Citologija yra mokslas, tiriantis ląstelių sąveiką ir ląstelių struktūrą, kuri, savo ruožtu, yra pagrindinė bet kurio gyvo organizmo sudedamoji dalis. Pats terminas kilęs iš senovės graikų sąvokų „kitos“ ir „logos“, reiškiančių atitinkamai narvą ir doktriną.
Mokslo atsiradimas ir ankstyvoji raida
Citologija yra viena iš visos mokslo galaktikos, kuri šiais laikais atsiskyrė nuo biologijos. Jo atsiradimo pirmtakas buvo mikroskopo išradimas XVII a. Stebėdamas gyvybę per tokią primityviausią konstrukciją, anglas pirmą kartą atrado, kad viskas susideda iš ląstelių, todėl jis padėjo pamatus šiandieninėms citologijos studijoms. Po dešimties metų kitas mokslininkas – Anthony Leeuwenhoekas – atrado, kad ląstelės turi griežtai sutvarkytą struktūrą ir veikimo modelius. Jam taip pat priklauso branduolių egzistavimo atradimas. Tačiau vis tiek ilgam laikui suprasti ląstelę ir jos funkcionavimą trukdė nepatenkinama to meto mikroskopų kokybė. Kiti svarbūs žingsniai buvo žengti XIX amžiaus viduryje. Tada technika buvo žymiai patobulinta, o tai leido sukurti naujas koncepcijas, dėl kurių citologija yra intensyviai vystoma. Tai visų pirma protoplazmos atradimas ir atsiradimas
 Ląstelių teorijos atsiradimas
Ląstelių teorijos atsiradimas
Remdamiesi iki tol sukauptomis empirinėmis žiniomis, biologai M. Schleidenas ir T. Schwannas beveik vienu metu mokslo pasauliui pasiūlė mintį, kad visos gyvūnų ir augalų ląstelės yra panašios viena į kitą, o kiekviena tokia ląstelė pati savaime turi visas savybes. ir gyvo organizmo funkcijas . Ši komplekso samprata gyvybės formų planetoje turėjo didelę įtaką citologijos keliui. Tai taip pat taikoma šiuolaikinei plėtrai.
Protoplazmos atradimas
Kitas svarbus laimėjimas minėtoje žinių srityje buvo protoplazmos savybių atradimas ir aprašymas. Tai medžiaga, kuri užpildo ląsteliniai organizmai, taip pat yra terpė ląstelių organams. Vėliau mokslininkų žinios apie šią medžiagą išsivystė. Šiandien ji vadinama citoplazma.
Tolesnis genetinio paveldėjimo vystymas ir atradimas
 XIX amžiaus antroje pusėje buvo aptikti atskiri kūnai, esantys Jie buvo vadinami chromosomomis. Jų tyrimas atskleidė žmonijai genetinio tęstinumo dėsnius. Didžiausią indėlį į šią sritį XIX amžiaus pabaigoje įnešė austras Gregoras Mendelis.
XIX amžiaus antroje pusėje buvo aptikti atskiri kūnai, esantys Jie buvo vadinami chromosomomis. Jų tyrimas atskleidė žmonijai genetinio tęstinumo dėsnius. Didžiausią indėlį į šią sritį XIX amžiaus pabaigoje įnešė austras Gregoras Mendelis.
Dabartinė mokslo padėtis
Šiuolaikinei mokslo bendruomenei citologija yra viena iš svarbiausių šakų biologines žinias. Mokslinės metodologijos ir techninių galimybių plėtra ją padarė tokią. Šiuolaikinės citologijos metodai plačiai naudojami žmonėms naudinguose tyrimuose, pavyzdžiui, tyrime vėžinis navikas, dirbtinių organų auginimas, taip pat veisimas, genetika, naujų gyvūnų ir augalų rūšių veisimas ir kt.
ląstelių mokslo struktūrinės ir funkciniai vienetai beveik visi gyvi organizmai. Daugialąsčiame organizme visos sudėtingos gyvybės apraiškos atsiranda dėl suderintos jį sudarančių ląstelių veiklos. Citologo užduotis yra nustatyti, kaip gyva ląstelė ir kaip jis atlieka įprastas funkcijas. Patologai taip pat tiria ląsteles, tačiau juos domina pokyčiai, atsirandantys ląstelėse ligos metu ar po mirties. Nepaisant to, kad mokslininkai jau seniai sukaupė daug duomenų apie gyvūnų ir augalų raidą bei sandarą, tik 1839 metais buvo suformuluotos pagrindinės ląstelių teorijos sąvokos ir pradėta kurti šiuolaikinė citologija.
Ląstelės yra mažiausi gyvybės vienetai, tai liudija audinių gebėjimas suskaidyti į ląsteles, kurios vėliau gali toliau gyventi „audine“ arba ląstelių kultūroje ir daugintis kaip mažyčiai organizmai. Pagal ląstelių teoriją visi organizmai susideda iš vienos ar kelių ląstelių. Yra keletas šios taisyklės išimčių. Pavyzdžiui, gleivinių pelėsių (miksomicetų) kūne ir kai kurių labai mažų plokščiųjų kirmėlių ląstelės nėra atskirtos viena nuo kitos, o sudaro daugiau ar mažiau susiliejusią struktūrą, vadinamą. sincitiumas. Tačiau galima daryti prielaidą, kad tokia struktūra antrą kartą atsirado sunaikinus ląstelių membranų dalis, kurios buvo šių organizmų evoliuciniuose protėviuose. Daugelis grybų auga ilguose siūliniuose vamzdeliuose arba hifuose. Šie hifai, dažnai pertvaromis suskirstyti į segmentus, taip pat gali būti laikomi savotiškais pailgos ląstelės. Protistų ir bakterijų kūnai susideda iš vienos ląstelės.
Tarp bakterijų ląstelių ir visų kitų organizmų ląstelių yra viena svarbus skirtumas: bakterijų ląstelių branduoliai ir organelės („mažieji organai“) nėra apsupti membranų, todėl šios ląstelės vadinamos prokariotinėmis („ikibranduolinėmis“); visos kitos ląstelės vadinamos eukariotinėmis (su „tikraisiais branduoliais“): jų branduoliai ir organelės yra uždarytos membranose. Šiame straipsnyje aptariamos tik eukariotinės ląstelės. taip pat žr .
Tačiau labiausiai svarbi programa citologiniai metodai medicinoje yra diagnozė piktybiniai navikai. IN vėžio ląstelės, ypač jų branduoliuose, yra specifinių pakitimų, kuriuos pripažįsta patyrę patologai. taip pat žr VĖŽYS.
Čencovas Y.S. . Bendroji citologija, 3 leidimas M., 1995 m
Green N., Stout W., Taylor D. Biologija, t. 1. M., 1996 m
MOLEKULINIS GENETINIS IR LĄSTELIŲ LYGIS
GYVENIMO ORGANIZACIJOS KAIP ORGANIZMO GYVENIMO PAGRINDAS
CITOLOGIJOS PAGRINDAI
Citologija– biologijos šaka, šiuo metu veikianti kaip savarankiškas mokslas, tiriantis struktūrinius, funkcinius ir genetinės savybės visų organizmų ląstelės.
Šiuo metu citologiniai tyrimai turi reikšmingas ligų diagnostikai, nes jos leidžia tirti patologiją remiantis elementariu gyvosios medžiagos sandaros, funkcionavimo ir dauginimosi vienetu - ląstelės. Ląstelių lygmenyje pasireiškia visos pagrindinės gyvo būtybės savybės: medžiagų apykaita, biologinės informacijos panaudojimas, dauginimasis, augimas, dirglumas, paveldimumas, gebėjimas prisitaikyti. Gyvų organizmų ląstelės išsiskiria įvairia morfologija ir struktūriniu sudėtingumu (net tame pačiame organizme), tačiau tam tikros savybės randamos visose be išimties ląstelėse.
atidarymas ląstelių organizacija prieš gyvas būtybes buvo išrasti didinamieji instrumentai. Taigi pirmąjį mikroskopą būtų sukonstravę olandų optikai Hansas ir Zachary Jansenai (1590). Didysis Galilėjus Galilėjus padarė mikroskopą 1612 m. Tačiau ląstelės tyrimo pradžia laikomi 1665 metai, kai anglų fizikas Robertas Huygensas panaudojo savo tautiečio Christiano Huygenso išradimą (1659 m. sukonstravo okuliarą), pritaikydamas jį tyrimams mikroskopu. smulki struktūra eismo kamščiai. Jis pastebėjo, kad kamštienos medžiaga susideda iš daugybės mažų ertmių, atskirtų viena nuo kitos sienelėmis, kurias pavadino ląstelėmis. Tai buvo mikroskopinių tyrimų pradžia.
Ypač pažymėtini A. Leeuwenhoek tyrimai, kurie 1696 metais atrado vienaląsčių organizmų (bakterijų ir blakstienų) pasaulį ir pirmą kartą pamatė gyvūnų ląsteles (eritrocitus ir spermatozoidus).
1825 metais J. Purkinje pirmą kartą pastebėjo branduolį vištienos kiaušinyje, o T. Schwann pirmasis aprašė branduolį gyvūnų ląstelėse.
Iki 19 amžiaus 30-ųjų buvo sukaupta reikšminga faktinė medžiaga mikroskopinė struktūra ląstelių ir 1838 m. M. Schleidenas iškėlė tapatybės idėją augalų ląstelės jų raidos požiūriu. T. Schwann padarė galutinį apibendrinimą, suprasdamas ląstelės reikšmę ir ląstelių struktūra kaip pagrindinė gyvųjų organizmų gyvybės ir vystymosi struktūra.
ląstelių teorija, sukurtas M. Schleiden ir T. Schwann, teigia, kad ląstelės yra gyvų būtybių struktūrinis ir funkcinis pagrindas. R. Virchow pritaikė Schleiden-Schwann ląstelių teoriją in medicininė patologija, papildant jį tokiomis svarbiomis nuostatomis kaip „kiekviena ląstelė iš ląstelės“ ir „kiekvienas skausmingas pokytis yra susijęs su kai kuriais patologinis procesas ląstelėse, sudarančiose kūną.
Pagrindinės nuostatos šiuolaikinės ląstelių teorija:
1. Ląstelė – elementarus visų gyvų organizmų sandaros, funkcionavimo, dauginimosi ir vystymosi vienetas, už ląstelės ribų gyvybės nėra.
2. Ląstelė yra vientisa sistema, turinti didelis skaičius tarpusavyje sujungti elementai – organelės.
3. Ląstelės įvairūs organizmai panašios (homologinės) struktūros ir pagrindinių savybių ir turi bendrą kilmę.
4. Ląstelių skaičiaus padidėjimas vyksta joms dalijantis, po jų DNR replikacijos: ląstelė – iš ląstelės.
5. Daugialąstis organizmas yra nauja sistema, sudėtingas daugybės ląstelių, sujungtų ir integruotų į audinių ir organų sistemas, sujungtų cheminiais veiksniais: humoraliniais ir nerviniais, visuma.
6. Ląstelės daugialąsčiai organizmai totipotentinė – bet kuri daugialąsčio organizmo ląstelė turi tą patį bendrą šio organizmo genetinės medžiagos fondą, galimos galiosšios medžiagos pasireiškimui – tačiau skiriasi atskirų genų raiškos (darbo) lygiu, o tai lemia jų morfologinę ir funkcinę įvairovę – diferenciaciją.
Taigi, ląstelių teorijos dėka, organinės gamtos vienybės idėja yra pagrįsta.
Šiuolaikiniai citologijos tyrimai:
Ląstelių sandara, jų kaip elementarios gyvos sistemos funkcionavimas;
Atskirų ląstelių komponentų funkcijos;
Ląstelių dauginimosi procesai, jų taisymas;
Prisitaikymas prie aplinkos sąlygų;
Specializuotų ląstelių ypatybės.
Citologiniai tyrimai yra būtini diagnozuojant žmonių ligas.
Pagrindiniai žodžiai ir sąvokos: citologija, ląstelė, ląstelių teorija
BENDRA INFORMACIJA APIE LĄSTELES
Visos žinomos gyvybės formos Žemėje gali būti klasifikuojamos taip:
NE LĄSTELĖS GYVYBĖS FORMOS
VIRUSAI
Virusas (lot. virusas- nuodai) - neląstelinis organizmas, kurio dydis svyruoja nuo 20 iki 300 nm.
Virionai (virusinės dalelės) susideda iš dviejų arba trys komponentai: viruso šerdis yra genetinė medžiaga DNR arba RNR pavidalu (kai kurios turi abiejų tipų molekules), aplink yra baltyminis apvalkalas (kapsidas), suformuotas iš subvienetų (kapsomerų). Kai kuriais atvejais atsiranda papildomas lipoproteinų apvalkalas plazmos membrana savininkas. Kiekviename viruse kapsido kapsomerai yra griežtai išdėstyti tam tikra tvarka, dėl kurių atsiranda ypatingas simetrijos tipas, pavyzdžiui, spiralinė (vamzdinė forma – tabako mozaikos virusas arba sferinė RNR turinčiuose gyvūnų virusuose) ir kubinė (izometriniai virusai) arba mišri (1 pav.).
Ryžiai. 1 Virusų sandaros schema: A - vamzdinis (lazdelės formos) spiralinės struktūros virusas: 1 - baltymai; 2 - nukleorūgštis. B - daugiakampis su mišrus tipas simetrija (bakteriofagas): 1 - galva su nukleino rūgštimi; 2 - uodegos procesas; 3 - bazinė plokštelė; 4 - proceso gijos.
Viruso gyvavimo ciklas apima keletą etapų: prisitvirtinimo prie ląstelės paviršiaus (kai kurie virusai, pvz., ŽIV, užkrečia tik tam tikro tipo ląsteles); įsiskverbimas į ląstelę įvairiomis strategijomis ir jos genetinės medžiagos „įšvirkštimas“.
Viruso gebėjimas sukelti ligą apibūdinamas terminu virulentiškumas. Dauguma ląstelių, susilietus su virusu, miršta dėl lizės. Daugialąsčiuose organizmuose, kai miršta daug ląstelių, visas organizmas kenčia dėl homeostazės pažeidimo, kuris sukelia ligas. Tačiau atskiri virusai gali egzistuoti kūno viduje ir yra gana nekenksmingi delsos laikas(herpeso virusai). Šioje būsenoje virusai gali būti netgi naudingi, nes jų buvimas gali sukelti imuninį atsaką prieš bakterinius patogenus. Tais atvejais, kai virusai sukelia visą gyvenimą trunkantį ir lėtinės infekcijos(hepatitas B ir C), sergantys žmonės veikia kaip užkrečiamo viruso rezervuaras. Jei nešiotojų dalis populiacijoje yra didelė, sako jie epidemijos.
Visus viruso perdavimo būdus galima sąlygiškai suskirstyti į du tipus:
Vertikalus viruso perdavimas – iš motinos vaikui (hepatitas B, ŽIV);
Horizontaliai nuo žmogaus iki žmogaus: lytinio akto metu pernešant kūno skysčius, pavyzdžiui, užsikrėtus ŽIV; per kraują, pavyzdžiui, užsikrėtus hepatito C virusu; seilių perdavimas Epstein-Barr virusas; nurijus užteršto vandens ar maisto – norovirusas; įkvepiant orą, kuriame jie yra virionai(virusinės dalelės) – pvz., gripo virusas; vabzdžių.
Perdavimo greitis virusinė infekcija priklauso nuo tokių veiksnių kaip gyventojų tankumas, žmogaus imuniteto būklė, sveikatos priežiūros kokybė ir kt.
Nemažai virusų yra patogeniški žmogui: tai raupų virusų šeima (sukelia daugybę raupų infekcijų), herpeso grupės virusai (lūpų pūslelinės, vėjaraupiai), adenovirusai (ligos kvėpavimo takai ir akis), papovavirusų šeima (karpos ir kiti odos dariniai), hepadnavirusai (hepatito B virusas); pikornavirusai (poliomielitas, hepatitas A, ūminis peršalimo); miksovirusai ir paramiksovirusai (priežastis skirtingos formos gripo, tymų ir kiaulytės(kiaulės)); arbovirusai (nešioja nariuotakojai - kai kurių encefalito, geltonosios karštinės ir kitų pavojingų ligų sukėlėjai); reovirusai yra reti kvėpavimo takų ir žarnyno ligos asmuo.
Atskirai diskusijai reikalingas žmogaus imunodeficito virusas (ŽIV), užkrečiantis vieną iš limfocitų formų – T pagalbininkus, aktyvuojančius patogenus naikinančius T žudikus. įvairių ligų sukelia AIDS (įgyto imunodeficito sindromą). ŽIV būdingas didelis kintamumas (maždaug penkis kartus didesnis nei gripo viruso ir šimtą kartų didesnis nei hepatito B viruso), o tai apsunkina vakcinos gavimo problemą ir apsunkina specifinę AIDS prevenciją. ŽIV perdavimo būdai: lytinis kontaktas, naudojimas nesterilus medicinos instrumentai, kuriuos dažnai vartoja narkomanai; galimas infekcijos perdavimas per kraują ir kai kurios vaistai, persodinant organus ir audinius; infekcija gali pasireikšti nėštumo metu, vaiko gimimo metu ar jo metu maitinimas krūtimi motina, užsikrėtusi ŽIV ar AIDS. AIDS būdingas labai ilgas inkubacinis periodas(nuo pralaimėjimo momento iki pirmųjų ligos požymių atsiradimo) – vidutiniškai 5 metai, per kuriuos užsikrėtę žmonės gali užkrėsti kitus žmones. Viena iš žmogaus užsikrėtimo AIDS virusu apraiškų yra centrinio nugalėjimas nervų sistema.
Ilgą laiką buvo galvojama apie virusus PRIONAI- infekcinės baltymų molekulės, kuriose nėra RNR ar DNR ir kurios priklauso toms pačioms priešląstelinėms gyvybės formoms kaip ir virusai. Prionai gali skatinti savo kopijų susidarymą. Priono forma, sąveikaudama su normaliu baltymu, skatina jo transformaciją į priono formą.
Bijūnų atradimas yra susijęs su vadinamųjų lėtųjų infekcijų doktrinos, kurioms būdinga:
Neįprastai ilgas (mėnesių ir metų) inkubacinis laikotarpis;
Lėtai progresuojantis kurso pobūdis;
Neįprasti organų ir audinių pažeidimai;
Mirties neišvengiamybė.
Šios ypatingos lėtųjų infekcijų grupės patomorfologija labai skiriasi ryškiu originalumu, kuris pasireiškia tik centrinės nervų sistemos pralaimėjimu. Ligų vystymosi mechanizmas yra toks: remiantis pirminiais degeneraciniais procesais, kurie vyksta be uždegimo požymių, jis vystosi. būdingas vaizdas kempinę primenančios pilkos būsenos susidarymas arba baltoji medžiaga galva, o kartais nugaros smegenys- vadinamoji užkrečiamoji spongiforminė encefalopatija (USE).
Šiuo metu prioninės ligos yra gana retos. Jų pasitaiko vienas milijonui per metus, tačiau kai kuriuose regionuose (Slovakijoje, Izraelyje, Čilėje) dažnis yra daug didesnis. Žmogaus prioninės ligos apima vištieną, Creutzfeldt-Jakob ligą, Gerstmann-Straussler-Scheinker sindromą ir mirtiną šeimyninę nemigą. Žmonėms šios ligos yra reta kančia.
Pagrindiniai žodžiai ir sąvokos: virusas, kapsidas, kapsomeras, bakteriofagas, virulentiškumas, latentinis laikotarpis, epidemija, virionas, ŽIV, AIDS, prionai.
LĄSTELĖS GYVYBĖS FORMOS
PROKARIOTAI
prokariotai – ikibranduoliniai organizmai, kurios ląstelėje nėra susiformavusio, membranomis apriboto branduolio. Prokariotams priskiriama apie 3 tūkstančius rūšių: spirochetų, eubakterijų, miksobakterijų, melsvadumblių, archebakterijų, riketsijų, mikoplazmų, aktinomicetų ir nemažai kitų organizmų, kurių sisteminė padėtis neaiški, todėl tikroji jų įvairovė gali siekti apie 100 tūkstančių rūšių.
Pagrindiniai prokariotų ląstelių struktūros bruožai (2 pav.):
Ryžiai. 2. Statinio schema bakterinė ląstelė: 1 - gleivinė kapsulė, 2 - ląstelių sienelės, 3 - citoplazminė membrana, 4 - citozolis, 5 - nukleoidas, 6 - ribosomos, 7 - mezosomos, 8 - pūslelės, 9 - tilakoidai, 10 - glikogeno granulės, 11 - aliejaus lašai, 12 - volutino grūdeliai, 13 - sieros lašai, 14 - žvyneliai.
- ląstelių sienelės bakterijos turi standžią struktūrą, kuri lemia ląstelės formą, apsaugą ir kontaktą su išorinė aplinka; ląstelių sienelės skirtingos bakterijos nevienodai nudažyta pagal gramą: gramteigiamų bakterijų ląstelės sienelė yra 20–80 nm storio ir 90 % sudaro mureino peptidoglikanas, anatomiškai vienalytis; gramneigiamose bakterijose ląstelės sienelė yra apie 10 m storio, joje yra 1–10 % mureino ir sluoksninės struktūros;
Vidinis bakterinės ląstelės turinys ( protoplastas) yra pripildytas pusiau skystos koloidinės masės – citozolio ir apsuptas plazmolema, kuris gali formuoti invaginacijas į ląstelę, mezosomos veikiantis kaip visos ląstelės membranos organelės ir turėti skirtinga forma: kanalėlių- vamzdinis pūslelės- burbulo forma tilakoidai- lamelinis; ląstelės protoplaste yra ribosomos- ne membranos organelės, kuriose vyksta baltymų biosintezė;
Vienintelė, dažniau žiedinė prokariotų chromosoma ( nukleoidas) atlieka branduolio vaidmenį, kuris nuo citoplazmos nėra atskirtas membranomis;
Bakterijų ląstelėse gali būti mažų genetinių struktūrų - plazmidės galintys savarankiškai pakartoti ir turėti guolius Papildoma informacija;
Jei prokariotinėje ląstelėje yra inkliuzų, jie arba atlieka rezervinę funkciją, arba yra atstovaujami bakterinės ląstelės medžiagų apykaitos produktų; atstovaujama polisacharidų grūdeliais, aliejaus lašais, siera;
Flagella, pili, blakstiena atlieka motorinę funkciją (flagellum), prisitvirtina prie substrato arba dalyvauja plazmidžių pernešimo iš ląstelės į ląstelę procese konjugacijos metu (pili).
Morfologiškai bakterijos skiriasi forma, dydžiu, santykinė padėtis ląstelės, žvynelių ir kapsulių buvimas ar nebuvimas, gebėjimas sporuliuoti ir kt.
Pagal formą bakterijų ląstelės skirstomos į (3 pav.):
rutulinės bakterijos– cocci- neturi žvynelių ir nesudaro sporų; sudaro kelių tipų mikrokolonijas:
Mikrokokai - dalijasi toje pačioje plokštumoje, po padalijimo išsidėsto pavieniui;
Diplokokai – po ląstelių dalijimosi išsidėsto poromis;
T etrakokai– ląstelės dalijasi dviejose viena kitai statmenose plokštumose, susidaro 4 ląstelių grupės;
SU treptokokai - ląstelės dalijasi vienoje plokštumoje, po dalijimosi ląstelės lieka grandinėmis;
- Sarcinai - ląstelės dalijasi į tris viena kitai statmenas plokštumas, susidaro 8 arba 64 ląstelių paketai;
SU tafilokokai - ląstelės dalijasi į neapibrėžtos kryptys, sudaro ląstelių sankaupą, primenančią vynuogių kekes;
lazdelės formos bakterijos - gausiausia ir įvairiausia bakterijų grupė, kurios morfologiškai skiriasi ląstelės dydžiu, jos galų forma, žvynelių buvimu ar nebuvimu, taip pat gebėjimu sporuliuoti. Dažniausiai jie skirstomi į pogrupius:
A) bakterijos- lazdelės formos (nesudaro sporų, ląstelių dalijimasis yra skersinis); gali būti sujungtos dvi ląstelės - diplobakterijos ir grandinėje streptobakterijos;
b) bacilos - strypo formos formos, galinčios Ne palankiomis sąlygomis sukelti ginčus; pagal ląstelių santykinę padėtį diplobacilos Ir streptobacilos;

3 pav. Bakterijų morfologija (paaiškinimai tekste)
susuktos bakterijos- išlenktos lazdos, kurios skiriasi kreivumo laipsniu:
vibrijos– trumpos lazdelės, 1-3 mikronų ilgio, sulenktos per pusę bangos ilgio, savo forma primena kablelį;
spirilla– 15-20 mikronų ilgio strypai, išlenkti iki viso bangos ilgio, primenantys ištemptą lotynišką raidę S;
spirochetos– plonos ilgos ląstelės, 20 - 30 mikronų, su daugybe vingių, primena išplėstą spiralę, turi išilginį ląstelių dalijimąsi.
Morfologiškai bakterijos taip pat skiriasi pagal žvynelinės vietos pobūdį:
monotiškas - turėti vieną poliarinį žvynelį;
lopotrichas - bakterijos turi vieną žvynelių ryšulį;
amfitricai - du žvynelių ryšuliai, esantys priešinguose poliuose;
P eritras - visas bakterijos ląstelės paviršius padengtas daugybe žvynelių.
Prokariotiniai organizmai dauginasi dalijantis ląstelėms per pusę, kartais pumpuruojant, retai konjuguojant. Jie dauginasi milžinišku greičiu. Taigi, esant palankioms sąlygoms, jų ląstelės dalijasi kas 20-30 minučių. Todėl jie gali greitai padidinti savo skaičių per trumpą laiką. Esant nepalankioms sąlygoms, bakterijos ląstelės paviršiuje susidaro tankus daugiasluoksnis apvalkalas. Visi gyvybės procesai ląstelėje yra sustabdyti, ji nesidalija. Taip susiformuoja ginčas. Prokariotinė ląstelė gali gyventi kaip sporos ilgas laikas, jis atlaiko aukštą arba žemos temperatūros, sausra. Esant palankioms sąlygoms, sporų apvalkalas sunaikinamas, ląstelėje vėl atsinaujina gyvybiniai procesai.
Prokariotinės bakterijos yra visur. Jie nusėda paviršiuje arba kitų organizmų (žmonių, gyvūnų, augalų) viduje, daug aptinkami dirvožemyje, gėlo ir sūraus vandens telkiniuose. Pavyzdžiui, tik viename grame dirvožemio yra milijonas bakterijų ląstelių. Puiki suma jų yra vandens arba atmosferos oro tūrio vienete. Daugelis jų yra patogeniški ir sukelia rimtų sveikatos problemų, vystymosi sutrikimų ir mirtį.
Bakterijų sukeliamų infekcinių ligų spektras yra labai įvairus, tačiau kiekvienos iš jų paplitimo laipsnis ir sunkumas paprastai priklauso nuo trijų veiksnių:
Pačių bakterijų savybės
Makroorganizmo (žmogaus) būklė ligos metu;
Organo, kuriame daugiausia vystosi patologinis procesas, ypatybės.
Galimas bakterijų gebėjimas sukelti infekcines ligas, kuri yra jų rūšiai būdinga, vadinama sergamumas, arba patogeniškumas. Yra žinoma, kad tos pačios rūšies bakterijose patogeninių savybių pasireiškimo laipsnis gali labai skirtis. Virulentiškumasįvardykite padermės patogeniškumo laipsnį tam tikros rūšies bakterijos.
Bakterijos sukelia daugybę ligų. Pavyzdžiui, streptokokai sukelia anginą, pneumokokai dažnai sukelia vidurinės ausies uždegimą; mikobakterijos sukelia tuberkuliozę; meningokokai prisideda prie galvos ir (arba) nugaros smegenų membranų uždegimo (meningito) atsiradimo. Kiti įsidėmėtini bakterinės infekcijos: stabligė, juodligė, šiltinė, cholera ir maras. Bakterijos sukelia ir kai kurias vaikų ligas, tokias kaip kokliušas, skarlatina, difterija.
Pagrindiniai žodžiai ir sąvokos: prokariotai, nukleoidas, plazmidės, kokos, mikrokokai, diplokokai, tetrakokai, streptokokai, sarkinai, stafilokokai, lazdelės formos bakterijos, bakterijos, diplobakterijos, streptobakterijos, bacilos, vibrionai, spiriliai, spirochetai, patogeniškumas.
EUKARIOTAI
būdingas bruožas eukariotai yra branduolio, kuris gali dalytis, buvimas ir seksualinis procesas.
Organus, turinčius eukariotinį organizacijos tipą, sąlygiškai galima suskirstyti į du potipius: pirmasis būdingas paprasčiausiams organizmams, antrasis – daugialąsčiams organizmams.
PROTOTIAI
Organizacijos bruožas pirmuonys yra tai, kad jie (išskyrus kolonijines formas) struktūriškai atitinka vienos ląstelės lygį, o fiziologiškai – visavertį individą. Paprasčiausių dydžiai svyruoja nuo 2 iki 50 mikronų ir daugiau.

Ryžiai. 4. Pirmuonių įvairovė: 1 - blakstieninis batas; 2 - žalia euglena; 3 - tripanosoma; 4 - lamblija; 5 - opalas; 6 - radiolarinis; 7 - ameba proteus; 8 - dizenterinė ameba; 9 - Trichomonas; 10 - volvoksas.
Pirmuonių morfologija (5 pav.). Pirmuonių kūnas sudarytas iš protoplazma, šerdis ir įvairūs intarpai. Protoplazma turi ląstelinę struktūrą ir joje, net ir labiausiai organizuotuose pirmuoniuose, išskiriami du sluoksniai: vidinis - endoplazma, kuriame yra branduolys ir kiti intarpai, o išorinis, išorinis, tankesnis ir skaidresnis sluoksnis – ektoplazma, kuri skirta judėti, gaudyti maistą ir apsaugoti pirmuonis.
Pirmuonių judėjimas Skirtingi keliai: šliaužti palei substratą pseudopodų, žvynelių ir blakstienų pagalba.
Pirmuonių branduolys yra panašus į kitų gyvūnų ląstelių branduolius. Daugeliu atvejų pirmuonys turi vieną branduolį, tačiau blakstienas turi du – makro ir mikrobranduolių. Kai kurie pirmuonys turi kelis branduolius. Branduolys gali būti burbulo formos (amebose ir daugelyje kitų pirmuonių) ir masyvus (blakstienų makrobranduolys).
Pirmuonys dauginasi nelytiškai ir lytiškai. At nelytinis dauginimasis pirma, jų branduolys dalijamas į dvi ar daugiau dalių, o vėliau citoplazma dalijama į dvi (lygias arba nelygias) arba daug (pagal naujai susidariusių branduolių skaičių) dalių. Dėl to iš vieno organizmo atsiranda du (vienodo ar nevienodo dydžio) arba keli nauji padarai. Lytinio dauginimosi metu du vienodi arba skirtingo dydžio ir struktūros individai (patinas ir patelė) susilieja vienas su kitu į vieną individą – zigotą, kuris vėliau pradeda daugintis. aseksualiai. Kartais du šio tipo pirmuonių individai tik liečia vienas kitą, keisdami dalį savo branduolių. 
5 pav. Bato blakstienų struktūra: 1 - blakstiena, 2 - virškinimo vakuolės, 3 - didelis branduolys (makrobranduolis), 4 - mažas branduolys (mikronranduolis), 5 - burnos anga ir ryklės, 6 - nesuvirškintų likučių išmestas maistas, 7 - trichocistos, 8 - susitraukiančios vakuolės
Daugelis pirmuonių, būdami nepalankiomis gyvenimo sąlygomis, virsta cistomis, o jų kūnas yra suapvalintas ir padengtas storu apvalkalu. Šioje būsenoje pirmuonys gali būti ilgą laiką, kol atsidurs palankesnėse jų egzistavimo sąlygose. Tada gyvūnas išeina iš cistos apvalkalo ir pradeda vesti judantis vaizdas gyvenimą.
At žarnyno forma amebiazė viršutinė dalis Storojoje žarnoje atsiranda uždegimo židinys, susidaro opos, kartais net audinių nekrozė. Autorius kraujagyslės amebos gali prasiskverbti į kepenis, sudarydamos ten antrinius židinius – pūlinius; V pleuros ertmė ir plaučiai, perikardas, pilvo ertmė; gali metastazuoti į smegenis, odą ir kitus organus.
Giardiaze dažniau serga vaikai (ypač dažnai nuo 1 iki 4 metų). Simptomai: skausmas viršutinėje pilvo dalyje arba bamboje, pilvo pūtimas, ūžesys, pykinimas, prakaitavimas, vėliau viduriavimas (geltonos išmatos su nedideliu gleivių kiekiu), diskinezija. tulžies takų, atopinis dermatitas, bendras silpnumas, nuovargis, dirglumas, apetito praradimas, galvos skausmai, Blogas sapnas. Kartais liga praeina be ryškių apraiškų ir paprastai nustatoma po kokios nors kitos ligos. Vaiko svorio augimas sulėtėja.
Toksoplazmozė- toksoplazmozės sukelta liga, kurios šaltiniai yra Skirtingos rūšys(virš 180) naminių ir laukinių žinduolių (katės, šunys, triušiai ir kt.).
Trichomonozė - infekcija, lytiniu keliu plintančią, sukelia vienaląstis mikroorganizmas – Trichomonas. Trichomonozė yra pirmoji pagal paplitimą tarp urogenitalinių takų ligų ir lytiniu keliu plintančių ligų. ši liga slypi tame, kad 70–80% vyrų ligų yra besimptomės ir pasireiškia tik komplikacijų stadijoje. Moterims liga dažniausiai pasireiškia per pirmąsias 1–4 savaites. Trichomonozė yra pavojinga pirmiausia dėl sunkių pasekmių komplikacijų, galinčių sukelti nevaisingumą, forma. Moteriai ligą galima nustatyti nėštumo metu – ji turi įtakos ne tik augimui, bet ir vaisiaus vystymuisi. Be to, urogenitalinė trichomonozė gali sukelti persileidimą, priešlaikinis gimdymas ar paties nėštumo komplikacijų.
Pagrindiniai žodžiai ir sąvokos: eukariotai, pirmuonys, protoplazma, endoplazma, ektoplazma, maliarija, amebiazė, toksoplazmozė, trichomonozė.
BENDROSIOS CHARAKTERISTIKOS
DAUGALASĖS ORGANIZMO GYVŪNŲ LĄSTELĖS STRUKTŪROS
biologinės membranos
Bet kurios ląstelės vidinės organizacijos tvarkingumo pagrindas yra skirstymas jo turinys – padalijimas į „ląsteles“, kuris vyksta dėl biologinių membranų, kurios turi vieną struktūrą visoms be išimties ląstelėms ir jų organelėms (6 pav.).

Ryžiai. 6. Molekulinė organizacija biologinė membrana: lipidų sluoksnis: 1 - hidrofilinė "galva", 2 - hidrofobinė "uodega": baltymų molekulės: 3 - paviršinė, 4 - periferinė (panardinta), 5 - integrali (skvarbi).
Šiuo metu yra visuotinai pripažintas 1971 m. Nicholson ir Singer pasiūlytas skysčio mozaikinis biologinės membranos modelis, pagal kurį jo struktūros pagrindas yra dvigubas lipidų sluoksnis (bilipidų sluoksnis). Iš esmės membranos lipidus atstovauja fosfolipidai. Lipidai turi amfifiliškumo (arba amfipatiškumo) savybę, nes savo sudėtyje turi polinę hidrofilinę „galvą“ ir nepolinę hidrofobinę „uodegą“, kurių ilgis vienas su kitu yra maždaug 1:4. Dėl šios struktūros in vandens aplinka lipidai linkę išsidėstyti taip, kad nepolinės uodegos liestųsi viena su kita, o polinės „galvutės“ – su vandeniu, todėl lipidai hidrofilinėje aplinkoje spontaniškai sudaro dvigubą sluoksnį (dvisluoksnį): hidrofobines sritis. yra viduje, o hidrofiliniai regionai yra išorėje. Ši lipidų molekulių organizacija atitinka būseną su mažiausia vertė Gibbso potencialas. Susidaręs bilipido sluoksnis nesudaro kraštų, bet linkęs užsidaryti ant savęs. Pavyzdžiui, susidaro liposomos.
Membranose yra daug įvairių baltymų. Membraninių baltymų koncentracija priklauso nuo ląstelės tipo. Pavyzdžiui, mielininiame aksono apvalkale baltymų yra 2,5 karto mažiau nei lipidų, o eritrocitų membranoje – priešingai – 2,5 karto daugiau baltymų. Vieni baltymai išsidėstę membranos paviršiuje (paviršiaus baltymai), kiti iš dalies panardinami į lipidų sluoksnį arba pro membraną prasiskverbia (periferiniai ir integraliniai baltymai).
Integraliniai baltymai, kaip ir lipidai, turi amfifiliškumo savybę: ta baltymo dalis, kuri yra membranoje, susideda iš hidrofobinių aminorūgščių, dažniausiai susisukusių į spiralę; kita dalis, išsikišusi iš membranos, yra sudaryta iš hidrofilinių aminorūgščių. Integralūs baltymai membranoje išlaikomi dėl hidrofobinių sąveikų, o be jų kartais dar ir citoskeleto molekulės – mikrovamzdeliai ar mikrofilamentai. Angliavandeniai gali būti prijungti prie vientiso baltymo galo, išsikišusio į tarpląstelinę aplinką. Šie kompleksai, vadinami glikoproteinais, dažnai yra receptoriai ir žaidžia svarbus vaidmuo V imuninės reakcijos organizmas. Integralinių baltymų pavyzdžiai yra fotoreceptorių membranų baltymas – bakteriorodopsinas, septynis kartus prasiskverbiantis pro membraną, eritrocitų baltymas – glikoforinas, prie kurio polipeptidinės grandinės gali būti prijungti įvairūs oligosacharidai; šie glikoproteinai lemia AB0 sistemos kraujo grupę. Daugelis vientisų baltymų sudaro kanalus, kuriais gali praeiti vandenyje tirpios molekulės ir jonai. Tokiu atveju vidinė dalis kanale yra hidrofilinių aminorūgščių radikalų.
Periferiniai baltymai gali būti tiek su vidiniais, tiek su lauke membranos. Paprastai jie yra susieti su membrana elektrostatinės sąveikos būdu, tai yra daug silpnesni nei integraliniai baltymai, todėl daugelis periferinių baltymų yra lengvai pašalinami iš membranos.
Storis biologinės membranos yra 5-10 nm, tačiau nepaisant to, jų dalis sausoje ląstelių masėje viršija 50%. Taip yra dėl tankios membranos komponentų pakuotės, taip pat dėl didelio bendro citoplazminių ir tarpląstelinių membranų ploto.
Visos natūralios membranos yra asimetriškos, tai yra, identiškos lipidų molekulės yra išorinėje ir vidiniai paviršiaiįvairios koncentracijos membranos. Šių dviejų paviršių lipidai, kaip taisyklė, skiriasi savo hidrofilinėmis galvutėmis. Asimetrijos savybė taip pat būdinga membraninių baltymų išsidėstymui. Pavyzdžiui, membraninių glikolipidų angliavandenių komponentai randami tik už ląstelės ribų. Membranų asimetrija dažnai prisideda prie to, kad medžiagos gabenamos tik viena kryptimi.
Ląstelių membranos atlieka nemažai funkcijų: barjerą (riboja), reguliuoja ir užtikrina selektyvų medžiagų pralaidumą, sudaro sąsajas tarp vandeninės (hidrofilinės) ir nevandeninės (hidrofobinės) fazės su fermentų kompleksų išdėstymu ant šių paviršių. Membranų molekulinė sudėtis, jų paviršiuje esančių junginių ir jonų rinkinys skiriasi priklausomai nuo struktūros. Taip pasiekiama funkcinė ląstelių membranų specializacija. Dėl receptorių molekulių įtraukimo į ląstelės membraną ji tampa jautri biologiškai aktyviems junginiams, tokiems kaip hormonai. Ant membranų surenkami organoidai (žr. „Golgi aparatas“) – tai sintetinė membranų funkcija. Impulsų (sužadinimo) perdavimas ląstelėje ir organizme taip pat vyksta dalyvaujant membranų kompleksams.
Ląstelių sienelės(citoplazminė membrana arba plazmolema)
Gyvūnų ląstelių plazmalemą sudaro membrana, kurios išorėje yra 10–20 nm storio glikokalikso sluoksnis. Jį sudaro membraniniai baltymai, glikolipidų angliavandenių dalys ir glikoproteinai. Glikokaliksas atlieka svarbų vaidmenį receptorių funkcijoje, užtikrina ląstelės „individualizavimą“ – joje yra audinių suderinamumo receptorių.
Iš vidaus prie membranos ribojasi 0,1-0,5 mikrono storio žievės (žievės) citoplazmos sluoksnis, kuriame ribosomų ir pūslelių nerandama, o reikšminga suma yra mikrovamzdeliai ir mikrofilamentai, kuriuose yra susitraukiančių baltymų.
Lauke ląstelės membrana reguliuoja nuolatinį medžiagų apykaitą tarp ląstelės ir aplinką. Molekulės praeina per membranas trimis skirtingais procesais: paprasta difuzija, palengvinta difuzija ir aktyviu transportavimu. Praleisdamos vandenį, ląstelių membranos nepraleidžia daugumos jame ištirpusių medžiagų. Tokios membranos vadinamos pusiau pralaidžiomis arba pasirinktinai pralaidžiomis. Vandens difuzija per pusiau pralaidžias membranas vadinama osmosu. Nepolinės (hidrofobinės) lipiduose tirpios medžiagos prasiskverbia pro membraną paprastos difuzijos būdu (įskaitant deguonį). Tai pasyvaus transportavimo pavyzdys, kurio kryptį lemia tik medžiagos koncentracijos skirtumas abiejose membranos pusėse. Dauguma ląstelei reikalingų medžiagų yra polinės ir per membraną pernešamos joje panardintų transportinių baltymų-nešiklių pagalba. Yra dvi baltymų transportavimo formos: palengvinta difuzija ir aktyvus transportavimas. Supaprastinta difuzija atsiranda dėl koncentracijos gradiento (koncentracijos skirtumo), o molekulės juda pagal šį gradientą. Nei paprasta, nei palengvinta difuzija negali prieštarauti koncentracijos gradientui. Tai yra dviejų rūšių pasyvus medžiagų transportavimas.
Tirpalų transportavimas prieš koncentracijos gradientą reikalauja energijos ir yra vadinamas aktyvus transportas. Viena iš labiausiai ištirtų aktyvaus transporto sistemų yra natrio-kalio pompa(8 pav.). Dauguma gyvūnų ir žmogaus ląstelių išlaiko skirtingus natrio ir kalio jonų koncentracijos gradientus – ląstelės viduje išlieka maža Na + ir didelė K + jonų koncentracija. Tai būtina norint palaikyti ląstelių tūrį (osmoreguliaciją), palaikyti nervo elektrinį aktyvumą ir raumenų ląstelės, taip pat aktyviam kitų medžiagų, tokių kaip aminorūgštys ir cukrus, pernešimui. Didelės koncentracijos kalio jonai taip pat reikalingi baltymų sintezei ir kitiems svarbiems procesams.
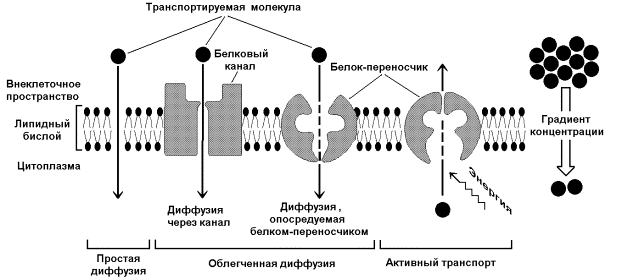
Ryžiai. 7. Transmembraninio transportavimo rūšys

Ryžiai. 8. Natrio-kalio siurblio schema
Per kitus aktyvius procesus per membraną gali būti pernešamos įvairios medžiagos endocitozė(vežimas į narvą) ir egzocitozė(išvežti iš kameros). Endocitozės metu medžiagos patenka į ląstelę dėl plazminės membranos invaginacijos, tada susidariusios pūslelės su turiniu yra atskiriamos nuo plazminės membranos ir perkeliamos į citoplazmą. Panašiai vyksta medžiagų išsiskyrimas iš ląstelės (egzocitozė), tik membrana formuoja išsikišimus.
Tankiųjų dalelių, tokių kaip bakterijos, gaudymas vadinamas fagocitozė. Daug vienaląsčiai organizmai(pavyzdžiui, ameba) maitinti tokiu būdu. Tirpiųjų medžiagų absorbcija vadinama pinocitozė, kuris randamas ir vienaląsčiuose, ir daugialąsčiuose organizmuose. Nors fagocitozė ir pinocitozė iš pirmo žvilgsnio skiriasi nuo membranų transportavimo sistemų, kuriose dalyvauja nešiklio molekulės, jų pagrindas yra tas pats. Visi šie mechanizmai priklauso nuo membranos gebėjimo „atpažinti“ tam tikras molekules.
Protoplazma
Protoplazma atstovaujamos ląstelės šerdis Ir citoplazma. Kita vertus, citoplazmoje yra hialoplazma- citoplazmos skystis, į kurį jie panardinami organelės.
Viskas po elektroniniu mikroskopu hialoplazma turi smulkiagrūdę struktūrą; tai sudėtinga koloidinė sistema, galinti pereiti iš zolinės (skystos) būsenos į gelį (tokių perėjimų procese dirbama). cheminiai junginiai, įskaitant fermentus, o tai rodo jo vaidmenį ląstelės biocheminėje veikloje. Hialoplazma jungia visas ląstelių struktūras ir užtikrina jų sąveiką.Matricos vienijančios ir karkasinės funkcijos gali būti siejamos su mikrotrabekuliniu tinklu, kurį sudaro plonos 2-3 nm storio fibrilės, prasiskverbiančios į visą citoplazmą.
ląstelės branduolys
Šerdis- reikalingas komponentas eukariotinės ląstelės(9 pav.). Ląstelėse, kurios dauginasi dalijantis, įprasta atskirti dvi morfologines branduolio būsenas - tarpfazę (intervale tarp dalijimosi) ir dalijimąsi. Branduolio forma skirtingos ląstelės taip pat gali skirtis: sferinis, elipsinis, pasagos ir kt. Daugumoje žmogaus ląstelių yra po vieną branduolį, tačiau yra dvibranduolių (kai kurios kepenų ląstelės) ir daugiabranduolinės (skersaruožių raumenų audinio skaidulose).
Ląstelės branduolys susideda iš membranos, branduolio sulčių, branduolio ir chromatino.
branduolinis apvalkalas atskiria genetinę medžiagą nuo citoplazmos, taip pat reguliuoja dvišalę sąveiką tarp branduolio ir citoplazmos. pastatytas branduolinis apvalkalas dvi uždaros membranos, atskirtos perinuklearine (perinuklearine) erdve, galinčia susisiekti su endoplazminio tinklo kanalėliais. Branduolinė membrana yra persmelkta 80–90 nm skersmens porų, apsuptų siūlinių struktūrų, galinčių susitraukti. Pati pora užpildyta tankia medžiaga. Ši sudėtinga struktūra vadinama porų kompleksas. Porų skaičius priklauso nuo funkcinė būklė ląstelės. Kuo didesnis sintetinis aktyvumas ląstelėje, tuo didesnis jų skaičius. Nustatyta, kad per branduolio membraną aktyviu transportu specialių nešančiųjų medžiagų pagalba praeina ne tik iRNR molekulės, bet ir didelės molekulės, ribosomų dalelės. Ribosomos yra branduolinės membranos paviršiuje, todėl čia vyksta baltymų sintezė. Dalijimosi metu branduolio membrana skyla į mažas pūsleles, iš kurių dukterinėse ląstelėse statomi branduolių apvalkalai.

Ryžiai. 9. Ląstelės branduolio sandaros schema (http://biology-of-cell.narod.ru): 1 - branduolio membrana (dvi membranos - vidinė ir išorinė, bei perinuklearinė erdvė), 2 - branduolio poros, 3 - kondensuotas chromatinas, 4 - difuzinis chromatinas, 5 - branduolys, 6 - tarpchromatino granulės (RNP), 7 - perichromatino granulės (RNP), 8 - perichromatino fibrilės (RNP), 9 - karioplazma, branduolių sultys
pagrindu branduolinės sultys(karioplazmos) sudaro baltymus (įskaitant siūlinius arba fibrilinius, su kuriais susijusi palaikymo funkcija); karioplazmoje yra pirminiai genetinės informacijos transkripcijos produktai – heterobranduolinė RNR (hn-RNR), kurie čia apdorojami, virsdami m-RNR.
branduolys- nemembraninės struktūros, kuriose susidaro ir bręsta ribosominė RNR (rRNR). rRNR genai užima tam tikras vienos ar kelių chromosomų dalis (žmonėms 13-15 ir 21-22 poras) branduolių organizatoriai, kuriame susidaro branduoliai. Tokios sritys metafazėse chromosomose atrodo kaip susiaurėjimai ir vadinamos antriniais susiaurėjimais.
Branduolys apima milžiniškas PKN pirmtakų molekules, iš kurių susidaro mažesnės subrendusios RNR (3–5% sausos branduolio masės) ir baltymų (80–85% sausos masės) (gijinės arba fibrilinės) molekulės. tada susidaro, taip pat lipidai. Brandinimo procese fibrilės virsta ribonukleoproteinų grūdeliais (granulėmis), kurie yra granuliuotas komponentas. Pagrindinė branduolio funkcija yra ribosomų susidarymas. Dalijimosi metu branduolys suyra, o jo pabaigoje formuojasi iš naujo.
Chromatino struktūros nukleoplazmoje išsibarsčiusių gumulėlių pavidalu jie yra tarpfazinė ląstelių chromosomų egzistavimo forma. Chromatino sudėtyje yra DNR kartu su baltymais. Dalijantis DNR grandinės paverčiamos chromosomomis, kurių skaičius ir forma griežtai priklauso nuo rūšies.
organelės (organelės))
Organelės yra nuolatinės citoplazmos struktūros, atliekančios gyvybines funkcijas ląstelėje.
Išskirkite organelius bendrą reikšmę ir ypatingas. Pastarųjų yra daug ląstelėse, kurios yra specializuotos atlikti tam tikrą funkciją, tačiau mažas kiekis taip pat galima rasti ir kitų tipų ląstelėse. Tai apima, pavyzdžiui, žarnyno epitelio ląstelės siurbimo paviršiaus mikrovilkus, trachėjos ir bronchų epitelio blakstienas, sinapsines pūsleles, pernešančias medžiagas - nešiklius. nervinis susijaudinimas iš vienos nervinės ląstelės į kitą arba darbo organo ląstelę – miofibriles, nuo kurių priklauso raumenų susitraukimas. Išsamus specialių organelių svarstymas įtrauktas į histologijos kurso užduotį.
Gyvūnų ir žmogaus ląstelių bendros svarbos organelės apima vamzdinės ir vakuolinės sistemos elementus grubaus ir lygaus endoplazminio tinklo pavidalu, Golgi kompleksą, mitochondrijas, ribosomas ir polisomas, lizosomas, peroksisomas, mikrofibriles ir mikrotubulus, centriolius. ląstelių centras.
Endoplazminis tinklas (ER)
Endoplazminis tinklas yra surištas su membrana sudėtinga sistema kanalėlių, vakuolių ir cisternų, kurios prasiskverbia per visą ląstelės citoplazmą (10 pav.). EPS skirtingose ląstelėse išsivysto nevienodai, o tai siejama su pastarųjų funkcijomis.
Yra dviejų tipų endoplazminis tinklas: šiurkštus(granuliuotas) ir sklandžiai(agranuliuotas). Šiurkštaus tinklo struktūrinė ypatybė yra ta, kad prie jo membranų yra prijungtos ribosomos, dėl kurių jis atlieka tam tikros kategorijos baltymų, kurie daugiausia pašalinami iš ląstelės, pavyzdžiui, išskiriami liaukos ląstelės, sintezės funkciją. Taip pat ant grubios ER susidaro baltymų ir lipidų. citoplazminės membranos ir jų surinkimas. Tankiai supakuotos į sluoksniuotą struktūrą grubaus tinklo cisternos yra aktyviausios baltymų sintezės vietos ir vadinamos ergastoplazma.
Lygiosios ER membranose nėra polisomų. Sklandus EPS yra susijęs su angliavandenių, riebalų ir kitų nebaltyminių medžiagų apykaita, pvz. steroidiniai hormonai(lytinėse liaukose – antinksčių žievės sluoksnyje).

10 pav. Endoplazminis tinklas: 1 - kanalėliai, 2 - polisomos, 3 - pūslelės, 4 - membrana, 5 - rezervuarai; aEPS – agraulinis EPS, PEPS – pereinamasis EPS, GREPS – granuliuotas EPS
Per kanalėlius ir cisternas medžiagos, ypač liaukos ląstelės išskiriama medžiaga, iš sintezės vietos patenka į pakavimo vietą į granules. Kepenų ląstelių srityse, kuriose gausu lygių tinklo struktūrų, kenksmingos medžiagos sunaikinamos ir neutralizuojamos. toksiškos medžiagos kai kurie vaistai (barbitūratai). Lygiojo dryžuotų raumenų tinklo pūslelėse ir kanalėliuose kaupiasi (nusėda) kalcio jonai, kurie atlieka svarbų vaidmenį susitraukimo procese.
Abiejų tipų EPS būdingas pradinis susintetintų produktų kaupimasis ir vėlesnis jų transportavimas į skirtingas ląstelės dalis, ypač į Golgi aparatą.
Ribosomos
Ribosomos yra sferinės ribonukleoproteino dalelės, kurių skersmuo 20–30 nm, neapribotos membrana, kurios apima baltymus ir RNR molekules maždaug lygiomis dalimis. Ribosomos gali laisvai išsidėstyti citoplazmoje arba prisitvirtinti prie išorinio endoplazminio tinklo membranų paviršiaus, ląstelės branduolyje ir mitochondrijose. Kiekviena ribosoma susideda iš didelių ir mažų sudėtingos konfigūracijos subvienetų: mažasis sulenktas telefono imtuvo pavidalu, didysis primena kibirą (11 pav.). Jų susiejimas, atsirandantis esant matricinei (pasiuntinei) RNR (mRNR). Jų sąlyčio vietoje susidaro siauras tarpelis. Viena mRNR molekulė paprastai sujungia keletą ribosomų kaip karoliukų virtinė. Tokia struktūra vadinama polisomas.
Ribosomos susidaro branduoliuose (žr. "Branduolinis")

Ryžiai. 11. Ribosama: 1 - mažas subvienetas, 2 - didelis subvienetas
Baltymų sintezė vyksta ribosomose. Be to, ant hialoplazmos polisomų susidaro baltymai pačios ląstelės poreikiams, o granuliuoto tinklo polisomose sintetinami baltymai, kurie išsiskiria iš ląstelės ir panaudojami organizmo reikmėms (pvz. virškinimo fermentai, voverės Motinos pienas ir taip toliau.).
Golgi aparatas (kompleksas)
„Golgi“ kompleksą sudaro trys pagrindiniai komponentai:
Plokščių rezervuarų sistema, apribota lygiomis membranomis - diktiosomos;
Nukrypsta nuo diktiozomų kanalėlių sistema, kurios sudaro sudėtingą tinklą, supantį ir jungiantį cisternas;
- pūslelės(pūslelės), nuvarstomos nuo galinių kanalėlių dalių.
Golgi komplekso membranos turi tokią pačią struktūrą kaip išorinė ląstelės membrana ir endoplazminio tinklo membranos. Visi šie trys ląstelės komponentai yra tarpusavyje susiję: Golgi aparato pūsleles (pūsleles) ląstelė naudoja plazmalemai kurti. Pats kompleksas susidaro dėl EPS aktyvumo (12 pav.), ką dar liudija ir tai, kad komplekse randama fermentų, tarp jų ir susijusių su polisacharidų ir lipidų sinteze.
Diferencijuotose stuburinių ir žmonių ląstelėse diktiosomos dažniausiai surenkamos perinuklearinėje citoplazmos zonoje.
Golgi komplekse kaupiasi ląstelės atliekos (organoidas netgi sugeba sukaupti toksines medžiagas, kurios patenka į ląstelę iš išorės), kurios turi būti pašalintos iš ląstelės, kurios yra apvilktos membraniniu apvalkalu. Lameliniame komplekse susidaro lizosomos. Ant ribosomų susidarę polipeptidai patenka į endoplazminio tinklo kanalus, o iš ten į Golgi aparatą, kur „bręsta“. Diktiosomose sintetinami polisacharidai, taip pat jų kompleksai su baltymais (glikoproteinais) ir riebalais (glikolipidais). kuriuos vėliau galima rasti glikokalikso ląstelės membranoje.

Ryžiai. 12. Endoplazminio tinklo ir Golgi aparato sąveika ir funkcionavimas (rodyklės rodo kryptingą susintetintų medžiagų transportavimą)
Sintezėje dalyvauja ir Golgi kompleksas konstrukciniai komponentai kolageno tipo ląstelės jungiamasis audinys, kuris atlieka tam tikrą vaidmenį kiaušinio trynio sintezėje, polisacharidų ir lipidų sintezėje.
Lizosomos
Lizosomos yra viena 0,2-0,8 mikrono skersmens membrana apriboti maišeliai, užpildyti sultimis su rūgštiniais hidrolazės fermentais (jų yra daugiau nei 40), kurie veikia kaip fermentai. Virškinimo sistema, katalizuoja nukleorūgščių, baltymų, riebalų, polisacharidų skilimą. Lizosomos aktyviai dalyvauja medžiagų, patenkančių į ląstelę fagocitozės ir pinocitozės būdu, lizės procese. Dėl lizosomų fermentų po mirties gali būti suvirškintos atskiros ląstelių struktūros ir net visos ląstelės. Membrana apsaugo ląstelę, kurioje yra lizosoma, nuo savaiminio virškinimo.
Priklausomai nuo aktyvumo laipsnio, lizosomos klasifikuojamos taip (13 pav.): pirminės lizosomos(skersmuo 100 nm) – neaktyvios organelės, antrinis - organelės, kuriose vyksta virškinimo procesas.Antrinės lizosomos susidaro iš pirminių. Jie skirstomi į heterolizosomos(fagolizosomos) ir autolizosomos(citolizosomos). Pirmajame į ląstelę iš išorės patekusi medžiaga virškinama pinocitozės ir fagocitozės būdu, o antrajame sunaikinamos savo funkciją baigusios ląstelės pačios struktūros. Vadinamos antrinės lizosomos, kuriose virškinimo procesas yra baigtas likutiniai kūnai(telolisosomos). Juose trūksta hidrolazių ir yra nesuvirškintų medžiagų.

13 pav. Lizosomų susidarymas ir funkcijos: 1 - fagosoma; 2 - pinocitinė pūslelė; 3 - pirminė lizosoma; 4 - Golgi aparatai; 5 – antrinė lizosoma
Mitochondrijos
Mitochondrijos(14 pav.) - tai dviejų membranų organoidas, apvalus arba strypo formos, 0,5 µm storio ir iki 5–10 µm ilgio. Daugumoje gyvūnų ląstelių mitochondrijų skaičius svyruoja nuo 150 iki 1500, tačiau moterų lytinėse ląstelėse jų skaičius siekia kelis šimtus tūkstančių. Spermatozoiduose dažnai yra viena milžiniška mitochondrija, spirale susisukusi aplink ašinę žvynelio dalį.
Išorinė membrana atskiria mitochondrijas nuo hialoplazmos. Vidinė membrana sudaro lapo formos invaginacijas ( cristae) arba vamzdinis ( kanalėlių) formuojasi.Vidinė membrana padalija mitochondrijų ertmę į dvi kameras, sukurdama tarpmembraninę erdvę tarp membranų ir vidinę mitochondrijų erdvę, užpildytą matrica, kuriame randami 20-40 nm skersmens grūdeliai. Jie kaupia kalcio ir magnio jonus, taip pat polisacharidus, tokius kaip glikogenas. Matricoje yra savo organelių baltymų biosintezės aparatas. Ją vaizduoja 2–6 žiedinės ir be histonų (kaip ir prokariotuose) DNR molekulės kopijos, ribosomos, transportinės RNR (tRNR) rinkinys, DNR replikacijos fermentai, paveldimos informacijos transkripcija ir vertimas. Savęs-DNR genai koduoja mitochondrijų rRNR ir tRNR nukleotidų sekas, taip pat kai kurių organelių baltymų, daugiausia jo vidinės membranos, aminorūgščių sekas.DNR yra užkoduotos daugumos mitochondrijų baltymų aminorūgščių sekos (pirminė struktūra). ląstelės branduolio ir susidaro už organelių ribų citoplazmoje.

14 pav. Mitochondrijų struktūra
Mitochondrijose yra oksidacinių fermentų sistema, dalyvaujanti ląstelių kvėpavimo procesuose. Įjungta išorinė membrana o aplinkinėje hialoplazmoje vyksta aerobinės oksidacijos procesai ( glikolizė), ir ant vidinės membranos (pusėje, nukreiptoje į matricą) - oksidacinis fosforilinimas- tai apdoroja organinės medžiagos suskaidomas į vandenį ir anglies dioksidą dalyvaujant deguoniui. Išsiskyrusi energija kaupiama ATP pavidalu. Ši energija tik iš dalies išleidžiama „vidiniams poreikiams“ ir dauguma išleista procesams, vykstantiems už mitochondrijų ribų.
Iš šalutinių mitochondrijų funkcijų galima paminėti dalyvavimą steroidinių hormonų ir kai kurių aminorūgščių (pavyzdžiui, glutamino) sintezėje.
Ląstelių centras (centrosoma)
Ląstelių centras(15 pav.) – bemembranos organelės, pateiktos ląstelėje centrioliai(nuo 2 iki 10), apsuptas diferencijuoto citoplazmos ploto, formuojantis centrosfera. Paprastai ląstelių centras esantis geometriniame ląstelės centre, taigi ir jos pavadinimas.
Centriolė turi „tuščiavidurio“ cilindro formą, kurio skersmuo yra apie 150 nm, o ilgis – 300–500 nm. Jos sienelę sudaro 27 mikrovamzdeliai, sugrupuoti į 9 tripletus, iš kurių išsikiša statmeni kūnai - palydovai kurios sudaro centrosferą. Pirmojo tripleto mikrotubulo (A-mikrotubulo) skersmuo yra apie 25 nm, o sienelės storis - 5 nm, kurį sudaro 13 rutulinių subvienetų. Kiekvieno tripleto ilgis lygus centriolės ilgiui. Antrasis ir trečiasis (B ir C) mikrovamzdeliai yra neišsamūs, juose yra 11 subvienetų ir yra glaudžiai greta jų kaimynų. Kiekvienas tripletas yra iki tokio cilindro spindulio maždaug 400 kampu. Be mikrovamzdelių, centriolė apima daugybę papildomų struktūrų. Vadinamosios „rankenos“, ataugos, nukrypsta nuo A mikrovamzdelio, iš kurių vienas (išorinis) nukreiptas į kaimyninio tripleto C mikrotubulą, o kitas (vidinis) - į cilindro centrą. Diplosomoje centrioliai išsidėstę stačiu kampu vienas kito atžvilgiu. Iš dviejų centriolių vienas vadinamas "motininiu", kitas - "dukra", pastarosios išilginė ašis yra statmena išilginė ašis motininės centriolės.Toks organoidas pateikiamas ramybės ląstelėse, besidalijančiose ląstelėse ląstelės centras tampa kompleksinės dalijimosi aparato struktūros dalimi. Nesiskiriančiose ląstelėse centrosomos nustato epitelio ląstelių poliškumą ir yra šalia Golgi aparato. Toks centrosomų ryšys su Golgi aparatu būdingas daugeliui ląstelių, įskaitant kraujo ląsteles ir nervų ląstelės. Centrosomos dažnai yra šalia branduolio, esančios jo invaginacijos zonose. Pavyzdžiui, polimorfiniuose leukocituose (neurofiluose) centrosoma yra pasagos formos branduolio invaginacijos viduje. Centrosfera išnyksta tam tikrose dalijimosi fazėse (daugelyje ląstelių ji išvis neegzistuoja).

Ryžiai. 15. Ląstelės centras: I - ląstelės centras, II - išcentrinė struktūra: 1 - centriolė, 2 - mirotubuliai, 3 - tripletas, 4 - palydovai.
Centriolių funkcija – mitozinių verpstės gijų formavimas, kurias formuoja ir mikrovamzdeliai.Centrioliai poliarizuoja ląstelių dalijimosi procesą, užtikrindami seserinių chromatidžių (chromosomų) atsiskyrimą mitozės anafazėje.
Ląstelių centras dalyvauja dalijimosi verpstės konstrukcijoje, citoplazminių mikrotubulių, blakstienų ir žvynelių susidaryme.
Mikrovamzdeliai -įvairaus ilgio vamzdiniai dariniai, kurių išorinis skersmuo 24 nm, spindžio plotis 15 nm, sienelės storis apie 5 nm Mikrovamzdeliai yra pagrindinėje citoplazmos medžiagoje, tačiau jų išsidėstymo skirtingose ląstelėse pobūdis yra ne tas pats. Mikrotubulai taip pat vaidina svarbų vaidmenį konstrukciniai elementaižvyneliai, blakstienos, mitozinis verpstė, centrioliai.
Mikrotubulai yra sukurti iš rutulinio baltymo tubulino. Mikrovamzdeliai daro daug įvairios funkcijos. Laisvoje būsenoje mikrotubulai atlieka atraminę funkciją, nustatydami ląstelių formą, taip pat yra kryptingo tarpląstelinių komponentų judėjimo veiksniai; jie praeina per aksonus. nervinių skaidulų, dalyvauja formuojant dalijimosi veleno gijas.
Mikrofilamentai vadinami ilgais plonais dariniais, kartais formuojančiais ryšulius ir aptinkamais visoje citoplazmoje. Yra keli skirtingi tipai mikrofilamentai. aktino mikrofilamentai dėl juose esančių susitraukiančių baltymų (aktino) jie laikomi struktūromis, kurios teikia ląstelių formos judesiai, tokie kaip ameboidas. Jiems taip pat priskiriamas karkasinis vaidmuo ir dalyvavimas organelių ir hialoplazmos sekcijų viduląstelinių judėjimų organizavime. Ląstelių periferijoje po plazmalema, taip pat perinuklearinėje zonoje randami 10 nm storio mikrofilamentų ryšuliai - tarpiniai siūlai. Epitelio, nervų, glijos, raumenų ląstelėse, fibroblastuose jie yra sudaryti iš skirtingų baltymų. Tarpinės gijos, matyt, atlieka mechaninę rėmo funkciją.Aktino mikrofibrilės ir tarpinės gijos, kaip ir mikrovamzdeliai, susidaro iš subvienetų. Dėl šios priežasties jų kiekis priklauso nuo polimerizacijos ir depolimerizacijos procesų santykio.
mikrokūnas sudaro organelių grupę. Tai burbuliukai, apriboti viena 0,1-1,5 mikrono skersmens membrana su smulkiagrūde matrica ir dažnai kristaloidiniais arba amorfiniais baltymų inkliuzais. Ši grupė visų pirma apima peroksisomos. Juose yra oksidazės fermentų, kurie katalizuoja vandenilio peroksido susidarymą, kuris, būdamas toksiškas, vėliau sunaikinamas veikiant peroksidazės fermentui. Šios reakcijos yra įtrauktos į įvairius medžiagų apykaitos ciklus, pavyzdžiui, mainus šlapimo rūgštis kepenų ir inkstų ląstelėse. Kepenų ląstelėje peroksisomų skaičius siekia 70-100.
Pagrindiniai žodžiai ir sąvokos: Skirstymas, lipidų dvisluoksnis, plazmolema, pasyvus transportas, aktyvus transportas, natrio-kalio pompa, egzocitozė, endocitozė, fagocitozė, pinocitozė, protoplazma, ląstelės branduolys, citoplazma, hialoplazma, porų kompleksas, branduolinė membrana, karioplazma, branduolys, chromatino struktūros, DNR, organelės, endoplazminis tinklas (ER), lygus ER (agraulinis), šiurkštus ER (gumbuotas), ergastoplazma, polisoma, ribosoma, Golgi aparatas (kompleksas), diktiosoma, lizosoma, pirminės lizosomos, antrinės lizosomos, heterolizosomos (fagolizosomos, autolizidosomos), autolizidosomos telolisosomos), mitochondrijos, krištolos, mitochondrijų matrica, kanalėliai, peroksisomos, glikolizė, ląstelių centras (centrosoma), centriolė, mikrovamzdeliai, mikrofilamentai, oksidacinis fosforilinimas
Struktūra, ultrastruktūra ir funkcija ląstelių organelėsŠiuo metu tiriama naudojant šiuos pagrindinius metodus: šviesus ir elektroninis, tamsus laukas, fazinis kontrastas, poliarizacija, liuminescencinis mikroskopija naudojama fiksuotų ląstelių struktūrai, ultrastruktūrai tirti ir diferencinei centrifugacijai, kuri leidžia išskirti atskirus organelius ir juos analizuoti citocheminiais, biocheminiais, biofiziniais ir kitais metodais.
Šviesos mikroskopija.
Metodo principas yra tas, kad šviesos spindulys, praeinantis per objektą, patenka į objektyvo lęšių sistemą ir sukuria pirminį vaizdą, kuris padidinamas okuliaro lęšių pagalba. Pagrindinė optinė mikroskopo dalis, kuri lemia pagrindines jo galimybes, yra objektyvas.
IN šiuolaikiniai mikroskopai lęšiai yra keičiami, todėl galite tirti ląsteles skirtingais didinimais. Pagrindinė savybė mikroskopas kaip optinė sistema yra skiriamoji geba, t.y. galimybė pateikti atskirą dviejų arti vienas kito objektų vaizdą.
Objektyvo pateikiamus vaizdus galima padidinti daug kartų naudojant stiprų okuliarą arba, pavyzdžiui, projekcijas į ekraną (iki 10 5 kartų). Šviesos mikroskopo skiriamąją gebą riboja šviesos bangos ilgis: kuo trumpesnis bangos ilgis, tuo didesnė skiriamoji geba. Paprastai šviesos mikroskopuose naudojami šviesos šaltiniai matomoje spektro srityje (400-700 nm), todėl maksimali mikroskopo skiriamoji geba šiuo atveju negali būti didesnė nei 200-350 nm (0,2-0,35 mikronai). Jei naudojate violetinę šviesą (260–280 nm), galite padidinti skiriamąją gebą iki 130–140 nm (0,13–0,14 mikronų). Tai bus šviesos mikroskopo teorinės skiriamosios gebos riba, kurią lemia šviesos banginis pobūdis.
Taigi viskas, ką šviesos mikroskopas gali duoti kaip pagalbinį prietaisą mūsų akiai, yra padidinti jos skiriamąją gebą maždaug 1000 kartų (žmogaus plika akies skiriamoji geba yra apie 0,1 mm, tai yra 100 mikronų). Tai yra „naudingas“ mikroskopo padidinimas, virš kurio tik padidinsime vaizdo kontūrus, neatskleisdami jame naujų detalių. Todėl, naudojant matomą šviesos sritį, 0,2–0,3 µm yra galutinė šviesos mikroskopo skiriamosios gebos riba.
Elektroninė mikroskopija.
Dėl nuskaitymo elektroninis mikroskopas medžiaga dažnai užšaldoma, kad susidarytų ledu padengtas paviršius. Tokiu atveju vandenyje tirpių medžiagų vandens praradimas neįtraukiamas, o cheminiai struktūrų pokyčiai taip pat mažesni. Analizuojant duomenis, gautus naudojant elektroninį mikroskopą, reikia atsiminti, kad šiuo metodu tiriamos statinės ląstelės būsenos tuo metu, kai greitai sustoja citoplazmos judėjimas, kurį sukelia fiksuojančių cheminių medžiagų veikimas.
Tamsiojo lauko mikroskopija.
Jo esmė ta, kad kaip dulkių dalelės šviesos spindulyje ( Tyndall efektas) šoninio apšvietimo ląstelėje šviečia smulkiausios dalelės (mažiau nei 0,2 mikrono), kurių atspindėta šviesa patenka į mikroskopo lęšį. Šis metodas buvo sėkmingai naudojamas tiriant gyvas ląsteles.
Biopolimerų sintezės vietų lokalizacijai nustatyti, medžiagų pernešimui ląstelėje nustatyti, atskirų ląstelių migracijai ar savybėms stebėti, jie plačiai naudojami. metodas autoradiografija - izotopais paženklintų medžiagų registracija. Pavyzdžiui, naudojant šį metodą, naudojant žymėtus RNR pirmtakus, buvo parodyta, kad visa RNR sintetinama tik tarpfaziniame branduolyje, o citoplazminės RNR buvimas yra susintetintų molekulių migracijos iš branduolio rezultatas.
Citologijoje įvairios analitiniai ir paruošiamieji metodai biochemija. Pastaruoju atveju atskirų frakcijų pavidalu galima gauti įvairių ląstelių komponentai ir ištirti jų chemiją, ultrastruktūrą ir savybes. Šiuo metu beveik visos ląstelių organelės ir struktūros gaunamos grynų frakcijų pavidalu.
Vienas iš pagrindinių izoliacijos būdų ląstelių struktūros yra diferencinė (atskyrimo) centrifuga. Jo taikymo principas – laikas, per kurį dalelės nusės homogenate, priklauso nuo jų dydžio ir tankio: kuo dalelė didesnė ar sunkesnė, tuo greičiau nusės į mėgintuvėlio dugną. Siekiant pagreitinti šį nusėdimo procesą, naudojami centrifugos sukuriami pagreičiai.
Pakartotinai frakciškai centrifuguojant mišrias subfrakcijas, galima gauti grynas frakcijas. Smulkesnio frakcijų atskyrimo atvejais naudojamas centrifugavimas sacharozės tankio gradiente, kuris leidžia gerai atskirti komponentus, net šiek tiek besiskiriančius vienas nuo kito. specifinė gravitacija. Prieš analizuojant biocheminiais metodais gautų frakcijų grynumas turi būti patikrintas elektroniniu mikroskopu.
Kontroliniai klausimai:
1. Gyvosios materijos organizavimo lygiai
2. Ląstelinė organizmų organizavimo teorija
3. Citologijos tyrimo metodai
4. Citologijos uždaviniai ir dalykas
5. Šviesos mikroskopo įtaisas
6. Elektroninio mikroskopo prietaisas
7. Citologinio darbo saugos priemonės
8. Biologinės medžiagos paruošimo citologiniam tyrimui reikalavimai
9. Fiksuojančios medžiagos, veikimo mechanizmas
10. Citochemija, medžiagų reikalavimai ir galimybės
11. Kiekybinė analizė (morfometrija), reikalavimai ir galimybės
12. Artefaktai citologijoje, rezultatų objektyvavimo būdai
1. Zavarzinas A.A., Kharazova A.D. Bendrosios citologijos pagrindai. - L., 1982 m.
2. Chentsov Yu.S. Citologijos pagrindai. - M., 1984 m.
3. Šubnikova E.A. Funkcinė audinių morfologija. - M., Maskvos valstybinio universiteto leidykla, 1981 m.