty - przez aktywność enzymatyczną, serowarianty - przez strukturę antygenową, warianty fagowe - przez wrażliwość na fagi.
Do oznaczenia mikroorganizmów przyjęto ogólną biologiczną nomenklaturę binarną lub dwumianową (podwójną) wprowadzoną przez K. Linneusza. Imię oznacza rodzaj i jest pisane Wielka litera. Drugie imię oznacza gatunek i jest pisane małą literą. Na przykład Staphylococcus aureus to Staphylococcus aureus. Nazwy mogą odzwierciedlać nazwiska naukowców, którzy odkryli mikroorganizmy: brucella - na cześć Bruce'a, Escherichia - na cześć Eschericha itp. Szereg nazw obejmuje narządy, które wpływają na ten mikroorganizm: pneumokoki - płuca, meningokoki - opony mózgowe itp.
bakteria
bakteria jest Jednokomórkowe organizmy pozbawiona chlorofilu. Średnie rozmiary komórka bakteryjna- 2-6 mikronów. Wielkość i kształt komórek bakteryjnych charakterystycznych dla mikroorganizmów pewien rodzaj, może ulec zmianie pod wpływem różne czynniki(w zależności od wieku kultury bakteryjnej, siedliska itp.). Zjawisko to nazywa się polimorfizmem.
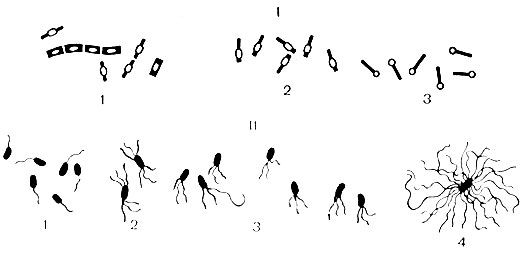
Ze względu na kształt komórki bakterie dzielą się na trzy grupy: kuliste, pałeczkowate i skręcone (ryc. 4).
bakterie kuliste nazywane są ziarniakami (od łac. coccus - jagoda) i mają średnicę komórki od 0,5 do 1 mikrona. Kształt ziarniaków jest różnorodny: kulisty, lancetowaty, w kształcie fasoli. Przez względne położenie rozróżnia się komórki po podziale wśród ziarniaków: mikrokoki (od łac. micros - małe) - komórki dzielą się w różnych płaszczyznach i są zlokalizowane pojedynczo; diplococci (od łac. diploos - podwójne) - komórki dzielą się w jednej płaszczyźnie, a następnie układają w pary; należą do nich lancetowate pneumokoki oraz gonokoki i meningokoki w kształcie fasoli; paciorkowce (od łac. streptos - łańcuch) - komórki dzielą się w jednej płaszczyźnie i nie rozchodzą się, tworząc łańcuch; gronkowce (od łac. gronkowiec - kiść) - komórki dzielą się w różnych płaszczyznach, tworząc skupiska w postaci kiści winogron; tetracocci (od łac. tetra - cztery) - komórki dzielą się w dwóch wzajemnie prostopadłych płaszczyznach i układają się w cztery; sarcins (od łac. sarcio – łączyć) – komórki podzielone są na trzy wzajemnie prostopadłe płaszczyzny i ułożone są w bele lub paczki po 8 lub 16 komórek.
Cocci są szeroko rozpowszechnione w środowisku, a także u ludzi i zwierząt. Prawie wszystkie grupy ziarniaków, z wyjątkiem mikrokoków, tetrakoków i sarcyn, obejmują patogeny choroba zakaźna.
formy w kształcie prętów zwane bakteriami. Ich średnie wymiary wynoszą od 1 do 6 mikronów długości i od 0,5 do 2 mikronów grubości.
Bakterie różnią się wygląd: ich końce mogą być zaokrąglone (E. coli), odcięte (patogen wąglik), spiczasty (czynnik sprawczy dżumy) lub pogrubiony (czynnik sprawczy błonicy). Po podziale bakterie mogą być ułożone parami - diplobacteria (Klebsiella), w łańcuchu (czynnik sprawczy wąglika), czasem pod kątem względem siebie lub poprzecznie (czynnik sprawczy błonicy). Większość bakterii jest rozmieszczona losowo.
Wśród bakterii występują zakrzywione formy - vibrios (czynnik sprawczy cholery).
Formy zawiłe obejmują spirillę i krętki. Kształt ich komórki przypomina spiralę. Większość spirillae nie jest chorobotwórcza.
Struktura komórki bakteryjnej
Aby zbadać strukturę komórki bakteryjnej, wraz z mikroskopem świetlnym, stosuje się badania mikroskopii elektronowej i mikrochemicznej w celu określenia ultrastruktury komórki bakteryjnej.
Komórka bakteryjna (ryc. 5) składa się z następujących części: błony trójwarstwowej, cytoplazmy z różnymi inkluzjami oraz substancji jądrowej (nukleoidu). Dodatkowe formacje strukturalne to kapsułki, zarodniki, wici, pilusy.
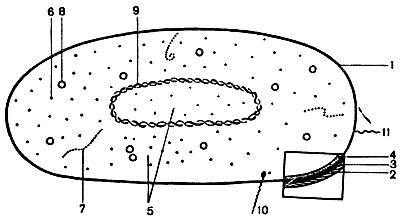
Powłoka Komórka składa się z zewnętrznej warstwy błony śluzowej, ściany komórkowej i błona cytoplazmatyczna.
Warstwa torebki śluzowej znajduje się na zewnątrz komórki i pełni funkcję ochronną.
Ściana komórkowa jest jednym z głównych elementów strukturalnych komórki, utrzymującym jej kształt i oddzielającym komórkę od środowisko. Ważna właściwośćŚciana komórkowa jest selektywnie przepuszczalna, co zapewnia przenikanie niezbędnych składników odżywczych (aminokwasów, węglowodanów itp.) do wnętrza komórki i usuwanie produktów przemiany materii z komórki. Ściana komórkowa utrzymuje stałe ciśnienie osmotyczne wewnątrz komórki. Wytrzymałość ściany zapewnia mureina, substancja o charakterze polisacharydowym. Niektóre substancje niszczą ścianę komórkową, na przykład lizozym.
Bakterie całkowicie pozbawione ściany komórkowej nazywane są protoplasty. Zachowują zdolność oddychania, podziału, syntezy enzymów; na wpływ czynników zewnętrznych: uszkodzenie mechaniczne, ciśnienie osmotyczne, napowietrzanie itp. Protoplasty można konserwować tylko w roztworach hipertonicznych.
Bakterie z częściowo zniszczoną ścianą komórkową nazywane są sferoplastami. Jeśli hamujesz proces syntezy ściany komórkowej za pomocą penicyliny, powstają formy L, które we wszystkich typach bakterii są kulistymi dużymi i małymi komórkami z wakuolami.
Błona cytoplazmatyczna ściśle przylega do ściany komórkowej wewnątrz. Jest bardzo cienka (8-10 nm) i składa się z białek i fosfolipidów. Jest to półprzepuszczalna warstwa graniczna, przez którą komórka jest odżywiana. Błona zawiera enzymy permeazy, które przeprowadzają aktywny transport substancji oraz enzymy oddechowe. Błona cytoplazmatyczna tworzy mezosomy, które biorą udział w podziale komórki. Kiedy komórkę umieszcza się w roztworze hipertonicznym, błona może oddzielić się od ściany komórkowej.
Cytoplazma- wnętrze komórki bakteryjnej. Jest to układ koloidalny składający się z wody, białek, węglowodanów, lipidów, różnych sole mineralne. Skład chemiczny i konsystencja cytoplazmy zmieniają się w zależności od wieku komórki i warunków środowiskowych. Cytoplazma zawiera substancję jądrową, rybosomy i różne inkluzje.
Nukleoid, substancja jądrowa komórki, jej aparat dziedziczny. Substancja jądrowa prokariotów, w przeciwieństwie do eukariontów, nie ma własnej błony. Nukleoid dojrzałej komórki to podwójna nić DNA zwinięta w pierścień. Cząsteczka DNA koduje informację genetyczną komórki. Zgodnie z terminologią genetyczną substancja jądrowa nazywana jest genoforem lub genomem.
Rybosomy znajdują się w cytoplazmie komórki i pełnią funkcję syntezy białek. Rybosom zawiera 60% RNA i 40% białka. Liczba rybosomów w komórce sięga 10 000. Łącząc się, rybosomy tworzą polisomy.
Inkluzje - granulki zawierające różne zapasy składniki odżywcze: skrobia, glikogen, tłuszcz, wolutyna. Znajdują się w cytoplazmie.
Komórki bakteryjne w procesie życia tworzą organelle ochronne - otoczki i zarodniki.
Kapsuła- zewnętrzna zbita warstwa śluzowa przylegająca do ściany komórkowej. Jest to narząd ochronny, który pojawia się u niektórych bakterii, gdy dostają się one do organizmu ludzi i zwierząt. Kapsułka chroni mikroorganizm przed czynnikami ochronnymi organizmu (czynniki sprawcze zapalenia płuc i wąglika). Niektóre mikroorganizmy mają trwałą torebkę (Klebsiella).
spór występuje tylko u bakterii o kształcie pręcików. Powstają, gdy mikroorganizm wejdzie w niekorzystne warunki. otoczenie zewnętrzne(działanie wysokie temperatury, wysychanie, zmiana pH, spadek ilości składników pokarmowych w pożywce itp.). Zarodniki znajdują się wewnątrz komórki bakteryjnej i reprezentują zwarty obszar cytoplazmy z nukleoidem, ubrany we własny gęsta skorupa. Przez skład chemiczny różnią się od komórek wegetatywnych niewielką ilością wody, zwiększoną zawartością lipidów i soli wapnia, co przyczynia się do dużej odporności przetrwalników. Zarodnikowanie następuje w ciągu 18-20 godzin; gdy mikroorganizm wejdzie w sprzyjające warunki, zarodnik kiełkuje do postaci wegetatywnej w ciągu 4-5 godzin. W komórce bakteryjnej powstaje tylko jeden zarodnik, dlatego zarodniki nie są narządami rozrodczymi, ale służą do przetrwania niekorzystnych warunków.
Kształtowanie zarodników bakterie tlenowe nazywane są pałeczkami, a beztlenowe - Clostridia.
Zarodniki różnią się kształtem, rozmiarem i umiejscowieniem w komórce. Mogą być zlokalizowane centralnie, podkońcowo i końcowo (ryc. 6). W czynniku sprawczym wąglika zarodnik znajduje się centralnie, jego rozmiar nie przekracza średnicy komórki. Zarodnik czynnika sprawczego zatrucia jadem kiełbasianym znajduje się bliżej końca komórki - podterminalnie i przekracza szerokość komórki. W czynniku wywołującym tężec zaokrąglony zarodnik znajduje się na końcu komórki - na końcu i znacznie przekracza szerokość komórki.
wici- narządy ruchu charakterystyczne dla bakterii pałeczkowatych. Są to cienkie włókna nitkowate, składające się z białka - flageliny. Ich długość znacznie przekracza długość komórki bakteryjnej. Wici rozciągają się od ciała podstawowego znajdującego się w cytoplazmie i wychodzą na powierzchnię komórki. Ich obecność można wykryć, określając ruchliwość komórek pod mikroskopem, w półpłynnej pożywce lub barwiąc specjalnymi metodami. Ultrastrukturę wici badano za pomocą mikroskopu elektronowego. W zależności od lokalizacji wici bakterie dzielą się na grupy (patrz ryc. 6): monotrichous - z jedną wicią (czynnik sprawczy cholery); amphitrichous - z wiązkami lub pojedynczymi wiciami na obu końcach komórki (spirilla); lophotrichous - z wiązką wici na jednym końcu komórki (kał alkaliczny); peritrichous - wici znajdują się na całej powierzchni komórki ( bakterie jelitowe). Szybkość przemieszczania się bakterii zależy od liczby i umiejscowienia wici (najbardziej aktywne są wici), od wieku bakterii oraz wpływu czynników środowiskowych.
Pili lub fimbrie- kosmki znajdujące się na powierzchni komórek bakteryjnych. Są krótsze i cieńsze niż wici, a także mają strukturę spiralną. Składa się z picia z białka - pilinu. Niektóre pilusy (jest ich kilkaset) służą do przyczepiania się bakterii do komórek zwierzęcych i ludzkich, inne (pojedyncze) są związane z przenoszeniem materiału genetycznego z komórki do komórki.
Mykoplazmy
Mykoplazmy to komórki, które nie mają ściany komórkowej, ale są otoczone trójwarstwową lipoproteinową błoną cytoplazmatyczną. Mykoplazmy mogą być kuliste, owalny kształt, w postaci nici i gwiazdek. Mykoplazmy zgodnie z klasyfikacją Bergiego są izolowane w oddzielna grupa. Obecnie drobnoustrojom tym poświęca się coraz więcej uwagi jako czynnikom wywołującym choroby zapalne. Ich rozmiary są różne: od kilku mikrometrów do 125-150 nm. Małe mykoplazmy przechodzą przez filtry bakteryjne i nazywane są formami filtrowalnymi.
Krętki
Krętki (patrz ryc. 52) (z łac. speira - zgięcie, chaite - włosy) - cienkie, skręcone, ruchliwe organizmy jednokomórkowe, mierzące od 5 do 500 mikronów długości i 0,3-0,75 mikronów szerokości. Z najprostszymi łączy się je metodą ruchu poprzez skrócenie wewnętrznego gwintu osiowego, składającego się z wiązki włókienek. Charakter ruchu krętków jest inny: translacyjny, obrotowy, zgięciowy, falisty. Reszta struktury komórkowej jest typowa dla bakterii. Niektóre krętki słabo barwią się barwnikami anilinowymi. Krętki dzieli się na rodzaje ze względu na liczbę i kształt skrętów nici oraz jej zakończenie. Oprócz form saprofitycznych, powszechnych w przyrodzie i organizmie człowieka, wśród krętków występują patogeny - czynniki wywołujące kiłę i inne choroby.
Rickettsia
Wirusy
Wśród wirusów wyróżnia się grupę fagów (łac. phagos – pożerających), powodujących lizę (zniszczenie) komórek mikroorganizmów. Zachowując właściwości i skład charakterystyczny dla wirusów, fagi różnią się budową wirionu (patrz rozdział 8). Nie powodują chorób u ludzi i zwierząt.
Pytania kontrolne
1. Opowiedz nam o klasyfikacji mikroorganizmów.
2. Jakie są główne właściwości przedstawicieli królestwa prokariotów.
3. Wymień i scharakteryzuj główne formy bakterii.
4. Wymień główne organelle komórkowe i ich przeznaczenie.
5. Podaj krótki opis głównych grup bakterii i wirusów.
Badanie morfologii mikroorganizmów
Do badania morfologii mikroorganizmów stosuje się metodę badań mikroskopowych. Ważny warunek pomyślne użycie ta metoda jest właściwe gotowanie rozmaz z badanego materiału lub hodowli bakteryjnej. Kultura odnosi się do mikroorganizmów hodowanych na pożywce w laboratorium.
Technika przygotowania rozmazu
Do pracy niezbędne jest posiadanie czystych i odtłuszczonych szkiełek i szkiełek nakrywkowych. Nowe szklanki gotuje się przez 15-20 minut w 2-5% roztworze sody lub wody z mydłem, spłukuje wodą i umieszcza w słabym kwasie solnym, a następnie dokładnie myje wodą.
Szklanki używane i zanieczyszczone barwnikami lub olejkiem immersyjnym można potraktować na dwa sposoby: 1) zanurzyć na 2 godziny w stężonym Kwas Siarkowy lub mieszaniną chromu, a następnie dokładnie spłukać; 2) gotować przez 30-40 minut w 5% roztworze sody lub zasady. Surowe szkło można odtłuścić przecierając je mydłem, a następnie czyszcząc suchą szmatką.
Uwaga! Jeśli szkło jest dobrze odtłuszczone, kropla wody rozprowadza się na nim równomiernie, nie rozpadając się na małe krople.
Szklanki przechowuje się w naczyniach ze szlifowanymi korkami w mieszaninie Nikiforowa (równe ilości alkoholu i eteru) lub w 96% alkoholu. Szklane roztwory usuwa się pęsetą.
Uwaga! Podczas pracy szkło należy trzymać palcami za krawędzie.
Materiał do badań nanosi się na szkiełko za pomocą ezy bakteryjnej, igły lub pipety Pasteura. Najczęściej stosuje się pętlę bakteryjną (ryc. 7), wykonaną z nici platynowej lub nichromowej o długości 5-6 cm, pętlę mocuje się w uchwycie pętli lub wlutowuje w szklany pręt. Koniec drutu jest zagięty w kształcie pierścienia o wymiarach 1×1,5 lub 2×3 µm.
Uwaga! Odpowiednio przygotowana pętla po zanurzeniu w wodzie i wyjęciu z niej zachowuje film wodny.
Przed przygotowaniem rozmazu część robocza pętli jest spalana w płomieniu palnika pozycja pionowa: najpierw sama pętla, a następnie metalowy pręt. Ta manipulacja jest przeprowadzana po zakończeniu siewu.
Przygotowanie rozmazu z kultury wyhodowanej na płynnej pożywce. Beztłuszczowe szkiełko wypala się w płomieniu palnika i chłodzi. Hodowlę nanosi się na szkiełko umieszczone na statywie (szalka Petriego, statyw). Probówka hodowlana jest duża i palce wskazujące lewa ręka. Pętla jest trzymana w prawej ręce. Bez zwalniania pętli, mały palec prawa ręka dociśnij korek do dłoni i ostrożnie wyjmij go z probówki. Ruchy powinny być płynne i spokojne. Gardło rury jest spalane w płomieniu palnika. Włóż pętlę do probówki. Ochłodzić pętlę przy ściance probówki, a następnie zanurzyć ją w hodowli. Usuń pętlę bez dotykania ścian rurki. Zamknąć korek po przepuszczeniu go przez płomień palnika. Umieść probówkę na statywie. Hodowlę nanosi się ezą na szkiełku podstawowym, ruchem okrężnym rozprowadzając go równomiernie. Następnie pętla jest spalana w płomieniu palnika. Rozmaz pozostawia się do wyschnięcia.
Uwaga! Rozmaz powinien być równomiernie rozprowadzony, cienki i mały (mniej więcej wielkości monety dwukopiowej).
Przygotowanie rozmazu z kultury wyhodowanej na pożywce stałej. Kroplę izotonicznego roztworu chlorku sodu (0,9%) nanosi się na przygotowane szkiełko za pomocą pipety Pasteura lub ezy. Hodowlę ostrożnie usuwa się ezą z agaru w probówce lub szalce Petriego i emulguje w kropli na szkle. Przygotowany rozmaz powinien być jednolity i niezbyt gruby. Po wyschnięciu na szklanym szkiełku pozostaje niewielka powłoka.
Przygotowanie rozmazu z ropy lub plwociny. Materiał pobiera się sterylną pipetą lub ezą i nakłada na środek szkiełka podstawowego. Przykryj pierwszy slajd drugim slajdem, tak aby jedna trzecia pierwszego i drugiego slajdu pozostała wolna. Okulary rozsuwają się z wysiłkiem. Przygotuj dwa duże waciki.
Przygotowanie rozmazu krwi. Kroplę krwi nanosi się na szkiełko w odległości jednej trzeciej od lewej krawędzi. Następnie krawędź specjalnie wypolerowanego szkła, nachylonego pod kątem 45°, dotyka kropli krwi. Dociskając polerowane szkło do przedmiotu, przesuwają je do przodu. Odpowiednio przygotowany rozmaz ma żółtawy kolor i prześwituje.
Sporządzanie rozmazów-odcisków z organów wewnętrznych zwłok oraz produktów spożywczych o stałej konsystencji. powierzchni narządu lub produkt spożywczy przyżega się gorącym skalpelem i wycina się z tego obszaru kawałek materiału. Kawałek ten ostrożnie chwyta się pęsetą, a wyciętą powierzchnię dotyka szkiełka w dwóch lub trzech miejscach, wykonując serię pociągnięć-odcisków.
Suszenie rozmazu
Rozmaz suszy się na powietrzu w temp temperatura pokojowa. W razie potrzeby można go wysuszyć w pobliżu płomienia palnika, trzymając szklankę pozycja pozioma przez krawędzie kciukiem i palcem wskazującym, pociągnij w górę.
Uwaga! W wysokich temperaturach może dojść do uszkodzenia struktury komórek.
Utrwalanie rozmazu
Smugi utrwala się po całkowitym wyschnięciu w celu: 1) utrwalenia mikroorganizmów na szkle; 2) zneutralizować materiał; 3) zabite mikroorganizmy lepiej postrzegają kolor. Utrwalony wacik nazywany jest preparatem.
metody mocowania. 1. Fizyczny - w płomieniu palnika: szkło jest pobierane pęsetą lub kciukiem i palcem wskazującym i przepuszczane przez trzy razy Górna część płomień palnika przez 6 s.
2. Chemiczny - w płynie: elementy komórkowe w rozmazach krwi i rozmazach - odciski pod wpływem wysokich temperatur ulegają zniszczeniu, dlatego traktuje się je jednym z płynów utrwalających: a) alkoholem metylowym - 5 min; B) alkohol etylowy- 10 minut; c) mieszanka Nikiforowa - 10-15 minut; d) aceton - 5 minut; e) opary kwasu i formaliny - kilka sekund.
Preparaty barwiące
Po utrwaleniu rozpoczyna się barwienie preparatu.
Preparaty są barwione na specjalnie wyposażonym stole pokrytym linoleum, plastikiem, szkłem itp. Na stole potrzebne jest naczynie z wodą destylowaną; stojak z dwóch rurek lub patyków połączonych gumowymi rurkami z obu stron (do umieszczania preparatów); pęsety, cylindry, pipety, bibuła filtracyjna, zestaw barwników, pojemnik do ich odwadniania. Stół do malowania powinien znajdować się w pobliżu kranu z wodą.
Stosunek mikroorganizmów do barwników nazywany jest ich właściwościami barwiącymi. Barwniki anilinowe są szeroko stosowane w mikrobiologii. Większość mikroorganizmów lepiej postrzega podstawowe barwniki.
Najczęściej stosowane są następujące barwniki: czerwony (podstawowa magenta, kwaśna magenta, czerwień Kongo, czerwień neutralna); niebieski (metylen i toluidyna); fiolet (goryczka, metyl, krystaliczny); brązowo-żółty (vesuvin, chrysoidin); zielony (diament, malachit).
Wszystkie barwniki produkowane są w postaci amorficznych lub krystalicznych proszków. Przygotowuje się z nich nasycone roztwory alkoholowe i fenolowe, a następnie do pracy stosuje się wodno-alkoholowe lub wodno-fenolowe roztwory barwników. Jeśli do barwienia używa się stężonych roztworów barwników, wówczas preparat jest wstępnie przykrywany bibułą filtracyjną, na którą nakłada się barwnik. W takim przypadku kawałki barwnika pozostają na papierze.
Uwaga! Kroplę barwnika nakładamy pipetą tak, aby pokryła cały preparat.
Przepisy na barwniki
1. Nasycone roztwory alkoholowe (początkowe):
Barwnik - 1 g alkoholu 96% - 10 ml
Mieszaninę umieszcza się w termostacie do całkowitego rozpuszczenia na kilka dni. Wstrząsnąć codziennie. Przechowywane w butelkach ze szlifowanymi korkami.
2. Carbol fuksyna Ziel (do barwienia mikroorganizmów kwasoodpornych, zarodników i kapsułek):
Nasycony alkoholowy roztwór fuksyny zasadowej - 10 ml roztworu kwasu karbolowego 5% - 90 ml
Uwaga! Kwas karbolowy wlewa się do barwnika, a nie odwrotnie.
Mieszaninę wytrząsa się energicznie przez kilka minut, przesącza i wlewa do fiolki do przechowywania.
3. Pfeiffer magenta (do barwienia metodą Grama i metodą prostą):
Fuchsina Tsilya - 1 ml wody destylowanej - 9 ml
Barwnik jest przygotowywany bezpośrednio przed użyciem.
4. Fiolet goryczki karbolowej (do barwienia metodą Grama):
nasycony roztwór alkoholu
fiołek goryczki - 10 ml
kwas karbolowy 5% - 100 ml
Roztwory miesza się i filtruje przez bibułę filtracyjną.
5. Roztwór Lugola (do barwienia Grama i odczynnika skrobiowego):
Jodek potasu - 2 g jodu krystalicznego - 1 g wody destylowanej - 10 ml
Mieszaninę umieszcza się w matowej szklanej butelce, dobrze zakorkuje i umieszcza w termostacie na jeden dzień, następnie dodaje 300 ml wody destylowanej.
6. Roztwór alkaliczny błękit metylenowy Lefflera:
Nasycony alkoholowy roztwór błękitu metylenowego - 30 ml 1% roztworu wodorotlenku potasu - 1 ml wody destylowanej - 100 ml
7. Papiery według Sineva (do barwienia metodą Grama):
1% roztwór alkoholu krystaliczny fiolet
Paski bibuły filtracyjnej moczy się w roztworze i suszy.
Metody barwienia dzielą się na orientacyjne (proste) i różnicowe (złożone), ujawniające cechy chemiczne i strukturalne komórki bakteryjnej.
Prosta metoda barwienia
Lek umieszcza się na stojaku do barwienia, materiałem testowym do góry. Za pomocą pipety nakłada się na nią roztwór barwnika. Po określonym czasie barwnik ostrożnie odsącza się, preparat przemywa wodą i suszy bibułą filtracyjną. Na prosta metoda używany jest jeden barwnik. Błękit metylenowy i błękit alkaliczny Leffler barwią preparat przez 3-5 minut, fuksyna Pfeiffera - 1-2 minuty (patrz ryc. 4).
Kroplę olejku immersyjnego nanosi się na wybarwiony i wysuszony preparat i
Wyrafinowane metody barwienia
barwienie metodą Grama ( metoda uniwersalna) . Najpopularniejszą metodą barwienia różnicowego jest barwienie metodą Grama.
W zależności od wyników barwienia wszystkie mikroorganizmy dzielą się na dwie grupy - Gram-dodatnie i Gram-ujemne.
Bakterie Gram-dodatnie zawierają w ścianie komórkowej sól magnezową RNA, która tworzy związek kompleksowy z jodem i głównym barwnikiem (goryczką, metylem lub fioletem krystalicznym). Kompleks ten nie ulega zniszczeniu pod wpływem alkoholu, a bakterie zachowują swój fioletowy kolor.
Bakterie Gram-ujemne nie są w stanie zatrzymać podstawowego barwnika, ponieważ nie zawierają soli magnezowej RNA. Pod wpływem alkoholu barwnik ulega wypłukaniu, komórki ulegają odbarwieniu i zabarwieniu dodatkowym barwnikiem (magenta) czerwonym.
1. Na preparat według Sineva nakłada się kartkę papieru i nanosi się kilka kropli wody lub roztwór fioletu goryczki. Bejca 1-2 min. Usuń papier lub osusz barwnik.
2. Bez przemywania wodą nanosi się płyn Lugola, aż zmieni kolor na czarny (1 min), a następnie odsącza się barwnik.
3. Bez spłukiwania wodą nakłada się 96% alkohol do momentu zniknięcia barwnika (30-60 s). Możesz obniżyć lek do szklanki alkoholu na 1-2 sekundy.
4. Zmyć preparat wodą.
5. Barwiono magentą Pfeiffer przez 3 minuty, przemywano wodą i suszono.
Mikroskopowo za pomocą systemu zanurzeniowego.
Barwienie metodą Ziehla-Nielsena (dla bakterii kwasoodpornych). Ta metoda służy do wykrywania bakterii gruźlicy i trądu, które mają błony komórkowe duża liczba lipidy, woski i hydroksykwasy. Bakterie są odporne na kwasy, zasady i alkohole. Aby zwiększyć przepuszczalność ściany komórkowej, pierwszy etap barwienia przeprowadza się z ogrzewaniem.
1. Utrwalony preparat przykrywamy bibułą filtracyjną i nakładamy fuksynę Ziela. Trzymając szklankę pęsetą, preparat podgrzewa się nad płomieniem palnika do momentu ulotnienia się oparów. Dodaj nową porcję barwnika i podgrzej jeszcze 2 razy. Po ochłodzeniu bibułę usuwa się i preparat przemywa wodą.
2. Lek odbarwia się 5% roztworem kwasu siarkowego, zanurza 2-3 razy w roztworze lub wylewa kwas na szkło, a następnie kilkakrotnie przemywa wodą.
3. Barwiono wodno-alkoholowym roztworem błękitu metylenowego przez 3-5 minut, przemywano wodą i suszono.
Mikroskopowo za pomocą systemu zanurzeniowego.
Bakterie kwasoodporne są zabarwione na czerwono, reszta na niebiesko (patrz ryc. 4).
Barwienie wg Orzeszki (identyfikacja zarodników). 1. Kilka kropli 0,5% roztworu kwasu chlorowodorowego wlewa się na wysuszoną na powietrzu rozmaz i ogrzewa do powstania oparów. Lek suszy się i utrwala nad płomieniem.
2. Barwione metodą Ziehla-Nielsena. Zarodniki kwasoodporne są zabarwione na różowo-czerwono, a komórka bakteryjna na niebiesko (patrz ryc. 4).
Barwienie Burri-Ginsu (wykrywanie kapsułki). Ta metoda nazywana jest negatywną, ponieważ tło leku i komórka bakteryjna są barwione, podczas gdy kapsułka pozostaje niezabarwiona.
1. Kropla czarnego atramentu, rozcieńczona 10-krotnie, jest nakładana na szkiełko. Dodają do tego trochę kultury. Rozmaz robi się krawędzią mielącego szkła, podobnie jak rozmaz krwi, i suszy.
2. Napraw środkami chemicznymi alkohol lub sublimacja. Ostrożnie spłukać wodą.
3. Barwić Pfeiffer magenta przez 3-5 minut. Umyć dokładnie i wysuszyć na powietrzu.
Uwaga! Nie stosować bibuły filtracyjnej, aby nie uszkodzić preparatu.
Mikroskopowo za pomocą systemu zanurzeniowego. Tło leku jest czarne, komórki są czerwone, kapsułki są niebarwione (patrz ryc. 4).
Żywotne barwienie mikroorganizmów
Do badania żywej kultury najczęściej stosuje się błękit metylenowy i inne barwniki w dużych rozcieńczeniach (1: 10 000). Kroplę badanego materiału miesza się na szklanym szkiełku z kroplą barwnika i przykrywa szkiełkiem nakrywkowym. Mikroskopowo z obiektywem 40x.
Badanie mobilności mikroorganizmów
Do badań kultura bakterii hodowana w płynnej pożywce lub zawiesinie bakterii w roztwór izotoniczny chlorek sodu.
metoda zgniecionej kropli. Umieścić kroplę kultury na szkiełku podstawowym i przykryć szkiełkiem nakrywkowym. Aby uniknąć tworzenia się pęcherzyków powietrza, szkiełko nakrywkowe jest doprowadzane krawędzią do krawędzi kropli i ostro opuszczane. Aby chronić lek przed wysychaniem, umieszcza się go w wilgotnej komorze.
Komora mokra to szalka Petriego z wilgotną bibułą filtracyjną na dnie. Na papierze umieszcza się dwie zapałki i umieszcza się na nich lek. Kubek zamykany jest pokrywką.
Mikroskopijne przy 40-krotnym powiększeniu obiektywu w ciemnym polu (patrz rozdział 2).
metoda wiszącej kropli(Rys. 8). Do przygotowania leku potrzebna jest szklanka z dziurką, szkiełko nakrywkowe i wazelina. Krawędzie otworu są pokryte cienką warstwą wazeliny.

Kroplę kultury nanosi się na szkiełko nakrywkowe. Następnie ostrożnie przykryj szkiełko nakrywkowe szklanką z otworem, tak aby kropla znajdowała się na środku. Lepkie szkiełka są szybko odwracane z uniesionym szkiełkiem nakrywkowym. Kropla znajduje się w hermetycznej komorze i jest przechowywana przez długi czas. Pod mikroskopem najpierw przy małym powiększeniu (8x) znajduje się krawędź kropli, a następnie preparat bada się przy dużym powiększeniu.
Pytania kontrolne
1. Jak przygotować pętlę bakteryjną?
2. Wymień cele i metody utrwalania rozmazów.
3. Wymień główne barwniki.
4. Jakie metody badają ruchliwość mikroorganizmów?
Ćwiczenia
1. Weź gotowe preparaty, zbadaj je i narysuj główne formy mikroorganizmów.
2. Przygotuj wymazy z inny materiał(posiewy, ropa, krew, rozmazy-odciski).
3. Preparaty do plam złożone metody(według Grama, Tsil - Nielsen, Ozheshko, Burri - Guins).
bakteria- to jest bardzo prosta formażycie roślinne, które składa się z pojedynczej żywej komórki. Rozmnażanie odbywa się przez podział komórki. Po osiągnięciu dojrzałości bakteria podzielony na dwa równe komórki. Z kolei każda z tych komórek osiąga dojrzałość i również dzieli się na dwie równe komórki. W idealne warunki bakteria osiąga stan dojrzałości i namnaża się w mniej niż 20–30 minut. Przy takim tempie rozmnażania jedna bakteria mogłaby teoretycznie wyprodukować 34 biliony potomstwa w ciągu 24 godzin! Na szczęście, koło życia bakterii jest stosunkowo krótki i trwa od kilku minut do kilku godzin. Dlatego nawet w idealnych warunkach nie mogą rozmnażać się w takim tempie.tempo wzrostu i hodowla bakterii i innych mikroorganizmów zależy od warunków środowiskowych. Temperatura, światło, tlen, wilgotność i pH (kwasowość lub zasadowość), wraz z dostępnością pożywienia, wpływają na tempo wzrostu bakterii. Spośród nich temperatura jest przedmiotem szczególnego zainteresowania techników i inżynierów. Dla każdej odmiany bakterii istnieje minimalna temperatura, w której mogą rosnąć. W temperaturach poniżej tego progu bakterie hibernują i nie są w stanie się rozmnażać. Dokładnie to samo dla każdego odmiany bakterii istnieje maksymalny próg temperatury. W temperaturach powyżej tej granicy bakterie ulegają zniszczeniu. Pomiędzy tymi granicami znajduje się optymalna temperatura, w której bakterie namnażają się z maksymalną szybkością. Optymalna temperatura dla większości bakterii żywiących się odchodami zwierzęcymi oraz martwymi tkankami zwierząt i roślin (saprofitami), od 24 do 30°C. Optymalna temperatura dla większości bakterii wywołujących infekcje i choroby żywiciela (bakterie chorobotwórcze) wynosi około 38°C. W większości przypadków możliwe jest znaczne zmniejszenie tempo wzrostu bakterii jeśli temperatura otoczenia jest obniżona. Wreszcie, istnieje kilka odmian bakterii, które najlepiej rozwijają się w temperaturze wody, podczas gdy inne najlepiej radzą sobie w temperaturach ujemnych.
Dodatek do powyższego
Pochodzenie, ewolucja, miejsce w rozwoju życia na Ziemi
Bakterie, obok archeonów, były jednymi z pierwszych żywych organizmów na Ziemi, pojawiły się około 3,9-3,5 miliarda lat temu. Relacje ewolucyjne między tymi grupami nie zostały jeszcze w pełni zbadane, istnieją co najmniej trzy główne hipotezy: N. Pace sugeruje, że mają wspólnego przodka protobakterii, Zavarzin uważa archeony za ślepą uliczkę ewolucji eubakterii, która opanowała ekstremalne siedliska; wreszcie, zgodnie z trzecią hipotezą, archeony są pierwszymi żywymi organizmami, z których wywodzą się bakterie.Eukarionty powstały w wyniku symbiogenezy z komórek bakteryjnych znacznie później: około 1,9-1,3 miliarda lat temu. Ewolucja bakterii charakteryzuje się wyraźnym odchyleniem fizjologicznym i biochemicznym: przy względnym ubóstwie form życia i prymitywnej strukturze opanowały one prawie wszystkie znane obecnie procesy biochemiczne. Biosfera prokariotyczna posiadała już wszystkie istniejące obecnie sposoby przemian substancji. Eukarionty, wnikając w nią, zmieniły tylko ilościowe aspekty swojego funkcjonowania, ale nie jakościowe; na wielu etapach pierwiastków bakterie nadal zachowują pozycję monopolistyczną.
Jedną z najstarszych bakterii są sinice. W skałach powstałych 3,5 miliarda lat temu znaleziono produkty ich życiowej aktywności, stromatolity; niepodważalne dowody na istnienie sinic pochodzą sprzed 2,2–2,0 miliarda lat. Dzięki nim w atmosferze zaczął gromadzić się tlen, który 2 miliardy lat temu osiągnął stężenia wystarczające do rozpoczęcia oddychania tlenowego. Do tego czasu należą formacje charakterystyczne dla bezwzględnie tlenowego Metallogenium.
Pojawienie się tlenu w atmosferze zadało poważny cios bakteriom beztlenowym. Albo wymierają, albo przechodzą do lokalnie zachowanych stref beztlenowych. Całkowita różnorodność gatunkowa bakterii w tym czasie jest zmniejszona.
Przyjmuje się, że ze względu na brak procesu płciowego ewolucja bakterii przebiega według zupełnie innego mechanizmu niż u eukariontów. Stały poziomy transfer genów prowadzi do niejasności w obrazie relacji ewolucyjnych, ewolucja przebiega niezwykle wolno (i być może wraz z pojawieniem się eukariontów całkowicie się zatrzymała), ale w zmieniających się warunkach następuje szybka redystrybucja genów między komórkami z niezmienionym wspólna pula genetyczna.
Struktura
Zdecydowana większość bakterii (z wyjątkiem promieniowców i sinic nitkowatych) to bakterie jednokomórkowe. W zależności od kształtu komórek mogą być okrągłe (cocci), w kształcie pręta (bacilli, clostridia, pseudomonads), zawiłe (vibrios, spirilla, krętki), rzadziej - gwiaździste, czworościenne, sześcienne, C- lub O- ukształtowany. Kształt determinuje takie zdolności bakterii jak przywieranie do powierzchni, ruchliwość, wchłanianie składników odżywczych. Zauważono np., że oligotrofy, czyli bakterie żyjące w środowisku o niskiej zawartości składników odżywczych, mają tendencję do zwiększania stosunku powierzchni do objętości, np. poprzez tworzenie wypustek (tzw. ).Z obowiązkowych struktury komórkowe wyróżnić trzy:
- nukleoid
- rybosomy
- błona cytoplazmatyczna (CPM)
Struktura protoplastu
CPM ogranicza zawartość komórki (cytoplazmy) przed środowiskiem zewnętrznym. Jednorodna frakcja cytoplazmy, zawierająca zestaw rozpuszczalnych RNA, białek, produktów i substratów reakcji metabolicznych, nazywana jest cytozolem. Inna część cytoplazmy jest reprezentowana przez różne elementy strukturalne.Jedną z głównych różnic między komórką bakteryjną a komórką eukariotyczną jest brak błony jądrowej i, ściśle mówiąc, brak jakichkolwiek błon wewnątrzcytoplazmatycznych, które nie są pochodnymi CPM. Jednakże, różne grupy prokarionty (szczególnie często u bakterii Gram-dodatnich) mają miejscowe wypustki CPM – mezosomy, które pełnią różne funkcje w komórce i dzielą ją na funkcjonalnie różne części. Wiele bakterii fotosyntetyzujących ma rozwiniętą sieć błon fotosyntetycznych pochodzących z CPM. Fioletowe bakterie zachowały swój związek z CPM, co można łatwo wykryć na sekcjach poniżej mikroskop elektronowy u sinic związek ten jest albo trudny do wykrycia, albo zatracony w toku ewolucji. W zależności od warunków i wieku kultury błony fotosyntetyczne tworzą różne struktury - pęcherzyki, chromatofory, tylakoidy.
Cała informacja genetyczna niezbędna do życia bakterii zawarta jest w jednym DNA (chromosomie bakteryjnym), najczęściej w postaci kowalencyjnie zamkniętego pierścienia (chromosomy liniowe występują u Streptomyces i Borrelia). Jest przyłączony do CPM w jednym punkcie i jest umieszczony w strukturze, która jest izolowana, ale nie jest oddzielona błoną od cytoplazmy i nazywana jest nukleoidem. Rozłożony DNA ma ponad 1 mm długości. Chromosom bakteryjny jest zwykle prezentowany w jednej kopii, to znaczy prawie wszystkie prokarioty są haploidalne, chociaż w pewnych warunkach jedna komórka może zawierać kilka kopii swojego chromosomu, a Burkholderia cepacia ma trzy różne chromosomy pierścieniowe (3,6; 3,2 i 1,1 miliona długości) ).pary zasad). Rybosomy prokariotów również różnią się od rybosomów eukariontów i mają stałą sedymentacji 70 S (80 S u eukariontów).
Oprócz tych struktur w cytoplazmie można znaleźć również inkluzje substancji zapasowych.
Ściana komórkowa i struktury powierzchniowe
Ściana komórkowa jest ważna element konstrukcyjny komórka bakteryjna, jednak opcjonalnie. Sztucznie otrzymano formy z częściowo lub całkowicie nieobecną ścianą komórkową (formy L), które mogły istnieć w sprzyjających warunkach, ale czasami traciły zdolność do podziału. Istnieje również grupa naturalnych bakterii, które nie posiadają ściany komórkowej – mykoplazmy.W bakteriach istnieją dwa główne typy struktury ściany komórkowej, charakterystyczne dla gatunków Gram-dodatnich i Gram-ujemnych.
Ściana komórkowa bakterii Gram-dodatnich jest jednorodną warstwą o grubości 20-80 nm, zbudowaną głównie z peptydoglikanu z mniejszą ilością kwasów teichojowych oraz niewielką ilością polisacharydów, białek i lipidów (tzw. lipopolisacharyd). Ściana komórkowa ma pory o średnicy 1–6 nm, dzięki czemu jest przepuszczalna dla wielu cząsteczek.
U bakterii Gram-ujemnych warstwa peptydoglikanu nie przylega ściśle do CPM i ma grubość zaledwie 2–3 nm. Jest otoczony zewnętrzną membraną, która z reguły ma nierówny, zakrzywiony kształt. Pomiędzy CPM, warstwą peptydoglikanu i błoną zewnętrzną znajduje się przestrzeń zwana peryplazmatyczną, wypełniona roztworem zawierającym białka transportowe i enzymy.
Po zewnętrznej stronie ściany komórkowej może znajdować się kapsułka - amorficzna warstwa, która zachowuje swoje połączenie ze ścianą. Warstwy śluzowe nie mają połączenia z komórką i łatwo się rozdzielają, podczas gdy osłonki nie są amorficzne, ale mają delikatną strukturę. Istnieje jednak wiele form przejściowych między tymi trzema wyidealizowanymi przypadkami.
Liczba wici bakteryjnej może wynosić od 0 do 1000. Obie opcje lokalizacji jednej wici na jednym biegunie (monotrych monopolarny), wiązki wici na jednym (monopolarny peritrich lub wici lophotrichial) lub na dwóch biegunach (bipolarny peritrich lub wici amphitricchial), oraz liczne wici na całej powierzchni komórki (peritrichous). Wić ma grubość 10–20 nm i długość 3–15 µm. Jego obrót odbywa się w kierunku przeciwnym do ruchu wskazówek zegara z częstotliwością 40-60 obr./min.
Oprócz wici wśród struktur powierzchniowych bakterii należy wymienić kosmki. Są cieńsze niż wici (średnica 5–10 nm, długość do 2 μm) i są niezbędne do przyczepienia się bakterii do podłoża; 3–10 μm) niż wici są niezbędne, aby komórka dawcy przeniosła DNA do biorcy podczas koniugacja.
Wymiary
Bakterie mają średnio 0,5–5 µm. Escherichia coli, na przykład ma rozmiar 0,3-1 na 1-6 mikronów, Staphylococcus aureus - średnicę 0,5-1 mikrona, Bacillus subtilis 0,75 na 2-3 mikrony. Największy z znane bakterie jest Thiomargarita namibiensis, osiągając rozmiar 750 mikronów (0,75 mm). Drugi to Epulopiscium fishelsoni, który ma średnicę 80 mikronów i długość do 700 mikronów i żyje w przewód pokarmowy ryby chirurgiczne Acanthurus nigrofuscus. Achromatium oxaliferum osiąga rozmiary 33 na 100 µm, Beggiatoa alba – 10 na 50 µm. Krętki mogą dorastać do 250 mikronów długości przy grubości 0,7 mikrona. Jednocześnie bakterie są najmniejszymi organizmami o budowie komórkowej. Mycoplasma mycoides mierzy 0,1–0,25 µm, co odpowiada wielkości dużych wirusów, takich jak mozaika tytoniu, krowianka lub grypa. Według obliczeń teoretycznych sferyczna komórka o średnicy mniejszej niż 0,15-0,20 mikrona staje się niezdolna do samoreprodukcji, ponieważ nie mieści fizycznie wszystkich niezbędnych biopolimerów i struktur w wystarczająco.Jednak opisano nanobakterie, które są mniejsze niż „dopuszczalne” i bardzo różnią się od zwykłych bakterii. W przeciwieństwie do wirusów są zdolne do niezależnego wzrostu i rozmnażania (niezwykle powolnego). Wciąż są mało zbadane, kwestionowana jest ich żywa natura.
Przy liniowym wzroście promienia komórki jej powierzchnia wzrasta proporcjonalnie do kwadratu promienia, a objętość - proporcjonalnie do sześcianu, dlatego u małych organizmów stosunek powierzchni do objętości jest większy niż u większych tych pierwszych, co oznacza dla tych pierwszych bardziej aktywną przemianę materii z otoczeniem. Aktywność metaboliczna mierzona przez różne wskaźniki, na jednostkę biomasy w formach małych jest wyższy niż w formach dużych. Dlatego małe rozmiary nawet dla mikroorganizmów dają bakteriom i archeonom przewagę w tempie wzrostu i rozmnażania w porównaniu z bardziej złożonymi organizmami eukariotycznymi i determinują ich ważną rolę ekologiczną.
Wielokomórkowość u bakterii
Formy jednokomórkowe zdolny do wykonywania wszystkich funkcji właściwych dla organizmu, niezależnie od sąsiednich komórek. Wiele jednokomórkowych prokariontów ma tendencję do tworzenia komórek, często utrzymywanych razem przez wydzielany przez nie śluz. W większości przypadków jest to przypadkowa kombinacja. poszczególne organizmy, ale w niektórych przypadkach tymczasowe skojarzenie wiąże się z realizacją określonej funkcji, na przykład tworzenie owocników przez myksobakterie sprawia, że możliwy rozwój cysty, mimo że pojedyncze komórki nie są w stanie ich wytworzyć. Takie zjawiska, wraz z tworzeniem się morfologicznie i funkcjonalnie zróżnicowanych komórek przez eubakterie jednokomórkowe, są niezbędnymi warunkami wstępnymi do pojawienia się w nich prawdziwej wielokomórkowości.Organizm wielokomórkowy musi spełniać następujące warunki:
- jego komórki muszą być agregowane,
- pomiędzy komórkami powinien być rozdział funkcji,
- między zagregowanymi komórkami powinny zostać ustanowione stabilne specyficzne kontakty.
Rozmnażanie bakterii
Niektóre bakterie nie mają procesu płciowego i rozmnażają się tylko przez binarne rozszczepienie poprzeczne równej wielkości lub pączkowanie. Dla jednej grupy jednokomórkowych cyjanobakterii opisano wielokrotny podział (seria szybkich, następujących po sobie podziałów binarnych, prowadzących do powstania od 4 do 1024 nowych komórek). Aby zapewnić plastyczność genotypu niezbędną do ewolucji i adaptacji do zmieniającego się środowiska, mają inne mechanizmy.Podczas podziału większość bakterii Gram-dodatnich i sinic nitkowatych syntetyzuje przegrodę poprzeczną od obwodu do środka z udziałem mezosomów. Bakterie Gram-ujemne dzielą się przez zwężenie: w miejscu podziału stwierdza się stopniowo zwiększającą się krzywiznę CPM i ściany komórkowej do wewnątrz. Podczas pączkowania nerka tworzy się i rośnie na jednym z biegunów komórki macierzystej, komórka macierzysta wykazuje oznaki starzenia i zwykle nie może wytworzyć więcej niż 4 komórki potomne. Pączkowanie występuje w różnych grupach bakterii i przypuszczalnie pojawiało się kilka razy w trakcie ewolucji.
U bakterii obserwuje się również rozmnażanie płciowe, ale w najbardziej prymitywnej formie. rozmnażanie płciowe bakterie różnią się od rozmnażania płciowego u eukariontów tym, że bakterie nie tworzą gamet i nie zachodzi fuzja komórek. Jednakże główne wydarzenie rozmnażanie płciowe, czyli wymiana materiału genetycznego, zachodzi również w tym przypadku. Ten proces nazywa się rekombinacją genetyczną. Część DNA (bardzo rzadko całe DNA) komórki dawcy jest przenoszona do komórki biorcy, której DNA różni się genetycznie od DNA dawcy. W takim przypadku przeniesione DNA zastępuje część DNA biorcy. Wymiana DNA obejmuje enzymy, które rozkładają i ponownie łączą nici DNA. W ten sposób powstaje DNA, które zawiera geny obu komórek rodzicielskich. Takie DNA nazywa się rekombinowanym. U potomstwa lub rekombinantów istnieje wyraźna różnorodność cech spowodowana błędem genetycznym. Ta różnorodność charakterów jest bardzo ważna dla ewolucji i jest główną zaletą rozmnażania płciowego. Istnieją 3 sposoby pozyskiwania rekombinantów. Są to, w kolejności ich odkrycia, transformacja, koniugacja i transdukcja.
2.1. Systematyka i nomenklatura drobnoustrojów
Świat drobnoustrojów można podzielić na formy komórkowe i niekomórkowe. Komórkowe formy drobnoustrojów reprezentowane są przez bakterie, grzyby i pierwotniaki. Można je nazwać mikroorganizmami. Formy niekomórkowe reprezentowane są przez wirusy, wiroidy i priony.
Nowa klasyfikacja drobnoustrojów komórkowych obejmuje następujące jednostki taksonomiczne: domeny, królestwa, typy, klasy, rzędy, rodziny, rodzaje, gatunki. Klasyfikacja mikroorganizmów opiera się na ich pokrewieństwie genetycznym, a także właściwościach morfologicznych, fizjologicznych, antygenowych i biologii molekularnej.
Wirusy są często uważane nie za organizmy, ale za autonomiczne struktury genetyczne, dlatego zostaną omówione osobno.
Formy komórkowe drobnoustrojów dzielą się na trzy domeny. Domeny bakteria I Archebakterie obejmują drobnoustroje o strukturze komórkowej typu prokariotycznego. Przedstawiciele domeny Eukarya są eukariontami. Składa się z 4 królestw:
Królestwa grzybów (Grzyby, Eumycota);
królestwa pierwotniaków (pierwotniaki);
królestwa Chromista(chrom);
Drobnoustroje o nieokreślonej pozycji taksonomicznej (Mikrospora, mikrosporydia).
Różnice w organizacji komórek prokariotycznych i eukariotycznych przedstawiono w tabeli. 2.1.
Tabela 2.1. Oznaki komórki prokariotycznej i eukariotycznej
2.2. Klasyfikacja i morfologia bakterii
Termin „bakterie” pochodzi od słowa bakteria, co oznacza różdżka. Bakterie są prokariotami. Są one podzielone na dwie domeny: bakteria I Archebakterie. Bakterie w domenie archebakterie, reprezentować jeden z starożytne formyżycie. Posiadają cechy strukturalne ściany komórkowej (brakuje im peptydoglikanu) oraz rybosomalnego RNA. Wśród nich nie ma patogenów chorób zakaźnych.
W obrębie domeny bakterie dzielą się na następujące kategorie taksonomiczne: klasa, gromada, rząd, rodzina, rodzaj, gatunek. Jedną z głównych kategorii taksonomicznych jest gatunek. Gatunek to zbiór osobników, które mają wspólne pochodzenie i genotyp, połączonych podobnymi właściwościami, które odróżniają je od innych członków rodzaju. Nazwa gatunku odpowiada nomenklaturze binarnej, tj. składa się z dwóch słów. Na przykład czynnik sprawczy błonicy jest zapisany jako Corynebacterium błonica. Pierwsze słowo jest nazwą rodzaju i jest pisane wielką literą, drugie słowo oznacza gatunek i jest pisane małą literą.
Kiedy gatunek jest wymieniany ponownie, nazwa rodzajowa jest na przykład skracana do pierwszej litery C. błonica.
Nazywa się zespół jednorodnych mikroorganizmów wyizolowanych na pożywce, charakteryzujących się podobnymi właściwościami morfologicznymi, barwiącymi (w stosunku do barwników), kulturowymi, biochemicznymi i antygenowymi czysta kultura. Nazywa się czystą kulturę mikroorganizmów wyizolowanych z określonego źródła i różniących się od innych członków gatunku napięcie. Bliskie pojęciu „szczepu” jest pojęcie „klonu”. Klon to zbiór potomstwa wyhodowanego z pojedynczej komórki drobnoustroju.
Do oznaczenia niektórych zestawów mikroorganizmów, które różnią się pewnymi właściwościami, stosuje się przyrostek „var” (odmiana), dlatego mikroorganizmy, w zależności od charakteru różnic, są określane jako morfowary (różnica w morfologii), produkty oporne (różnica w oporność, na przykład na antybiotyki), serowary (różnice w antygenach), fagowary (różnice we wrażliwości na bakteriofagi), biowary (różnice we właściwościach biologicznych), chemowary (różnice we właściwościach biochemicznych) itp.
Wcześniej podstawą klasyfikacji bakterii była strukturalna cecha ściany komórkowej. Podział bakterii według cech strukturalnych ściany komórkowej wiąże się z możliwą zmiennością ich zabarwienia w jednym lub drugim kolorze zgodnie z metodą Grama. Według tej metody, zaproponowanej w 1884 r. przez duńskiego naukowca H. Grama, w zależności od wyników barwienia, bakterie dzieli się na Gram-dodatnie, wybarwione na niebiesko-fioletowo i Gram-ujemne, wybarwione na czerwono.
Obecnie klasyfikacja opiera się na stopniu pokrewieństwa genetycznego, w oparciu o badanie struktury genomu rybosomalnego RNA (rRNA) (patrz rozdział 5), określając odsetek w genomie par guanina-cytozyna (pary GC), konstrukcja mapy restrykcyjnej genomu, badanie stopnia hybrydyzacji. Uwzględniane są również wskaźniki fenotypowe: stosunek do barwienia Grama, morfologiczne, kulturowe i właściwości biochemiczne, struktura antygenowa.
Domena bakteria obejmuje 23 typy, z których następujące mają znaczenie medyczne.
Większość bakterii Gram-ujemnych jest pogrupowana w typ Proteobakterie(wg nazwy grecki bóg Odmieniec, mogą przybierać różne formy). Typ Proteobakterie podzielone na 5 klas:
Klasa alfaproteobakterie(narodziny Rickettsia, Orientia, Erlichia, Bartonella, Brucella);
Klasa betaproteobakterie(narodziny Bordetella, Burholderia, Neisseria, Spirillum);
Klasa Gammaproteobakterie(członkowie rodziny Enterobacteriaceae, poród Francisella, Legionella, Coxiella, Pseudomonas, Vibrio);
Klasa Deltaproteobakterie(rodzaj bilofila);
Klasa Epsilonproteobakterie(narodziny Campylobacter, Helicobacter). Bakterie Gram-ujemne są również zaliczane do następujących typów:
typ Chlamydia(narodziny Chlamydie, Chlamydofile) typ krętki(narodziny Spirocheta, Borrelia, Treponema, Leptospira); typ Bacteroides(narodziny Bacteroides, Prevotella, Porphyromonas).
Bakterie Gram-dodatnie występują w następujących typach:
Typ Firmicutes obejmuje klasę Clostridium(narodziny Clostridium, Peptococcus), Klasa Bacilli (Listeria, Staphylococcus, Lactobacillus, Streptococcus) i klasa Mollicutes(narodziny mykoplazma, ureaplazma), które są bakteriami, które nie mają ściany komórkowej;
typ promieniowce(narodziny Actinomyces, Micrococcus, Corynebacterium, Mycobacterium, Gardnerella, Bifidobacterium, Propionibacterium, Mobiluncus).
2.2.1. Formy morfologiczne bakteria
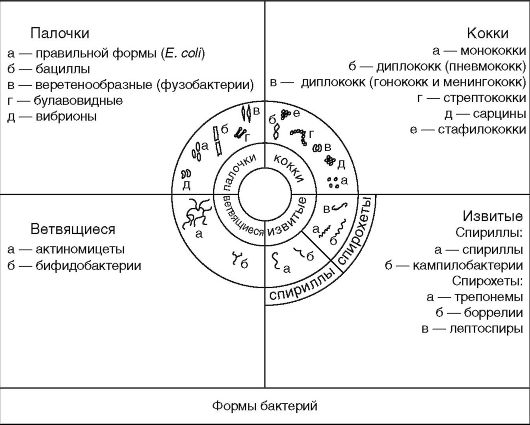
Istnieje kilka podstawowych form bakterii: kokosowate, pałeczkowate, skręcone i rozgałęzione (ryc. 2.1).
Kuliste kształty lub cocci- kuliste bakterie wielkości 0,5-1 mikrona, które dzielą się przez wzajemne ułożenie na mikrokoki, diplokoki, paciorkowce, tetrakoki, sarcyny i gronkowce.
Mikrokoki (z greckiego. mikro- małe) - oddzielnie zlokalizowane komórki.
Diplokoki (z gr. dyplomy- podwójne) lub sparowane ziarniaki, ułożone w pary (pneumokoki, gonokoki, meningokoki), ponieważ komórki nie rozchodzą się po podziale. Pneumokoki (czynnik wywołujący zapalenie płuc) mają lancetowaty kształt po przeciwnych stronach, a gonokoki (czynnik sprawczy rzeżączki) i meningokoki (czynnik sprawczy)
 Ryż. 2.1. Kształty bakterii
Ryż. 2.1. Kształty bakterii
przyczyną epidemicznego zapalenia opon mózgowych) mają kształt ziaren kawy, których wklęsłe powierzchnie są skierowane ku sobie.
Paciorkowce (z gr. paciorkowce- łańcuch) - komórki o zaokrąglonym lub wydłużonym kształcie, które tworzą łańcuch w wyniku podziału komórek w tej samej płaszczyźnie i zachowania połączenia między nimi w miejscu podziału.
Sarcins (z łac. Sarcina- wiązka, bela) są ułożone w postaci paczek po 8 ziarniaków lub więcej, ponieważ powstają podczas podziału komórki w trzech wzajemnie prostopadłych płaszczyznach.
Gronkowce (z gr. gronkowiec- kiść winogron) - ziarniaki ułożone w formie kiści winogron w wyniku podziału na różne płaszczyzny.
bakterie w kształcie pałeczek różnią się rozmiarem, kształtem końców komórki i względnym położeniem komórek. Długość komórki 1-10 µm, grubość 0,5-2 µm. Kije mogą mieć rację
(E. coli itp.) i nieregularne maczugi (corynebacteria itp.). Rickettsiae należą do najmniejszych bakterii w kształcie pałeczek.
Końce pałeczek mogą być niejako odcięte (bakterie wąglika), zaokrąglone (E. coli), spiczaste (fusobacteria) lub w postaci zgrubienia. W tym drugim przypadku patyk wygląda jak maczuga (Corynebacterium diphtheria).
Lekko zakrzywione pręty nazywane są vibrios (Vibrio cholerae). Większość bakterii w kształcie pałeczek układa się losowo, ponieważ po podziale komórki rozchodzą się. Jeśli po podziale komórki pozostają połączone wspólnymi fragmentami ściany komórkowej i nie rozchodzą się, to są one ustawione względem siebie pod kątem (corynebacterium diphtheria) lub tworzą łańcuch (bałteria wąglika).
Zawiłe formy- bakterie spiralne, które są dwojakiego rodzaju: spirilla i krętki. Spirilla ma wygląd zwiniętych komórek w kształcie korkociągu z dużymi lokami. Patogenne spirille obejmują czynnik wywołujący sodoku (chorobę pogryzioną przez szczura), a także Campylobacter i Helicobacteria, których krzywizny przypominają skrzydła mewy latającej. Krętki to cienkie, długie, zawiłe bakterie, które różnią się od spirilli mniejszymi lokami i charakterem ruchu. Poniżej opisano ich strukturę.
rozgałęzienie - bakterie w kształcie pałeczek, które mogą rozgałęziać się w formie litera łacińska Y, występujący w bifidobakterii, może być również prezentowany jako nitkowate, rozgałęzione komórki, które mogą splatać się, tworząc grzybnię, co obserwuje się u promieniowców.
2.2.2. Struktura komórki bakteryjnej
Struktura bakterii jest dobrze badana za pomocą mikroskopii elektronowej całych komórek i ich ultracienkich skrawków, a także innych metod. Komórka bakteryjna jest otoczona błoną składającą się ze ściany komórkowej i błony cytoplazmatycznej. Pod skorupą znajduje się protoplazma, składająca się z cytoplazmy z inkluzjami i aparatu dziedzicznego - analogu jądra, zwanego nukleoidem (ryc. 2.2). Istnieją dodatkowe struktury: kapsułka, mikrokapsułka, śluz, wici, pilusy. Niektóre bakterie w niesprzyjających warunkach są zdolne do tworzenia przetrwalników.
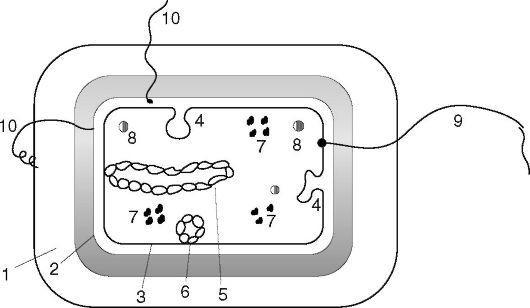 Ryż. 2.2. Budowa komórki bakteryjnej: 1 - otoczka; 2 - ściana komórkowa; 3 - błona cytoplazmatyczna; 4 - mezosomy; 5 - nukleoid; 6 - plazmid; 7 - rybosomy; 8 - inkluzje; 9 - wici; 10 - wypił (kosmki)
Ryż. 2.2. Budowa komórki bakteryjnej: 1 - otoczka; 2 - ściana komórkowa; 3 - błona cytoplazmatyczna; 4 - mezosomy; 5 - nukleoid; 6 - plazmid; 7 - rybosomy; 8 - inkluzje; 9 - wici; 10 - wypił (kosmki)
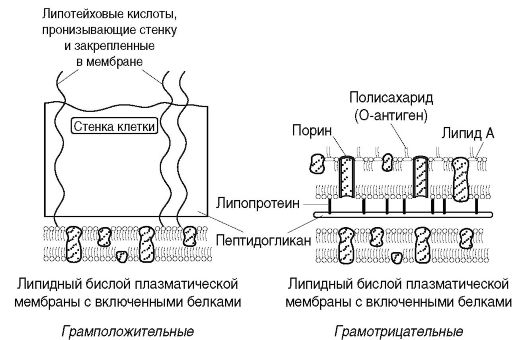
Ściana komórkowa- mocna, elastyczna struktura, która nadaje bakteriom określony kształt i wraz z leżącą pod nią błoną cytoplazmatyczną ogranicza wysokie ciśnienie osmotyczne w komórce bakteryjnej. Bierze udział w procesie podziału komórki i transporcie metabolitów, posiada receptory dla bakteriofagów, bakteriocyn i różne substancje. Najgrubsza ściana komórkowa bakterii Gram-dodatnich (ryc. 2.3). Tak więc, jeśli grubość ściany komórkowej bakterii Gram-ujemnych wynosi około 15-20 nm, to u bakterii Gram-dodatnich może osiągnąć 50 nm lub więcej.
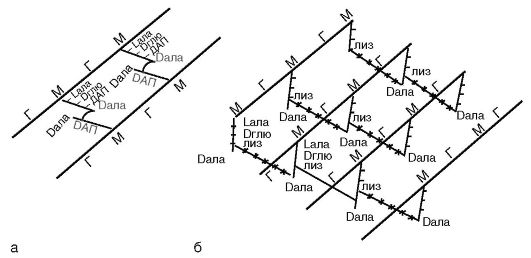
Ściana komórkowa bakterii jest zbudowana peptydoglikan. Peptydoglikan jest polimerem. Jest reprezentowany przez równoległe łańcuchy glikanów polisacharydowych, składające się z powtarzających się reszt N-acetyloglukozaminy i kwasu N-acetylomuraminowego połączonych wiązaniem glikozydowym. Wiązanie to jest rozrywane przez lizozym, który jest acetylomuramidazą.
Tetrapeptyd jest przyłączony do kwasu N-acetylomuraminowego wiązaniami kowalencyjnymi. Tetrapeptyd składa się z L-alaniny, która jest połączona z kwasem N-acetylomuraminowym; D-glutamina, która u bakterii Gram-dodatnich jest połączona z L-lizyną, a u bakterii Gram-dodatnich
 Ryż. 2.3. Schemat architektury ściany komórkowej bakterii
Ryż. 2.3. Schemat architektury ściany komórkowej bakterii
bakterie – z kwasem diaminopimelinowym (DAP), który jest prekursorem lizyny w procesie bakteryjnej biosyntezy aminokwasów i jest unikalnym związkiem występującym tylko u bakterii; Czwartym aminokwasem jest D-alanina (ryc. 2.4).
Ściana komórkowa bakterii Gram-dodatnich zawiera niewielką ilość polisacharydów, lipidów i białek. Głównym składnikiem ściany komórkowej tych bakterii jest wielowarstwowy peptydoglikan (mureina, mukopeptyd), który stanowi 40-90% masy ściany komórkowej. Tetrapeptydy różnych warstw peptydoglikanu w bakteriach Gram-dodatnich są połączone ze sobą łańcuchami polipeptydowymi złożonymi z 5 reszt glicyny (pentaglicyny), co nadaje peptydoglikanowi sztywną strukturę geometryczną (ryc. 2.4, b). Kowalencyjnie związany z peptydoglikanem ściany komórkowej bakterii Gram-dodatnich kwasy teichojowe(z gr. techos- ściana), której cząsteczkami są łańcuchy 8-50 reszt glicerolu i rybitolu połączone mostkami fosforanowymi. Kształt i wytrzymałość bakterii wynika ze sztywnej włóknistej struktury wielowarstwowej z usieciowanymi peptydowymi wiązaniami krzyżowymi peptydoglikanu.
 Ryż. 2.4. Budowa peptydoglikanu: a - bakterie Gram-ujemne; b - bakterie Gram-dodatnie
Ryż. 2.4. Budowa peptydoglikanu: a - bakterie Gram-ujemne; b - bakterie Gram-dodatnie
Zdolność bakterii Gram-dodatnich do zatrzymywania fioletu goryczki w połączeniu z jodem (niebiesko-fioletowy kolor bakterii) podczas barwienia metodą Grama jest związana z właściwością wielowarstwowego peptydoglikanu do interakcji z barwnikiem. Ponadto późniejsze traktowanie rozmazu bakterii alkoholem powoduje zwężenie porów w peptydoglikanie i tym samym zatrzymuje barwnik w ścianie komórkowej.
bakterie Gram-ujemne po ekspozycji na alkohol tracą barwnik, co jest spowodowane mniejszą ilością peptydoglikanu (5-10% masy ściany komórkowej); odbarwiają się alkoholem, a po potraktowaniu fuksyną lub safraniną stają się czerwone. Wynika to z cech strukturalnych ściany komórkowej. Peptydoglikan w ścianie komórkowej bakterii Gram-ujemnych jest reprezentowany przez 1-2 warstwy. Tetrapeptydy warstw są połączone bezpośrednim wiązaniem peptydowym między grupą aminową DAP jednego tetrapeptydu a grupą karboksylową D-alaniny tetrapeptydu innej warstwy (ryc. 2.4, a). Na zewnątrz peptydoglikanu znajduje się warstwa lipoproteina, wiąże się z peptydoglikanem przez DAP. Po nim następuje zewnętrzna męmbrana Ściana komórkowa.
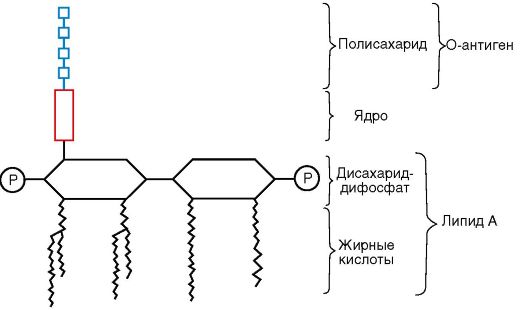
zewnętrzna męmbrana jest strukturą mozaikową reprezentowaną przez lipopolisacharydy (LPS), fosfolipidy i białka. Jej wewnętrzną warstwę reprezentują fosfolipidy, a LPS znajduje się w warstwie zewnętrznej (ryc. 2.5). W ten sposób zewnętrzna membrana
 Ryż. 2.5. Struktura lipopolisacharydu
Ryż. 2.5. Struktura lipopolisacharydu
brana jest asymetryczna. LPS błony zewnętrznej składa się z trzech fragmentów:
Lipid A - konserwatywna struktura, prawie taka sama u bakterii Gram-ujemnych. Lipid A składa się z fosforylowanych jednostek disacharydowych glukozaminy, do których przyłączone są długie łańcuchy. Kwasy tłuszczowe(patrz rys. 2.5);
Rdzeń lub pręt części krowiej (z łac. rdzeń- rdzeń), względnie konserwatywna struktura oligosacharydów;
Wysoce zmienny O-specyficzny łańcuch polisacharydowy utworzony przez powtarzanie identycznych sekwencji oligosacharydowych.
LPS jest zakotwiczony w błonie zewnętrznej przez lipid A, który określa toksyczność LPS i dlatego jest identyfikowany z endotoksyną. Zniszczenie bakterii przez antybiotyki prowadzi do uwolnienia dużych ilości endotoksyn, które mogą wywołać u pacjenta wstrząs endotoksyczny. Z lipidu A odchodzi rdzeń lub część rdzenia LPS. Najbardziej stałą częścią rdzenia LPS jest kwas ketodeoksyoktonowy. O-specyficzny łańcuch polisacharydowy rozciągający się od części rdzeniowej cząsteczki LPS,
składający się z powtarzających się jednostek oligosacharydowych, określa serogrupę, serowar (rodzaj bakterii wykrywany za pomocą surowicy odpornościowej) określonego szczepu bakterii. Tak więc koncepcja LPS jest związana z ideami dotyczącymi antygenu O, według którego można różnicować bakterie. Zmiany genetyczne mogą prowadzić do defektów, skrócenia bakteryjnego LPS, aw efekcie do pojawienia się szorstkich kolonii form R, które tracą swoją specyficzność względem antygenu O.
Nie wszystkie bakterie Gram-ujemne mają kompletny łańcuch polisacharydowy O-specyficzny składający się z powtarzających się jednostek oligosacharydowych. W szczególności bakterie z rodzaju Neisseria mają krótki glikolipid zwany lipooligosacharydem (LOS). Jest porównywalny z formą R, która utraciła specyficzność antygenową O, obserwowaną w zmutowanych szczepach szorstkich. E coli. Struktura LZO przypomina strukturę glikosfingolipidu błony cytoplazmatycznej człowieka, więc LZO naśladuje mikroba, pozwalając mu uniknąć odpowiedzi immunologicznej gospodarza.
Białka macierzy błony zewnętrznej przenikają przez nią w taki sposób, że cząsteczki białka, tzw poryny, graniczą z porami hydrofilowymi, przez które przechodzi woda i małe cząsteczki hydrofilowe o masie względnej do 700 D.
Pomiędzy błonami zewnętrznymi i cytoplazmatycznymi znajduje się Przestrzeń periplazmatyczna, lub peryplazmy zawierające enzymy (proteazy, lipazy, fosfatazy, nukleazy, β-laktamazy), a także składniki systemów transportowych.
W przypadku naruszenia syntezy ściany komórkowej bakterii pod wpływem lizozymu, penicyliny, czynników ochronnych organizmu i innych związków, powstają komórki o zmienionym (często kulistym) kształcie: protoplasty- bakterie całkowicie pozbawione ściany komórkowej; sferoplasty Bakterie z częściowo zachowaną ścianą komórkową. Po usunięciu inhibitora ściany komórkowej, takie zmienione bakterie mogą się odwrócić, tj. uzyskać pełnoprawną ścianę komórkową i przywrócić jej pierwotny kształt.
Bakterie typu sferoidalnego lub protoplastowego, które utraciły zdolność syntezy peptydoglikanu pod wpływem antybiotyków lub innych czynników i są zdolne do namnażania, nazywane są w kształcie litery L(od nazwy Instytutu D. Listera, gdzie po raz pierwszy
studiowałeś). Formy L mogą również powstać w wyniku mutacji. Są to wrażliwe osmotycznie, kuliste, kolbowate komórki o różnej wielkości, w tym przechodzące przez filtry bakteryjne. Niektóre formy L (niestabilne), gdy czynnik, który doprowadził do zmian w bakteriach, zostanie usunięty, mogą się odwrócić, powracając do pierwotnej komórki bakteryjnej. Formy L mogą tworzyć wiele patogenów chorób zakaźnych.
błona cytoplazmatyczna pod mikroskopem elektronowym ultracienkich skrawków jest to membrana trójwarstwowa (2 ciemne warstwy o grubości 2,5 nm każda oddzielone są jedną jasną - pośrednią). W strukturze jest podobny do plazmolemy komórek zwierzęcych i składa się z podwójnej warstwy lipidów, głównie fosfolipidów, z osadzonymi białkami powierzchniowymi i integralnymi, jakby przenikającymi przez strukturę błony. Niektóre z nich to permeazy biorące udział w transporcie substancji. w odróżnieniu komórki eukariotyczne, w błonie cytoplazmatycznej komórki bakteryjnej (z wyjątkiem mykoplazmy) nie ma steroli.
Błona cytoplazmatyczna jest strukturą dynamiczną z ruchomymi elementami, dlatego przedstawiana jest jako ruchoma płynna struktura. Otacza zewnętrzną część cytoplazmy bakterii i bierze udział w regulacji ciśnienia osmotycznego, transporcie substancji i metabolizmie energetycznym komórki (za sprawą enzymów łańcucha transportu elektronów, adenozynotrifosfatazy – ATPazy itp.). Przy nadmiernym wzroście (w porównaniu ze wzrostem ściany komórkowej) błona cytoplazmatyczna tworzy wklęsłości - wklęsłości w postaci skomplikowanych skręconych struktur błonowych, tzw. mezosomy. Mniej złożone skręcone struktury nazywane są błonami wewnątrzcytoplazmatycznymi. Rola mezosomów i błon wewnątrzcytoplazmatycznych nie została w pełni wyjaśniona. Sugeruje się nawet, że są one artefaktem powstałym po przygotowaniu (utrwaleniu) preparatu do mikroskopii elektronowej. Niemniej jednak uważa się, że pochodne błony cytoplazmatycznej biorą udział w podziale komórki, dostarczając energii do syntezy ściany komórkowej, biorą udział w wydzielaniu substancji, tworzeniu przetrwalników, tj. w procesach o dużym zużyciu energii. Cytoplazma zajmuje większość bakterii
komórka końcowa i składa się z rozpuszczalnych białek, kwasów rybonukleinowych, inkluzji i licznych małych granulek - rybosomów odpowiedzialnych za syntezę (translację) białek.
Rybosomy bakterie mają wielkość około 20 nm i współczynnik sedymentacji 70S, w przeciwieństwie do charakterystycznych dla komórek eukariotycznych rybosomów 80S. Dlatego niektóre antybiotyki wiążą się z rybosomami bakteryjnymi i hamują syntezę białek bakteryjnych bez wpływu na syntezę białek w komórkach eukariotycznych. Rybosomy bakteryjne mogą dysocjować na dwie podjednostki: 50S i 30S. rRNA - konserwatywne elementy bakterii ("zegar molekularny" ewolucji). 16S rRNA jest częścią małej podjednostki rybosomów, a 23S rRNA jest częścią dużej podjednostki rybosomów. Badanie 16S rRNA stanowi podstawę systematyki genów, umożliwiając ocenę stopnia pokrewieństwa organizmów.
W cytoplazmie znajdują się różne inkluzje w postaci granulek glikogenu, polisacharydów, kwasu β-hydroksymasłowego i polifosforanów (wolutina). Kumulują się wraz z nadmiarem składników pokarmowych w środowisku i pełnią rolę substancji rezerwowych dla potrzeb żywieniowych i energetycznych.
Woliutin ma powinowactwo do barwników zasadowych i jest łatwo wykrywalny za pomocą specjalnych metod barwienia (na przykład według Neissera) w postaci metachromatycznych granulek. Błękit toluidynowy lub błękit metylenowy barwi wolutinę na czerwono-fioletowo, a cytoplazmę bakteryjną na niebiesko. Charakterystyczny układ ziarnistości wolutyny ujawnia się w prątku błonicy w postaci intensywnie wybarwionych biegunów komórki. Związane jest z metachromatycznym barwieniem wolutyny wysoka zawartość spolimeryzowany nieorganiczny polifosforan. Pod mikroskopem elektronowym wyglądają jak gęste elektronowo granulki o wielkości 0,1–1 µm.
nukleoid jest odpowiednikiem jądra w bakteriach. Znajduje się w centralnej strefie bakterii w postaci dwuniciowego DNA, ciasno upakowanego jak kula. Bakteryjny nukleoid, w przeciwieństwie do eukariontów, nie ma otoczka jądrowa, jąderko i główne białka (histony). Większość bakterii zawiera jeden chromosom, reprezentowany przez cząsteczkę DNA zamkniętą w pierścieniu. Ale niektóre bakterie mają dwa chromosomy w kształcie pierścienia. (V. cholera) i chromosomy liniowe (patrz sekcja 5.1.1). Nukleoid jest wykrywany pod mikroskopem świetlnym po barwieniu specyficznym DNA
metody: według Felgena lub według Romanowskiego-Giemsy. Na wzorach dyfrakcji elektronów ultracienkich skrawków bakterii nukleoid ma postać jasnych stref z włóknistymi, nitkowatymi strukturami DNA związanymi z pewnymi obszarami z błoną cytoplazmatyczną lub mezosomem zaangażowanym w replikację chromosomu.
Oprócz nukleoidu komórka bakteryjna zawiera pozachromosomalne czynniki dziedziczności - plazmidy (patrz rozdział 5.1.2), które są kowalencyjnie zamkniętymi pierścieniami DNA.
Kapsułka, mikrokapsułka, śluz.Kapsuła - struktura śluzowa o grubości ponad 0,2 mikrona, mocno związana ze ścianą komórkową bakterii i mająca wyraźnie określone granice zewnętrzne. Kapsułka jest rozpoznawalna w rozmazach-odciskach z materiału patologicznego. W czyste kultury torebka bakteryjna powstaje rzadziej. Ujawnia się o godz specjalne metody barwienie rozmazu wg Burri-Ginsa, tworzące negatywny kontrast substancji kapsułki: tusz tworzy ciemne tło wokół kapsułki. Kapsułka składa się z polisacharydów (egzopolisacharydów), czasami polipeptydów, na przykład w prątku wąglika składa się z polimerów kwasu D-glutaminowego. Kapsułka jest hydrofilowa, zawiera dużą ilość wody. Zapobiega fagocytozie bakterii. Kapsułka jest antygenowa: przeciwciała skierowane przeciwko otoczce powodują jej wzrost (reakcja pęcznienia kapsułki).
Powstaje wiele bakterii mikrokapsułka- tworzenie śluzu o grubości mniejszej niż 0,2 mikrona, wykrywane tylko za pomocą mikroskopii elektronowej.
Aby odróżnić od kapsułki szlam - egzopolisacharydy śluzowe, które nie mają wyraźnych granic zewnętrznych. Slime jest rozpuszczalny w wodzie.
Egzopolisacharydy śluzowe są charakterystyczne dla śluzowatych szczepów Pseudomonas aeruginosa, często znajdowanych w plwocinie pacjentów z mukowiscydozą. Egzopolisacharydy bakteryjne biorą udział w adhezji (przyklejaniu się do podłoża); nazywane są również glikokaliksem.
Torebka i śluz chronią bakterie przed uszkodzeniem i wysychaniem, ponieważ będąc hydrofilowymi, dobrze wiążą wodę i zapobiegają działaniu czynników ochronnych makroorganizmów i bakteriofagów.
wici bakterie determinują ruchliwość komórki bakteryjnej. Wici to cienkie włókna, które nabierają
pochodzące z błony cytoplazmatycznej są dłuższe niż sama komórka. Wici mają grubość 12–20 nm i długość 3–15 µm. Składają się z trzech części: spiralnej nitki, haczyka i korpusu podstawowego zawierającego pręcik ze specjalnymi krążkami (jedna para krążków u bakterii Gram-dodatnich i dwie pary u bakterii Gram-ujemnych). Krążki wici są przyczepione do błony cytoplazmatycznej i ściany komórkowej. Tworzy to efekt silnika elektrycznego z prętem - wirnikiem, który obraca wić. Jako źródło energii wykorzystuje się różnicę potencjałów protonów na błonie cytoplazmatycznej. Mechanizm rotacji zapewnia syntetaza protonu ATP. Prędkość obrotu wici może osiągnąć 100 obrotów na minutę. Jeśli bakteria ma kilka wici, zaczynają one obracać się synchronicznie, splatając się w jedną wiązkę, tworząc coś w rodzaju śmigła.
Wici składają się z białka zwanego flageliną. (rozłóg- wici), która jest antygenem - tzw. antygenem H. Podjednostki flageliny są zwinięte.
Liczba wici u bakterii różne rodzaje waha się od jednego (monotrich) u Vibrio cholerae do dziesięciu lub setek rozciągających się wzdłuż obwodu bakterii (peritrich), w coli, proteus itp. Lofotrichous mają wiązkę wici na jednym końcu komórki. Amphitrichous mają jedną wici lub wiązkę wici na przeciwległych końcach komórki.
Wici wykrywa się za pomocą mikroskopii elektronowej preparatów spryskanych metalami ciężkimi lub w mikroskopie świetlnym po przetworzeniu specjalnymi metodami opartymi na trawieniu i adsorpcji różnych substancji, prowadzących do zwiększenia grubości wici (np. po srebrzenie).
Kosmki lub pilusy (fimbriae)- nitkowate formacje, cieńsze i krótsze (3-10 nm * 0,3-10 mikronów) niż wici. Pili wystają z powierzchni komórki i składają się z białka pilinu. Znanych jest kilka rodzajów pił. picie typ ogólny odpowiada za przyczepność do podłoża, odżywianie i wymiana wodno-solna. Jest ich bardzo dużo - po kilkaset na komórkę. Pilusy płciowe (1-3 na komórkę) tworzą kontakt między komórkami, przekazując między nimi informację genetyczną poprzez koniugację (patrz rozdział 5). Szczególnie interesujące są pilusy typu IV, w których końce są hydrofobowe, w wyniku czego się skręcają, pilusy te nazywane są również lokami. Usytuowany-
znajdują się na biegunach komórki. Piły te znajdują się w bakterie chorobotwórcze. Mają właściwości antygenowe, wchodzą w kontakt między bakterią a komórką gospodarza oraz uczestniczą w tworzeniu biofilmu (patrz rozdział 3). Wiele pilusów jest receptorami dla bakteriofagów.
Spory - specyficzna forma bakterii spoczynkowych o Gram-dodatnim typie budowy ściany komórkowej. bakterie tworzące przetrwalniki z rodzaju bakcyl, w których wielkość zarodnika nie przekracza średnicy komórki, nazywane są pałeczkami. Bakterie tworzące przetrwalniki, w których wielkość przetrwalnika przekracza średnicę komórki, dlatego przybierają postać wrzeciona, nazywane są Clostridia, takich jak bakterie z rodzaju Clostridium(od łac. Clostridium- wrzeciono). Zarodniki są kwasoodporne, dlatego wybarwiają się na czerwono według metody Aujeszky'ego lub według metody Ziehla-Nelsena, a komórka wegetatywna jest niebieska.
Zarodnikowanie, kształt i umiejscowienie zarodników w komórce (wegetatywne) to cecha gatunkowa bakterii, która umożliwia ich odróżnienie. Kształt zarodników jest owalny i kulisty, lokalizacja w komórce jest końcowa, tj. na końcu sztyftu (w czynniku sprawczym tężca), podterminalny - bliżej końca sztyftu (w czynnikach sprawczych zatrucia jadem kiełbasianym, zgorzel gazowa) i centralny (w prątku wąglika).
Proces sporulacji (sporulacji) przebiega przez szereg etapów, podczas których część cytoplazmy i chromosom wegetatywnej komórki bakteryjnej są oddzielane, otoczone rosnącą błoną cytoplazmatyczną i powstaje prospora.
Protoplast prospory zawiera nukleoid, system syntezy białek i system wytwarzania energii oparty na glikolizie. Cytochromy są nieobecne nawet w tlenowcach. Nie zawiera ATP, energia do kiełkowania magazynowana jest w postaci fosforanu 3-glicerolu.
Prospora jest otoczona dwiema błonami cytoplazmatycznymi. Nazywa się warstwę otaczającą wewnętrzną błonę zarodnika ściana zarodników, składa się z peptydoglikanu i jest głównym źródłem ściany komórkowej podczas kiełkowania zarodników.
Pomiędzy zewnętrzną błoną a ścianą zarodnika tworzy się gruba warstwa, składająca się z peptydoglikanu, który ma wiele wiązań poprzecznych, - kora.
Na zewnątrz znajduje się zewnętrzna błona cytoplazmatyczna skorupa zarodników, składający się z białek podobnych do keratyny,
zawierające wiele wewnątrzcząsteczkowych wiązań dwusiarczkowych. Powłoka ta zapewnia odporność na czynniki chemiczne. Zarodniki niektórych bakterii mają dodatkową osłonę - egzosporium charakter lipoproteinowy. W ten sposób powstaje wielowarstwowa słabo przepuszczalna otoczka.
Zarodnikowaniu towarzyszy intensywna konsumpcja przez prosporę, a następnie wyłaniająca się otoczka zarodników kwasu dipikolinowego i jonów wapnia. Zarodnik nabywa odporność na ciepło, co jest związane z obecnością w nim dipikolinianu wapnia.
Zarodnik może przetrwać przez długi czas ze względu na obecność wielowarstwowej otoczki, dipikolinianu wapnia, niskiej zawartości wody i powolnych procesów metabolicznych. Na przykład w glebie patogeny wąglika i tężca mogą przetrwać przez dziesięciolecia.
W sprzyjających warunkach zarodniki kiełkują przez trzy kolejne etapy: aktywację, inicjację, wzrost. W tym przypadku jedna bakteria powstaje z jednego zarodnika. Aktywacja to gotowość do kiełkowania. W temperaturze 60-80°C zarodnik jest aktywowany do kiełkowania. Inicjacja kiełkowania trwa kilka minut. Faza wzrostu charakteryzuje się szybkim wzrostem, któremu towarzyszy zniszczenie skorupy i uwolnienie sadzonki.
2.2.3. Cechy struktury krętków, riketsj, chlamydii, promieniowców i mykoplazm
Krętki- cienkie, długie, zawiłe bakterie. Składają się z zewnętrznej błoniastej ściany komórkowej otaczającej cylinder cytoplazmatyczny. Na wierzchu błony zewnętrznej znajduje się przezroczysta otoczka o charakterze glikozoaminoglikanu. Pod zewnętrzną ścianą komórkową błony znajdują się fibryle, które owijają się wokół cylindra cytoplazmatycznego, nadając bakteriom spiralny kształt. Fibryle są przyczepione do końców komórki i skierowane ku sobie. Liczba i układ włókienek różni się u różnych gatunków. Fibryle biorą udział w ruchu krętków, nadając komórkom ruch obrotowy, zginający i translacyjny. W tym przypadku krętki tworzą pętle, loki, zagięcia, które nazywane są lokami wtórnymi. Krętki źle postrzegają barwniki. Zwykle są barwione według Romanowskiego-Giemsy lub srebrzone. Na żywo
forma krętka jest badana za pomocą mikroskopii z kontrastem fazowym lub ciemnego pola.
Krętki są reprezentowane przez trzy rodzaje patogenne dla człowieka: Treponema, Borrelia, Leptospira.
treponema(rodzaj treponema) mają wygląd cienkich nitek skręconych korkociągiem z 8-12 jednolitymi małymi lokami. Wokół protoplastu treponemy znajdują się 3-4 włókienka (wici). Cytoplazma zawiera włókna cytoplazmatyczne. Przedstawiciele patogenów są T. pallidum- czynnik sprawczy kiły T. pertenue- patogen choroba tropikalna- odchylenia. Są też saprofity - mieszkańcy jamy ustnej człowieka, muł zbiorników wodnych.
Borrelia(rodzaj Borrelia, w przeciwieństwie do krętków są dłuższe, mają 3-8 dużych loków i 7-20 włókienek. Należą do nich czynnik wywołujący nawracającą gorączkę (B. recurrentis) i czynniki wywołujące boreliozę (B. burgdorferi) i inne choroby.
Leptospira(rodzaj Leptospira) mają płytkie i częste loki w postaci skręconej liny. Końce tych krętków są zakrzywione jak haczyki ze zgrubieniami na końcach. Tworząc wtórne loki, przybierają postać liter S lub C; mają dwa włókienka osiowe. Przedstawiciel chorobotwórczy L. przesłuchujący powoduje leptospirozę po spożyciu z wodą lub pokarmem, prowadząc do krwotoków i żółtaczki.
Rickettsia mają metabolizm niezależny od komórki gospodarza, jednak mogą otrzymywać związki makroergiczne z komórki gospodarza w celu rozmnażania. W rozmazach i tkankach są one barwione według Romanovsky'ego-Giemsy, według Machiavello-Zdrodovsky'ego (riketsje są czerwone, a zakażone komórki są niebieskie).
U ludzi riketsje powodują epidemię dur plamisty (R. prowazekii), riketsjoza przenoszona przez kleszcze (R. sibirica), gorączka plamista Gór Skalistych (R. rickettsii) i inne riketsjozy.
Struktura ich ściany komórkowej przypomina bakterie Gram-ujemne, chociaż istnieją różnice. Nie zawiera typowego peptydoglikanu: kwas N-acetylomuraminowy jest całkowicie nieobecny w jego składzie. Ściana komórkowa składa się z podwójnej błony zewnętrznej, która zawiera lipopolisacharyd i białka. Pomimo braku peptydoglikanu ściana komórkowa chlamydii jest sztywna. Cytoplazma komórki jest ograniczona wewnętrzną błoną cytoplazmatyczną.
Główną metodą wykrywania chlamydii jest barwienie Romanowskiego-Giemsy. Kolor plamy zależy od etapu cyklu życiowego: ciałka elementarne stają się fioletowe na tle niebieskiej cytoplazmy komórki, ciałka siatkowate stają się niebieskie.
U ludzi chlamydia powoduje uszkodzenia oczu (jaglica, zapalenie spojówek), układu moczowo-płciowego, płuc itp.
promieniowce- rozgałęzione, nitkowate lub pałeczkowate bakterie Gram-dodatnie. Jego nazwa (z gr. akt- Ray, Mykes- grzyb) otrzymali w związku z tworzeniem się druz w dotkniętych tkankach - granulki ściśle splecionych nitek w postaci
promienie rozciągające się od środka i kończące się zgrubieniami w kształcie kolb. Promieniowce, podobnie jak grzyby, tworzą grzybnię - nitkowate przeplatające się komórki (strzępki). Tworzą grzybnię substratową, która powstaje w wyniku wrastania komórek w pożywkę oraz powietrze, narastające na powierzchni pożywki. Promieniowce mogą dzielić się poprzez fragmentację grzybni na komórki podobne do bakterii pałkowatych i kokosowych. Na strzępkach powietrznych promieniowców powstają zarodniki, które służą do rozmnażania. Zarodniki promieniowców zwykle nie są odporne na ciepło.
Wspólną gałąź filogenetyczną z promieniowcami tworzą tak zwane promieniowce nokardiopodobne (nocardioform) - zbiorowa grupa bakterii w kształcie pałeczek nieregularny kształt. Ich poszczególni przedstawiciele tworzą formy rozgałęzione. Należą do nich bakterie z rodzajów Corynebacterium, Mycobacterium, Nocardia promieniowce nokardioidalne wyróżniają się obecnością w ścianie komórkowej cukrów arabinozy, galaktozy, a także kwasów mikolowych i dużych ilości kwasów tłuszczowych. Kwasy mikolowe i lipidy ściany komórkowej decydują o kwasoodporności bakterii, w szczególności Mycobacterium tuberculosis i trądu (po barwieniu wg Ziehla-Nelsena są one czerwone, a bakterie niekwasoodporne i elementy tkankowe, plwocina jest niebieska).
Promieniowce chorobotwórcze powodują promienicę, nokardiozę - nokardiozę, prątki - gruźlicę i trąd, maczugowce - błonicę. Saprofityczne formy promieniowców i promieniowców nokardiopodobnych są szeroko rozpowszechnione w glebie, wiele z nich jest producentami antybiotyków.
Mykoplazmy- małe bakterie (0,15-1 µm) otoczone jedynie błoną cytoplazmatyczną zawierającą sterole. Należą do klasy Mollicutes. Ze względu na brak ściany komórkowej mykoplazmy są wrażliwe osmotycznie. Mają różnorodne kształty: kokosowe, nitkowate, kolbowate. Formy te są widoczne w mikroskopie z kontrastem fazowym czystych kultur mykoplazmy. Na gęstej pożywce mykoplazmy tworzą kolonie przypominające jajka sadzone: środkowa nieprzezroczysta część zanurzona w pożywce i półprzezroczyste obrzeże w kształcie koła.
Mykoplazmy powodują SARS u ludzi (Mycoplasma pneumoniae) i uszkodzenia dróg moczowych
(M. człowiek itd.). Mykoplazmy powodują choroby nie tylko zwierząt, ale także roślin. Przedstawiciele niepatogenni są dość rozpowszechnioni.
2.3. Budowa i klasyfikacja grzybów
Grzyby należą do domeny eukaria, Królestwo Grzyby (Mycota, Mycetes). Grzyby i pierwotniaki zostały ostatnio podzielone na niezależne królestwa: królestwo Eumycota(prawdziwe grzyby), królestwo Chromista i królestwo pierwotniaki. Niektóre mikroorganizmy, które wcześniej uważano za grzyby lub pierwotniaki, zostały przeniesione do nowego królestwa Chromista(chromy). Grzyby to wielokomórkowe lub jednokomórkowe niefotosyntetyczne (wolne od chlorofilu) mikroorganizmy eukariotyczne o grubej ścianie komórkowej. Mają jądro z otoczką jądrową, cytoplazmę z organellami, cyto błona plazmatyczna oraz wielowarstwowa sztywna ściana komórkowa, składająca się z kilku rodzajów polisacharydów (mannanów, glukanów, celulozy, chityny), a także białka, lipidów itp. Niektóre grzyby tworzą otoczkę. Błona cytoplazmatyczna zawiera glikoproteiny, fosfolipidy i ergosterole (w przeciwieństwie do cholesterolu, głównego sterolu tkanek ssaków). Większość grzybów to obligatoryjne lub fakultatywne tlenowce.
Grzyby są szeroko rozpowszechnione w przyrodzie, zwłaszcza w glebie. Niektóre grzyby przyczyniają się do produkcji chleba, sera, produktów mlecznych i alkoholu. Inne grzyby wytwarzają antybiotyki przeciwdrobnoustrojowe (np. penicylinę) i leki immunosupresyjne (np. cyklosporynę). Grzyby są wykorzystywane przez genetyków i biologów molekularnych do modelowania różnych procesów. Grzyby fitopatogenne powodują znaczne szkody w rolnictwie, powodując m.in choroby grzybowe rośliny zbożowe i ziarna. Infekcje wywołane przez grzyby nazywane są grzybicami. Występują strzępki i drożdżaki.
Grzyby strzępkowe (pleśniowe) lub hyphomycetes składają się z cienkie nici Grubości 2-50 mikronów, zwane strzępkami, które są wplecione w grzybnię lub grzybnię (pleśń). Ciało grzyba nazywa się plechą. Rozróżnij hyphomycetes demacia (pigmentowane - brązowe lub czarne) i hialinowe (niepigmentowane). Strzępki wrastające w podłoże odżywcze są odpowiedzialne za odżywianie grzyba i nazywane są strzępkami wegetatywnymi. strzępki, ra-
wyrastające ponad powierzchnię podłoża nazywane są strzępkami powietrznymi lub rozrodczymi (odpowiedzialnymi za rozmnażanie). Kolonie spowodowane grzybnią powietrzną mają puszysty wygląd.
Istnieją grzyby niższe i wyższe: strzępki grzybów wyższych są oddzielone przegrodami lub przegrodami z otworami. Strzępki grzybów niższych nie mają przegród, reprezentujących komórki wielojądrzaste zwane koenocytami (z gr. koenos- pojedynczy, ogólny).
Grzyby drożdżowe (drożdże) są reprezentowane głównie przez pojedyncze owalne komórki o średnicy 3-15 mikronów, a ich kolonie, w przeciwieństwie do grzybów strzępkowych, mają zwarty wygląd. W zależności od rodzaju rozmnażania płciowego są one rozmieszczone wśród grzybów wyższych - ascomycete i basidiomycete. Podczas rozmnażania bezpłciowego drożdże tworzą pąki lub dzielą się. Mogą tworzyć pseudostrzępki i grzybnię fałszywą (pseudomycelium) w postaci łańcuchów wydłużonych komórek - „parówek”. Grzyby, które są podobne do drożdży, ale nie rozmnażają się płciowo, nazywane są drożdżopodobnymi. Rozmnażają się wyłącznie bezpłciowo - przez pączkowanie lub podział. Pojęcia „grzyby drożdżopodobne” są często utożsamiane z pojęciem „drożdże”.
Wiele grzybów ma dymorfizm - zdolność do wzrostu strzępkowego (grzybniowego) lub drożdżopodobnego, w zależności od warunków uprawy. W zakażonym organizmie rosną jako komórki drożdżopodobne (faza drożdży) i tworzą strzępki i grzybnię na pożywce. Dymorfizm jest związany z czynnikiem temperaturowym: w temperaturze pokojowej tworzy się grzybnia, aw temperaturze 37 ° C (w temperaturze ciała człowieka) powstają komórki drożdżopodobne.
Grzyby rozmnażają się płciowo lub bezpłciowo. Rozmnażanie płciowe grzybów następuje wraz z tworzeniem gamet, zarodników płciowych i innych form płciowych. Formy seksualne nazywane są teleomorfami.
Rozmnażanie bezpłciowe grzybów następuje wraz z tworzeniem odpowiednich form, zwanych anamorfami. Takie rozmnażanie zachodzi przez pączkowanie, fragmentację strzępek i zarodników bezpłciowych. Endogenne zarodniki (sporangiospory) dojrzewają wewnątrz zaokrąglonej struktury - zarodni. Egzogenne zarodniki (konidia) powstają na wierzchołkach strzępek owocujących, tzw. konidioforach.
Istnieją różne konidia. Arthroconidia (arthrospores) lub talloconidia powstają z równomierną przegrodą i rozcięciem strzępek, a blastoconidia powstają w wyniku pączkowania. Małe jednokomórkowe konidia nazywane są mikrokonidiami, duże wielokomórkowe konidia nazywane są makrokonidiami. Do form bezpłciowych grzybów należą również chlamydoconidia, czyli chlamydospory (grubościenne duże komórki spoczynkowe lub kompleks małych komórek).
Istnieją doskonałe i niedoskonałe grzyby. Grzyby doskonałe rozmnażają się płciowo; obejmują one zygomycetes (zygomykota), workowce ( Ascomycota ) i podstawczaki (podstawczaki). Niedoskonałe grzyby mają tylko aseksualny sposób hodowla; obejmują formalne typ warunkowy/ grupa grzybów - deuteromycetes (Deiteromycota).
Zygomycetes należą do niższe grzyby(grzybnia nie przegrodzona). Należą do nich przedstawiciele rodzaju Mucor, Rhizopus, Rhizomucor, Absidia, Basidiobolus, Conidiobolus. Rozprzestrzeniany w glebie i powietrzu. Mogą powodować zygomykozę (mukormykozę) płuc, mózgu i innych narządów człowieka.
Podczas rozmnażania bezpłciowego zygomycetes na owocującej strzępce (sporangioforze) powstaje zarodnia - kuliste zgrubienie z otoczką zawierającą liczne sporangiospory (ryc. 2.6, 2.7). Rozmnażanie płciowe w zygomycetes odbywa się za pomocą zygospor.
Ascomycetes (torbacze) mają grzybnię z przegrodami (z wyjątkiem drożdży jednokomórkowych). Wzięły swoją nazwę od głównego organu owocującego - worka lub woreczka, zawierającego 4 lub 8 haploidalnych zarodników płciowych (askospor).
Ascomycetes to poszczególni przedstawiciele (teleomorfy) rodzajów kropidlak I Penicillium. Większość rodzajów grzybów Aspergillus, Penicillium są anamorfami, tj. rozmnażać się tylko nieszkodliwie
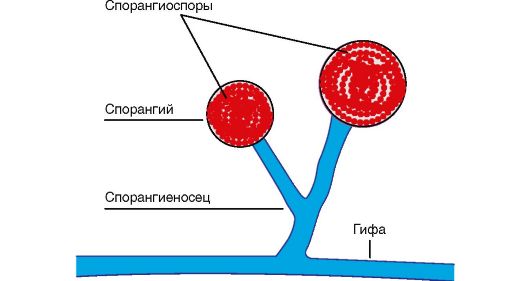 Ryż. 2.6. Grzyby z rodzaju Mukor(Fot. A.S. Bykow)
Ryż. 2.6. Grzyby z rodzaju Mukor(Fot. A.S. Bykow)
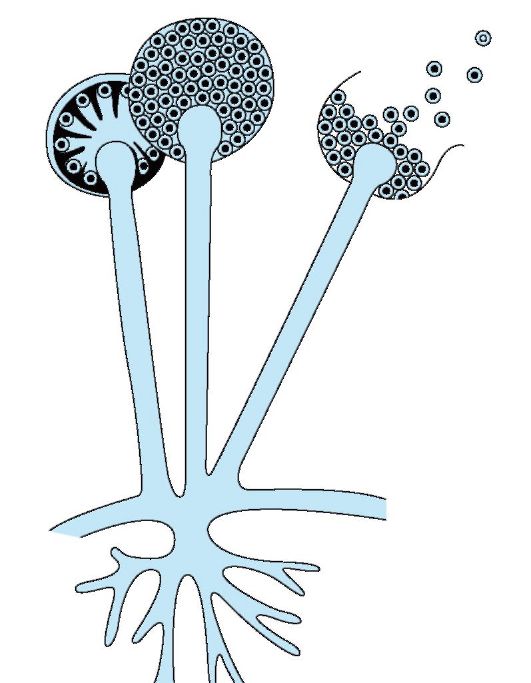 Ryż. 2.7. Grzyby z rodzaju Rhizopus Rozwój zarodni, sporangiospor i ryzoidów
Ryż. 2.7. Grzyby z rodzaju Rhizopus Rozwój zarodni, sporangiospor i ryzoidów
w sposób lym przy pomocy zarodników bezpłciowych - konidiów (ryc. 2.8, 2.9) i według tej cechy należy klasyfikować jako grzyby niedoskonałe. U grzybów z rodzaju kropidlak na końcach strzępek owocujących, konidioforów, znajdują się zgrubienia - sterigmy, fialidy, na których tworzą się łańcuchy konidiów ("pleśń lecha").
U grzybów z rodzaju Penicillium(racus) strzępka owocująca przypomina pędzel, ponieważ tworzą się z niej zgrubienia (na konidioforze), rozgałęziające się na mniejsze struktury - sterigmy, fialidy, na których znajdują się łańcuchy konidiów. Niektóre rodzaje aspergillus mogą powodować aspergilozę i aflatoksykozę, penicillium może powodować penicylozę.
Przedstawicielami workowców są teleomorfy rodzajów Trichophyton, Microsporum, Histoplasma, Blastomyces, a także drżenie
 Ryż. 2.8. Grzyby z rodzaju Penicillium.Łańcuchy konidiów rozciągają się od fialid
Ryż. 2.8. Grzyby z rodzaju Penicillium.Łańcuchy konidiów rozciągają się od fialid
 Ryż. 2.9. Grzyby z rodzaju Aspergillus fumigatus.Łańcuchy konidiów rozciągają się od fialid
Ryż. 2.9. Grzyby z rodzaju Aspergillus fumigatus.Łańcuchy konidiów rozciągają się od fialid
Basidiomycetes obejmują grzyby kapeluszowe. Mają grzybnię z przegrodami i tworzą zarodniki płciowe - basidiospory, odrywając się od podstawki - końcowej komórki grzybni, homologicznej do worka. Niektóre drożdże, takie jak teleomorfy, to podstawczaki. Cryptococcus neoformans.
Deuteromycetes to niedoskonałe grzyby (Grzyby niedoskonałe, grzyby anamorficzne, grzyby konidialne). Jest to warunkowy, formalny takson grzybów, łączący grzyby, które nie rozmnażają się płciowo. Ostatnio zamiast terminu „deuteromycetes” zaproponowano określenie „grzyby mitosporowe” – grzyby rozmnażające się przez zarodniki bezpłciowe, tj. przez mitozę. Ustalając fakt rozmnażania płciowego niedoskonałych grzybów, przenosi się je do jednego ze znanych typów - Ascomycota Lub podstawczak, podając nazwę formy teleomorficznej. Deuteromycetes mają grzybnię z przegrodami i rozmnażają się tylko przez bezpłciowe tworzenie konidiów. Deuteromycetes obejmują niedoskonałe drożdże (grzyby drożdżopodobne), na przykład niektóre grzyby z rodzaju Candida wpływając na skórę, błony śluzowe i narządy wewnętrzne(kandydoza). Są owalne, o średnicy 2-5 mikronów, dzielą się przez pączkowanie, tworzą pseudostrzępki (pseudomycelium) w postaci łańcuchów wydłużonych komórek, czasem tworzą strzępki. Dla Candida albicans charakterystyczne jest tworzenie się chlamydospor (ryc. 2.10). Deuteromycetes obejmują również inne grzyby, które nie mają płciowego trybu rozmnażania, spokrewnione z rodzajami Epidermophyton, Coccidioides, Paracoccidioides, Sporothrix, Aspergillus, Phialophora, Fonsecaea, Exophiala, Cladophialophora, Bipolaris, Exerohilum, Wangiella, Alrernaria itd.
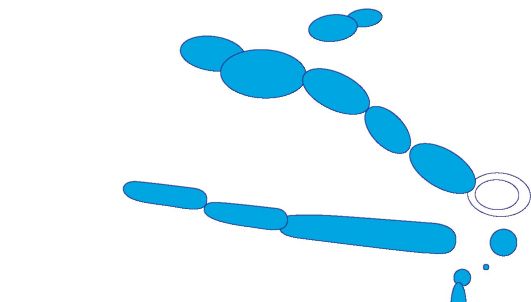 Ryż. 2.10. Grzyby z rodzaju Candida albicans(Fot. A.S. Bykow)
Ryż. 2.10. Grzyby z rodzaju Candida albicans(Fot. A.S. Bykow)
2.4. Budowa i klasyfikacja pierwotniaków
Najprostsze należą do domeny eukaria, Królestwo zwierząt (Zwierzęta) podkrólestwo pierwotniaki. Ostatnio zaproponowano wyróżnienie pierwotniaków do rangi królestwa pierwotniaki.
Komórka pierwotniaka jest otoczona błoną (błonką) - analogiem błony cytoplazmatycznej komórek zwierzęcych. Ma jądro z błoną jądrową i jąderkiem, cytoplazmę zawierającą retikulum endoplazmatyczne, mitochondria, lizosomy i rybosomy. Rozmiary pierwotniaków wahają się od 2 do 100 mikronów. Po zabarwieniu według Romanovsky'ego-Giemsy jądro pierwotniaka jest czerwone, a cytoplazma jest niebieska. Pierwotniaki poruszają się za pomocą wici, rzęsek lub pseudopodiów, niektóre z nich mają wakuole trawienne i kurczliwe (wydalnicze). Mogą żerować w wyniku fagocytozy lub tworzenia specjalnych struktur. Według rodzaju odżywiania są one podzielone na heterotrofy i autotrofy. Wiele pierwotniaków (ameby czerwonki, Giardia, Trichomonas, Leishmania, Balantidia) może rosnąć na pożywkach zawierających natywne białka i aminokwasy. Do ich hodowli wykorzystywane są również kultury komórkowe, zarodki kurze oraz zwierzęta laboratoryjne.
Rasa pierwotniaków bezpłciowo- podziały podwójne lub wielokrotne (schizogonia), a niektóre płciowe (sporogonia). Niektóre pierwotniaki rozmnażają się pozakomórkowo (Giardia), podczas gdy inne wewnątrzkomórkowo (Plasmodium, Toxoplasma, Leishmania). Cykl życiowy pierwotniaków charakteryzuje się etapami - powstawaniem stadium trofozoitu i stadium cysty. Cysty to stadia uśpione, odporne na zmiany temperatury i wilgotności. Cysty są kwasoodporne Sarcocystis, Cryptosporidium I Izospora.
Dawniej najprostsze chorobotwórczy u ludzi reprezentowane były przez 4 typy 1 ( Sarcomastigophora, Apicomplexa, Ciliophora, Microspora). Te typy zostały ostatnio przeklasyfikowane do większej liczby, pojawiły się nowe domeny - pierwotniaki I Chromista(Tabela 2.2). Do nowego królestwa Chromista(chromowiki) obejmowały niektóre pierwotniaki i grzyby (blastocysty, oomycetes i Rhinosporidium seeberi). Królestwo pierwotniaki obejmuje ameby, wiciowce, sporozoany i orzęski. Dzielą się na Różne rodzaje, wśród których są ameby, wiciowce, sporozoany i orzęski.
Tabela 2.2. przedstawiciele królestwa pierwotniaki I Chromista, o znaczeniu medycznym
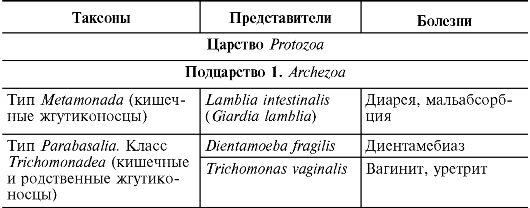 1 typ Sarcomastigophora składał się z podtypów Sarkodina I mastigofora. Podtyp Sarkodina(kod sarkodu) obejmował amebę czerwonki i podtyp mastigofora(wiciowce) - trypanosomy, leishmania, giardia i Trichomonas. Typ apikompleks zawiera klasę sporozoa(sporozoa), które obejmowały plazmodia malarii, toksoplazmę, cryptosporidium itp. Typ ciliofora obejmuje balantidia i typ mikrospora- mikrosporydia.
1 typ Sarcomastigophora składał się z podtypów Sarkodina I mastigofora. Podtyp Sarkodina(kod sarkodu) obejmował amebę czerwonki i podtyp mastigofora(wiciowce) - trypanosomy, leishmania, giardia i Trichomonas. Typ apikompleks zawiera klasę sporozoa(sporozoa), które obejmowały plazmodia malarii, toksoplazmę, cryptosporidium itp. Typ ciliofora obejmuje balantidia i typ mikrospora- mikrosporydia.
Koniec stołu. 2.2
 Ameby są czynnikiem sprawczym ludzkiej pełzakowicy - czerwonki amebowej (Entamoeba histolytica), wolno żyjące i niepatogenne ameby ( ameba jelitowa itd.). Ameby rozmnażają się binarnie bezpłciowo. Ich cykl życiowy składa się ze stadium trofozoitu (komórka rosnąca, ruchoma, niestabilna) oraz stadium cysty. Trofozoity poruszają się za pomocą pseudopodiów, które wychwytują i zanurzają składniki odżywcze w cytoplazmie. Z
Ameby są czynnikiem sprawczym ludzkiej pełzakowicy - czerwonki amebowej (Entamoeba histolytica), wolno żyjące i niepatogenne ameby ( ameba jelitowa itd.). Ameby rozmnażają się binarnie bezpłciowo. Ich cykl życiowy składa się ze stadium trofozoitu (komórka rosnąca, ruchoma, niestabilna) oraz stadium cysty. Trofozoity poruszają się za pomocą pseudopodiów, które wychwytują i zanurzają składniki odżywcze w cytoplazmie. Z
trofozoit, tworzy się cysta odporna na czynniki zewnętrzne. W jelicie zamienia się w trofozoit.
Wiciowce charakteryzują się obecnością wici: Leishmania ma jedną wici, Trichomonas ma 4 wolne wici i jedną wici połączoną z krótką falistą błoną. Oni są:
Wiciowce krwi i tkanek (leishmania - czynniki sprawcze leiszmaniozy; trypanosomy - czynniki sprawcze śpiączki i choroby Chagasa);
Wiciowce jelitowe (giardia - czynnik sprawczy lambliozy);
Wiciowce układu moczowo-płciowego (Trichomonas vaginalis - czynnik sprawczy rzęsistkowicy).
Rzęski są reprezentowane przez balantidia, które wpływają na ludzkie jelito grube (balantidiasis czerwonka). Balantidia mają stadium trofozoitu i cysty. Trofozoit jest ruchomy, ma liczne rzęski, cieńsze i krótsze niż wici.
2.5. Budowa i klasyfikacja wirusów
Wirusy to najmniejsze drobnoustroje należące do królestwa Virae(od łac. wirus- I). Oni nie mają struktura komórkowa i składać się
Strukturę wirusów, ze względu na ich niewielkie rozmiary, bada się za pomocą mikroskopii elektronowej zarówno wirionów, jak i ich ultracienkich skrawków. Wielkość wirusów (wirionów) określa się bezpośrednio za pomocą mikroskopii elektronowej lub pośrednio przez ultrafiltrację przez filtry o znanej średnicy porów, przez ultrawirowanie. Rozmiar wirusów waha się od 15 do 400 nm (1 nm to 1/1000 mikronów): małe wirusy, których rozmiar jest podobny do rozmiaru rybosomów, obejmują parwowirusy i wirusa polio, a największe wirusy ospa(350 nm). Wirusy różnią się formą wirionów, które mają postać pręcików (wirus mozaiki tytoniu), pocisków (wirus wścieklizny), kul (wirusy polio, HIV), włókien (filowirusy), plemników (wiele bakteriofagów).
Wirusy zadziwiają wyobraźnię swoją różnorodnością budowy i właściwości. W przeciwieństwie do genomów komórkowych, które zawierają jednorodny dwuniciowy DNA, genomy wirusowe są niezwykle zróżnicowane. Istnieją wirusy zawierające DNA i RNA, które są haploidalne, tj. mieć jeden zestaw genów. Tylko retrowirusy mają diploidalny genom. Genom wirusów zawiera od 6 do 200 genów i jest reprezentowany przez różne rodzaje kwasów nukleinowych: dwuniciowe, jednoniciowe, liniowe, koliste, fragmentaryczne.
Wśród wirusów zawierających jednoniciowy RNA wyróżnia się genomowy plusniciowy RNA i minusniciowy RNA (polarność RNA). Plus-nić (nić dodatnia) RNA tych wirusów, oprócz funkcji genomowej (dziedzicznej), pełni funkcję informacji, czyli macierzy RNA (mRNA lub mRNA); jest matrycą do syntezy białek na rybosomach zakażonej komórki. Plusniciowy RNA jest zakaźny: po wprowadzeniu do wrażliwych komórek może wywołać zakaźny pro-
cesja Nić ujemna (nić ujemna) wirusów zawierających RNA pełni jedynie funkcję dziedziczną; do syntezy białek, komplementarna nić jest syntetyzowana na ujemnej nici RNA. Niektóre wirusy mają ambipolarny genom RNA. (Atmosfera z greckiego ambi- po obu stronach, podwójna komplementarność), tj. zawiera plus i minus segmenty RNA.
Rozróżnia się wirusy proste (np. wirus zapalenia wątroby typu A) i wirusy złożone (np. grypy, opryszczki, koronawirusy).
Proste lub bezotoczkowe wirusy mają tylko kwas nukleinowy związany ze strukturą białkową zwaną kapsydem (od łac. kapsa- sprawa). Białka związane z kwasem nukleinowym są znane jako nukleoproteiny, a połączenie wirusowych białek kapsydu wirusa z wirusowym kwasem nukleinowym nazywa się nukleokapsydem. Niektóre proste wirusy mogą tworzyć kryształy (np. wirus pryszczycy).
Kapsyd zawiera powtarzające się podjednostki morfologiczne - kapsomery, złożone z kilku polipeptydów. Kwas nukleinowy wirionu wiąże się z kapsydem, tworząc nukleokapsyd. Kapsyd chroni kwas nukleinowy przed degradacją. W prostych wirusach kapsyd bierze udział w przyczepianiu (adsorpcji) do komórki gospodarza. Proste wirusy opuścić komórkę w wyniku jej zniszczenia (lizy).
Złożone lub otoczkowe wirusy (ryc. 2.11), oprócz kapsydu, mają podwójną powłokę lipoproteinową błony (synonim: superkapsyd lub peplos), która jest nabywana przez pączkowanie wirionu przez błonę komórkową, na przykład przez błona plazmatyczna, błona jądrowa lub błona retikulum endoplazmatycznego. Na otoczce wirusa znajdują się skoki glikoprotein,
lub kolce, popioły. Zniszczenie otoczki za pomocą eteru i innych rozpuszczalników dezaktywuje złożone wirusy. Pod powłoką niektórych wirusów znajduje się białko macierzy (białko M).
Wiriony mają helikalną, dwudziestościenną (sześcienną) lub złożoną symetrię kapsydu (nukleokapsydu). Helikalny typ symetrii wynika ze spiralnej struktury nukleokapsydu (na przykład w wirusach grypy, koronawirusach): kapsomery są ułożone spiralnie wraz z kwasem nukleinowym. Dwudziestościenny typ symetrii wynika z tworzenia się izometrycznego wydrążonego korpusu z kapsydu zawierającego wirusowy kwas nukleinowy (na przykład w wirusie opryszczki).
Kapsyd i otoczka (superkapsyd) chronią wiriony przed wpływami środowiska, determinują selektywne oddziaływanie (adsorpcję) ich białek receptorowych z określonym
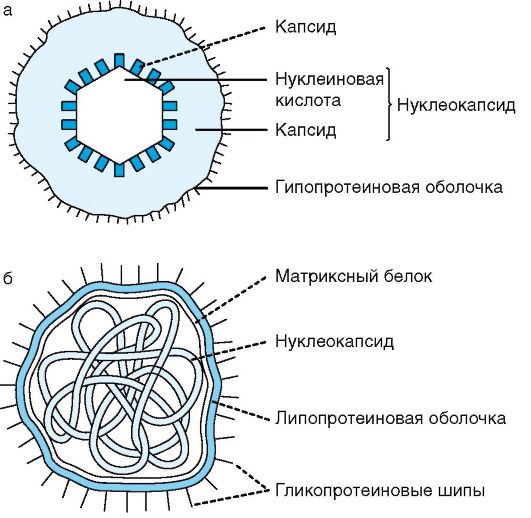 Ryż. 2.11. Struktura wirusów otoczkowych z dwudziestościennym (a) i helikalnym (b) kapsydem
Ryż. 2.11. Struktura wirusów otoczkowych z dwudziestościennym (a) i helikalnym (b) kapsydem
komórek, a także właściwości antygenowe i immunogenne wirionów.
Wewnętrzne struktury wirusów nazywane są rdzeniem. W adenowirusach rdzeń składa się z białek histonopodobnych związanych z DNA, w reowirusach składa się z białek kapsydu wewnętrznego.
Laureat nagroda Nobla D. Baltimore zaproponował system klasyfikacji Baltimore oparty na mechanizmie syntezy mRNA. Ta klasyfikacja dzieli wirusy na 7 grup (Tabela 2.3). Międzynarodowy Komitet Taksonomii Wirusów (ICTV) przyjęli uniwersalny system klasyfikacji, który wykorzystuje kategorie taksonomiczne, takie jak rodzina (nazwa kończy się na wirusowate), podrodzina (nazwa kończy się na wirusy), rodzaj (nazwa kończy się na wirus). Typ wirusa nie otrzymał nazwy dwumianowej, jak w przypadku bakterii. Wirusy klasyfikuje się ze względu na rodzaj kwasu nukleinowego (DNA lub RNA), jego budowę oraz liczbę nici. Mają dwuniciowe lub jednoniciowe kwasy nukleinowe; dodatnia (+), ujemna (-) polarność kwasu nukleinowego lub mieszana polarność kwasu nukleinowego, ambipolarna (+, -); liniowy lub kolisty kwas nukleinowy; fragmentowany lub niefragmentowany kwas nukleinowy. Wielkość i morfologia wirionów, liczba kapsomerów i rodzaj symetrii nukleokapsydu, obecność otoczki (superkapsydu), wrażliwość na eter i dezoksycholan, miejsce rozmnażania w komórce, właściwości antygenowe itp. uwzględnić.
Tabela 2.3. Główne wirusy o znaczeniu medycznym
 Kontynuacja tabeli. 2.3
Kontynuacja tabeli. 2.3
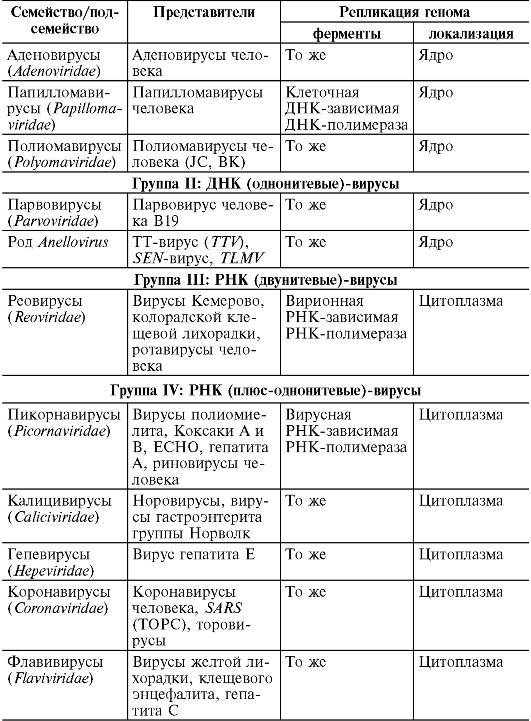 Koniec stołu. 2.3
Koniec stołu. 2.3
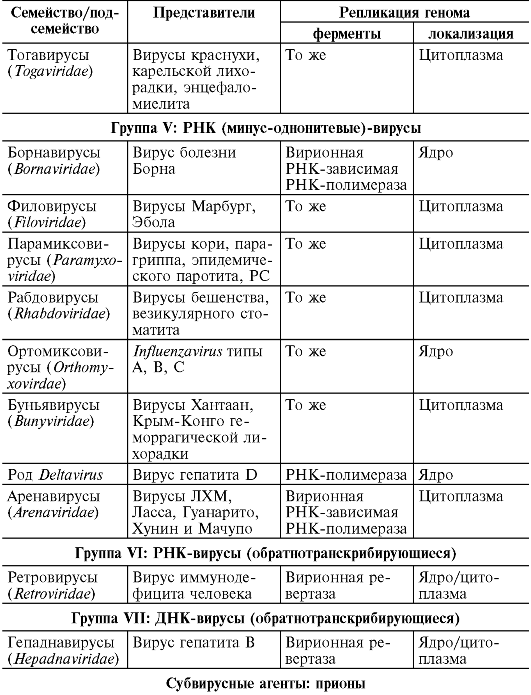 Wirusy infekują zwierzęta, bakterie, grzyby i rośliny. Będąc głównymi czynnikami sprawczymi chorób zakaźnych człowieka, wirusy biorą również udział w procesach karcynogenezy, mogą być przenoszone różne sposoby, w tym przez łożysko (wirus różyczki, wirus cytomegalii itp.), wpływając na ludzki płód. Mogą również prowadzić do powikłań poinfekcyjnych - rozwoju zapalenia mięśnia sercowego, zapalenia trzustki, niedoboru odporności itp.
Wirusy infekują zwierzęta, bakterie, grzyby i rośliny. Będąc głównymi czynnikami sprawczymi chorób zakaźnych człowieka, wirusy biorą również udział w procesach karcynogenezy, mogą być przenoszone różne sposoby, w tym przez łożysko (wirus różyczki, wirus cytomegalii itp.), wpływając na ludzki płód. Mogą również prowadzić do powikłań poinfekcyjnych - rozwoju zapalenia mięśnia sercowego, zapalenia trzustki, niedoboru odporności itp.
Pozakomórkowe formy życia, oprócz wirusów, obejmują priony i wiroidy. Wiroidy to małe cząsteczki okrągłego, superskręconego RNA, które nie zawierają białka i powodują choroby roślin. Patologiczne priony to zakaźne cząsteczki białek, które powodują szczególne choroby konformacyjne w wyniku zmiany struktury normalnego komórkowego białka prionowego ( PrP c), który znajduje się w organizmie zwierząt i ludzi. PrP z pełni funkcje regulacyjne. Jest kodowany przez normalny gen prionu (gen PrP) znajdujący się na krótkim ramieniu 20. ludzkiego chromosomu. Choroby prionowe są przenoszone gąbczaste encefalopatie(choroba Creutzfeldta-Jakoba, kuru itp.). W tym przypadku białko prionowe nabywa inne, postać zakaźna, oznaczone jako PrP sc(sc z trzęsawka- scrapie - infekcja prionowa owiec i kóz). To zakaźne białko prionowe jest podobne do fibryli i różni się od normalnego białka prionowego strukturą trzeciorzędową lub czwartorzędową.
Zadania do samokształcenia (samokontrola)
A. Wymień drobnoustroje, które są prokariotami:
2. Wirusy.
3. Bakterie.
4. Priony.
B. Podaj cechy charakterystyczne komórki prokariotycznej:
1. Rybosomy 70S.
2. Obecność peptydoglikanu w ścianie komórkowej.
3. Obecność mitochondriów.
4. Diploidalny zestaw genów.
W. Wymień składniki peptydoglikanu:
1. Kwasy teichojowe.
2. N-acetyloglukozamina.
3. Lipopolisaryd.
4. Tetrapeptyd.
G. Zwróć uwagę na cechy strukturalne ściany komórkowej bakterii Gram-ujemnych:
1. Kwas mezodiaminopimelinowy.
2. Kwasy teichojowe.
4. Białka porinowe.
D. Wymień funkcje zarodników u bakterii:
1. Zapisz widok.
2. Odporność na ciepło.
3. Osiadanie podłoża.
4. Powielanie.
1. Rickettsia.
2. promieniowce.
3. Krętki.
4. Chlamydia.
I. Wymień cechy promieniowców:
1. Mają termolabilne zarodniki.
2. Bakterie Gram-dodatnie.
3. Nie ma ściany komórkowej.
4. Miej skręcony kształt.
Z. Wymień cechy krętków:
1. Bakterie Gram-ujemne.
2. Mają motoryczny aparat włóknisty.
3. Mają skręcony kształt.
I. Wymień pierwotniaki, które mają kompleks wierzchołkowy, który umożliwia im penetrację wnętrza komórki:
1. Plasmodium malarii.
3. Toksoplazma.
4. Cryptosporidium.
DO. nazwa cecha wyróżniająca złożone wirusy:
1. Dwa rodzaje kwasu nukleinowego.
2. Obecność błony lipidowej.
3. Podwójny kapsyd.
4. Obecność białek niestrukturalnych. Ł. Zaznacz wyższe grzyby:
1. Mukor.
2. Candida.
3. Penicillium.
4. kropidlak.
Bakterie to mikroorganizmy, które można zobaczyć tylko pod mikroskopem, większość z nich jest jednokomórkowa i ruchliwa dzięki obecności wici.
Kształt komórki jest różny u różnych gatunków. Bardziej powszechne są bakterie, które można przypisać następującym formom: kuliste - ziarniaki; prosty w kształcie pręta - prątki i bakterie; wygięty w kształcie pręta - vibrios; spiralnie skręcony - spirilla. Komórka bakteryjna składa się z błony zawierającej protoplazmę, a czasem wakuole. Przy odpowiednim wybarwieniu protoplazmy można znaleźć ciała, które przypominają jądra, ale być może nie odpowiadają jądrom komórkowym innych organizmów. Większość bakterii jest bezbarwna, niektóre mają czerwone i zielone pigmenty.
Wiele bakterii, zwłaszcza pałeczkowate i niektóre kuliste, tworzy zarodniki (endospory), które rozwijają się w zgrubiałej części treści protoplazmatycznej i są zaopatrzone na ich powierzchni w gęstą, trudną do penetracji otoczkę. Ta otoczka daje zarodnikom większą odporność na niesprzyjające warunki. warunki zewnętrzne i zachowują żywotność przez bardzo długi czas. Na korzystne warunki zarodniki rosną, rozwijając się w nową komórkę wegetatywną.
Bakterie rozmnażają się przez poprzeczny podział komórki na dwie części, a przerwa między dwoma podziałami wynosi często tylko pół godziny, dlatego znacznie przewyższają wszystkie inne organizmy szybkością rozmnażania.
Większość bakterii ma odżywianie heterotroficzne, to znaczy zaspokaja swoje potrzeby kosztem gotowej materii organicznej, a tylko nieliczne charakteryzują się odżywianiem autotroficznym kosztem węgla w dwutlenku węgla. Jeśli chodzi o odżywianie azotem, niektóre z tych drobnoustrojów wymagają azotu organicznego, podczas gdy większość może zawierać azot amonowy i azotanowy, a niektóre są w stanie przyswoić azot elementarny.
Większość bakterii to organizmy tlenowe, niektóre mogą rozwijać się tylko przy wystarczającej ilości tlenu w środowisku, ale są również znane bakterie beztlenowe; nie potrzebują tlenu. Są niezwykle powszechne, zwłaszcza tam, gdzie występują materia organiczna, które niszczą, tworząc różnego rodzaju zgniliznę lub fermentację. Wiele drobnoustrojów pasożytuje w organizmach roślinnych lub zwierzęcych. Wiele jest czynników sprawczych chorób zakaźnych, na przykład gruźlicy, duru brzusznego, cholery, dżumy, błonicy. Inne zapewniają bardzo ważne procesy w życiu człowieka ( Różne rodzaje fermentacja, akumulacja azotu w glebie bakterie guzkowe), nie mówiąc już o tym, że bez ich udziału proces powstawania gleby i obieg substancji w przyrodzie są nie do pomyślenia. Niektóre mikroorganizmy mają bezpośrednie znaczenie w medycynie, na przykład te, z których są silne
Bakterie wyróżniają się następującymi cechami:
- morfologiczny;
- barwiarski;
- fizjologiczny;
- kulturalny;
- Biochemiczne;
- antygenowy;
- genetyka molekularna.
Cechy morfologiczne
Bakterie - głównie. Budowa komórki prokariotycznej znacznie różni się od budowy komórek grzybów, roślin i zwierząt. Nie mają błony jądrowej, mitochondriów, aparatu Golgiego. W komórce bakteryjnej wyróżnia się następujące elementy:
- struktury powierzchniowe;
- Błona komórkowa;
- cytoplazma z nukleotydem.
Struktury powierzchniowe nazywane są formacjami zlokalizowanymi poza błoną cytoplazmatyczną. Należą do nich kapsułka, warstwa śluzowa, otoczka, ściana komórkowa, wici, kosmki.
Kapsułka, otoczka lub warstwa śluzu jest wymagana do ochrony bakterii przed wpływami środowiska. Chronią przed różnymi substancje chemiczne, chronią przed wysychaniem, stanowią przeszkodę w penetracji fagów. Bakterie bytujące w żołądku są chronione przez specjalną substancję śluzową, która chroni komórkę przed działaniem kwasu nadchlorowego, enzymów soku żołądkowego.
Wici określają zdolność poruszania się. Ich liczba, lokalizacja jest stała, dlatego są brane pod uwagę przy systematyzowaniu prokariontów.
Mogą być zlokalizowane na biegunach lub w obszarze polarnym komórki bakteryjnej. Według dostępności lokalizacja dzieli się na:
- monotrichous - jedna wici znajduje się na biegunie komórki (cholera vibrio);
- lophotrichous - wiązka wici umieszczona z jednej strony (Pseudomonas aeruginosa);
- amphitrichous - wiązki wici znajdują się na różnych biegunach komórki bakteryjnej (spirilla);
- peritrichous - wici rozmieszczone na całej powierzchni komórki (salmonella).
Ściana komórkowa znajduje się bezpośrednio pod torebką, warstwą śluzu lub otoczką.
Wewnątrz komórki bakteryjnej znajduje się cytoplazma, organelle i nukleotydy. Od innych prokariotów, do których należy królestwo sinic, różnią się brakiem chlorofilu.
Bakterie są klasyfikowane według ich kształtu. Mogą być kuliste (ziarniaki, diplokoki, paciorkowce, gronkowce), cylindryczne (pręty), spiralne (krętki, spirilla, vibrios).
Znaki barwiące (stosunek do specjalnych barwników)
Metodę Ziehla-Nelsona stosuje się do wykrywania kwasoopornych i alkoholoodpornych Mycobacterium tuberculosis, czynników wywołujących trąd, bakterii przyczyniających się do rozwoju wrzodów żołądka.
Metoda srebrzenia Morozowa służy do izolowania krętków i inkluzji wirusowych.
Barwnik Romanowskiego-Giemsy służy do identyfikacji bakterii, grzybów i pierwotniaków. Do barwienia stosuje się złożony barwnik, który barwi różne rodzaje mikroorganizmów w różnych kolorach.
Znaki fizjologiczne
Objawy fizjologiczne obejmują:
- Biochemiczne;
- kulturalny.
Znaki biochemiczne
Rodzaje bakterii rozróżnia się na podstawie parametrów biochemicznych (wydzielane enzymy, odżywianie, oddychanie). Na przykład Helicobacter pylori, który żyje w żołądku, jest w stanie wydzielać sekret, który stymuluje pojawianie się wrzodów. Dlatego, gdy pojawiają się objawy wrzodu, zaleca się badanie bakteryjne. W chorobach wywołanych infekcją, zwłaszcza u dzieci, dochodzi do zatrucia toksynami uwalnianymi przez mikroorganizmy. Do niektórych dodaje się enzymy bakteryjne proszki do prania. Stosowany do otrzymywania związków kwasu octowego, mlekowego.
Szczególne znaczenie w określaniu infekcji ma rodzaj oddychania. Wyróżnić:
- bezwzględne bakterie beztlenowe. Tlen jest dla nich toksyczny. Do oddychania potrzebują azotanów, siarczanów lub fumaranów. Żyją w przestrzeni pozbawionej powietrza. Wpływa na żołądek, jelita, układ moczowo-płciowy. Szczególnie niebezpieczne są infekcje powodujące zatrucie jadem kiełbasianym, tężec, zgorzel gazową.
- obowiązkowe aeroby. Mogą istnieć tylko w obecności tlenu. W związku z tym są czynnikami sprawczymi chorób Układ oddechowy, choroby skóry u dorosłych i dzieci.
- fakultatywne beztlenowce. Należą do nich niektóre rodzaje bakterii i grzybów, które nie potrzebują tlenu do wzrostu i rozmnażania.
Według rodzaju odżywiania bakterie dzielą się na:
- autotrofy;
- heterotrofy.
Niektóre bakterie są normalna mikroflora Ludzkie ciało. W takim przypadku choroby mogą być spowodowane spadkiem liczby pożyteczne bakterie. Tak więc przyjmowanie antybiotyków zmniejsza ilość E. coli, powodując zaburzenia żołądka lub jelit. Saprofity nie pozwalają się rozwijać mikroorganizmy chorobotwórcze, wraz ze spadkiem liczby bakterii, mogą rozpocząć się choroby wywołane przez patogenne gatunki grzybów.
znaki kulturowe
W zależności od rodzaju odżywiania, siedliska, bakterie wyróżniają się cechami kulturowymi.
Aby określić, do jakiego rodzaju mikroorganizmów należą, wykonuje się siew. Do badań bakteryjnych można wykorzystać nie tylko cząsteczki skóry, ślinę, plwocinę, ale także zawartość żołądka.
Na przykład przy objawach zapalenia migdałków u dzieci i dorosłych obowiązkowe jest badanie mikrobiologiczne. Usunąć sterylnym wacikiem biała powłoka na gardle. Posadzone w specjalnych szalkach Petriego. Następnie przyglądają się, na jakiej pożywce mikroorganizmy namnażały się najlepiej, gdzie się znajdują (rozproszone pojedyncze punkty lub skupione w jednym miejscu). Duże znaczenie ma również kolor i wielkość kolonii bakteryjnej.
Po zidentyfikowaniu rodzaju infekcji, która wywołała chorobę u dzieci i dorosłych, analizują oporność szczepu na antybiotyki. Następnie przepisuje się odpowiednie leczenie.
Cechy antygenowe
Określ rodzaj hodowli bakteryjnej, wykonując badanie krwi. W końcu w przypadku choroby spowodowanej infekcją nasz organizm wydziela przeciwciała przeciwko patogenowi. Po zidentyfikowaniu ich obecności ustala się szczep mikroorganizmów, który spowodował chorobę.
Aby określić rodzaj infekcji, która spowodowała chorobę, wykonuje się badanie surowicy krwi na obecność przeciwciał lub antygenu - reakcje serologiczne. Oni przywykli:
- na toksyny, antygen przy użyciu surowicy immunodiagnostycznej;
- w celu wykrycia przeciwciała przy użyciu znanego antygenu.
W przypadkach podejrzenia stosuje się analizę serologiczną ukryta forma choroba. Zbadaj osoby, które miały kontakt z osobą zarażoną, aby wykryć chorobę we wczesnym stadium.
Czasami osoba może sama nie zachorować, ale zostać nosicielem. Lub choroba spowodowana infekcją przebiega bezobjawowo, jest przewlekła.
Aby zdiagnozować gruźlicę, podaje się dzieciom test skórny- Test Mantoux. Do jego wykonania stosuje się suchą tuberkulinę (wyciąg z resztek bakteryjnych). Zaczerwienienie, obrzęk skóry obserwuje się, jeśli ciało dziecka ma przeciwciała przeciwko różdżce Kocha. Powstają tylko w obecności infekcji.
Molekularne cechy genetyczne
Podstawą różnicowania mikroorganizmów według cech genetycznych jest fundamentalna różnica w budowie genów. Tak więc w wirusach i fagach materiał dziedziczny jest zamknięty w 1 cząsteczce DNA lub RNA. U pierwotniaków i grzybów chromosomy znajdują się w Jądro komórkowe. Aparat genetyczny bakterii różni się tym, że:
- Chromosom znajduje się w cytoplazmie. Składa się z 3000 genów.
- Zawartość DNA zmienia się, może wzrosnąć. Ta unikalna zdolność reguluje tempo rozmnażania, co ma pozytywny wpływ na przetrwanie gatunku.
- Informacje dziedziczne zawarte są w genach chromosomu i plazmidach.
- Przekazywanie informacji odbywa się pionowo (z komórki macierzystej do komórki potomnej), poziomo (bakterie są w stanie wymieniać między sobą geny). W ten sposób zmutowane geny są przenoszone z jednej komórki bakteryjnej do drugiej. Na przykład jeden miał pozytywny wynik mutacja genów które zapewniały antybiotykooporność infekcji. Podczas wymiany informacji z innymi bakteriami inna kolonia otrzyma tę samą zdolność.
Kiedy pojawiają się objawy choroby, jeśli istnieje podejrzenie, że jest to spowodowane przez bakterie, konieczne jest przeprowadzenie badania mającego na celu zróżnicowanie czynników sprawczych choroby, określenie oporności na antybiotyki.
Jeśli zostanie zidentyfikowany zakaźny charakter choroby, należy pamiętać, że bakterie mogą powodować epidemie. Z powodu słaba odporność choroba zakaźna najczęściej chorują dzieci. Aby zapobiec rozprzestrzenianiu się choroby, podejmuje się szereg działań:
- Jeśli w szkołach lub przedszkolach wystąpi kilka przypadków zachorowań, aby nie narażać innych dzieci, placówki są zamykane na czas kwarantanny.
- Dla zapobiegania poważna choroba, co może prowadzić do śmiertelny wynik zalecił szczepienie dzieci.
- W czasie epidemii nie zaleca się uczestniczenia w imprezach towarzyskich.
- Dzieci i dorośli, zwłaszcza w okresie sezonowych chorób, muszą przyjmować leki immunostymulujące.
Podczas diagnozowania chorób spowodowanych infekcją należy wziąć pod uwagę główne. Różnią się budową, kształtem komórek, rodzajem odżywiania i oddychania. Dyrygowanie badania mikrobiologiczne, konieczne jest wykonanie barwienia Grama, zaszczepienie na pożywce (określenie cech kulturowych). Czasami przepisywany jest test serologiczny (badanie surowicy krwi określa obecność antygenu lub przeciwciał). Prowadzone jest również badanie genetyki molekularnej, które opiera się na różnicach w budowie i umiejscowieniu nukleotydu.